家蠶卵形指數(shù)對(duì)孵化率的影響及孵化相關(guān)基因表達(dá)分析
2020-03-24 10:02:50王雪珍徐嬌王閃閃浦月霞沈興家唐順明
南方農(nóng)業(yè)學(xué)報(bào) 2020年12期
王雪珍 徐嬌 王閃閃 浦月霞 沈興家 唐順明


摘要:【目的】分析家蠶卵形指數(shù)對(duì)孵化率的影響規(guī)律,以及孵化酶基因(BmHE I和BmHE II )、幾丁質(zhì)酶基因(Chitinase)和周期蛋白基因(Period)在不同卵形間的表達(dá)規(guī)律,為闡明家蠶卵形指數(shù)與孵化率的相關(guān)性及其分子機(jī)理打下基礎(chǔ)。【方法】調(diào)查家蠶Nistari品種野生型正常形品種(Nistari+)及其新突變體紡錘形品種(Nistari-)的卵形指數(shù)、孵化率、受精率和轉(zhuǎn)青死卵率,分析其卵形指數(shù)與孵化率的相關(guān)性;并通過實(shí)時(shí)熒光定量PCR檢測(cè)蠶卵胚胎發(fā)育后期孵化相關(guān)基因的表達(dá)情況。【結(jié)果】Nistari+蠶卵的平均卵形指數(shù)為1.26,極顯著低于Nistari-蠶卵(1.73)(P<0.01,下同);孵化率為96.22%,極顯著高于Nistari-蠶卵(51.33%)。當(dāng)Nistari+蠶卵的卵形指數(shù)為1.24~1.25時(shí),其孵化率(97.39%)和受精率(98.84%)均最高,轉(zhuǎn)青死卵率最低(2.61%);而Nistari-蠶卵的卵形指數(shù)為1.77~1.81時(shí),其孵化率(56.52%)和受精率(74.87%)均最高,轉(zhuǎn)青死卵率最低(43.48%)。相關(guān)分析結(jié)果表明,Nistari蠶卵孵化率、受精率和轉(zhuǎn)青死卵率與卵形指數(shù)存在一定的相關(guān)性,且受卵形指數(shù)影響。在蠶卵胚胎發(fā)育后期(孵化前),Nistari+蠶卵的BmHE I、BmHE II、Chitinase和Period基因大量表達(dá),對(duì)應(yīng)的相對(duì)表達(dá)量顯著(P<0.05)或極顯著高于NISTAR-蠶卵。【結(jié)論】家蠶Nistari品種的卵形指數(shù)與孵化率存在一定相關(guān)性,而BmHE I、BmHE II、Chitinase和Period基因的表達(dá)差異是導(dǎo)致Nistari-蠶卵與Nistari+蠶卵孵化率差異的分子基礎(chǔ)。
關(guān)鍵詞: 家蠶;Nistari品種;卵形指數(shù);孵化率;孵化相關(guān)基因
中圖分類號(hào): S882.4? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? 文獻(xiàn)標(biāo)志碼: A 文章編號(hào):2095-1191(2020)12-3109-07
Abstract:【Objective】The effects of silkworm egg shape index (ESI) on hatching rate(HR), and the expression patterns of hatching enzyme genes(BmHE I and BmHE II), chitinase gene(Chitinase) and period protein gene(Period ) in different egg shapes were investigated, which laid a foundation for elucidating the correlation between egg shape index and hatching rate of silkworm eggs and its molecular mechanism. 【Method】The ESI, HR, fertilization rate(FR), and mortality rate in egg with pigmentary body of embryo(MR) were investigated between the wild type normal egg variety(Nistari+)and its new mutant spindle egg variety (Nistari-), and the correlation of ESI with HR were analyzed. And investigated the differences in gene expression levels related to hatching process in the later stages of embryonic development by real-time quantitative PCR. 【Result】The mean ESI of Nistari+ was 1.26, which was extremely lower than that of Nistari-(1.73)(P<0.01,the same below); the HR of Nistari+was 96.22%, which was extremely higher than that of Nistari- silkworm eggs(51.33%). When the ESI of? Nistari+ was 1.24-1.25, the HR (97.39%) and FR(98.84%) were the highest and the MR(2.61%) was the lowest, while when the ESI of Nistari- was 1.77-1.81, the HR(56.52%) and FR(74.87%) of its eggs were the highest, MR was the lowest(43.48%). The results of correlation analysis showed that there was a certain correlation between the Nistari ESI and HR, FR and MR, and was affected by ESI. In the later stages of silkworm egg embryo development(before hatching), the expression levels of BmHE I, BmHE II, Chitinase and Period genes of Nistari+ silkworm eggs were high, and were significantly(P<0.05) or extremely(P<0.01) higher than those of Nistari_ silkworm eggs(P<0.05). 【Conclusion】There is certain correlation between ESI and HR of silkworm Nistari varieties. The differe-nces in expression of four genes, BmHE I, BmHE II, Chitinase and Period, are the molecular basis for the differences in HR between silkworm varieties of Nistari- and Nistari+.
Key words: sailkworm(Bombyx mori); Nistari varieties; egg shape index; hatching rate; hatching related genes
Foundation item:National Natural Science Foundation of China(31372376,31672490);Jiangsu Natural Science Foundation(BK20141285)
0 引言
【研究意義】家蠶(Bombyx mori)是一種重要的鱗翅目經(jīng)濟(jì)昆蟲,在農(nóng)業(yè)經(jīng)濟(jì)體系中占據(jù)重要地位。孵化率是衡量蠶種質(zhì)量的一項(xiàng)關(guān)鍵性指標(biāo),其影響因素包括蠶種品質(zhì)、制種季節(jié)、蠶卵營(yíng)養(yǎng)、冷藏時(shí)間和催青條件等(李燕飛等,2018;高海青等,2019),其中蠶種的卵形及卵重等品質(zhì)指標(biāo)對(duì)蠶卵孵化率有明顯影響(潘明德等,2019)。因此,研究分析家蠶卵形指數(shù)對(duì)孵化率的影響及其分子機(jī)理,對(duì)提高蠶種孵化率、蠶種選育與保存具有重要意義。【前人研究進(jìn)展】昆蟲胚胎孵化后才能進(jìn)行后續(xù)的生長(zhǎng)發(fā)育(Young et al.,2000;Bowles et al.,2008),除了溫度、濕度和光照等外部條件對(duì)昆蟲的胚胎發(fā)育有影響外,其內(nèi)部的基因表達(dá)調(diào)控網(wǎng)絡(luò)也會(huì)對(duì)昆蟲孵化及后期發(fā)育有顯著影響(張艷,2020)。高頌等(2012)研究發(fā)現(xiàn),幾丁質(zhì)酶(Chitinase)主要在昆蟲表皮和中腸中表達(dá),與幼蟲蛻皮、化蛹和成蟲羽化及翅的延展有關(guān)。Tao等(2017)研究表明,大多數(shù)昆蟲的孵化過程受生物鐘調(diào)控機(jī)制嚴(yán)格調(diào)節(jié),其中周期蛋白基因(Period)在生物鐘信號(hào)通路中扮演重要角色。家蠶孵化也呈明顯的時(shí)間節(jié)律性,故推測(cè)其受Period基因的調(diào)控。至今,有關(guān)蠶卵孵化的研究已有較多報(bào)道。Lu等(2010)在蠶卵孵化過程中發(fā)現(xiàn),消化液從成熟胚中分泌溶解并軟化卵殼,從而有利于蟻蠶孵化。Tang等(2012)研究表明,在蠶卵孵化過程中BmHE I和BmHE II兩種孵化酶基因的表達(dá)趨勢(shì)與其孵化進(jìn)程基本一致。Tao等(2017)研究發(fā)現(xiàn),家蠶蟻蠶從含有大量幾丁質(zhì)的卵殼中孵化出來,需依賴幾丁質(zhì)酶將幾丁質(zhì)降解為寡糖,即蠶卵的孵化過程需要幾丁質(zhì)酶參與。【本研究切入點(diǎn)】已有研究證實(shí)禽蛋孵化率與蛋形指數(shù)(禽蛋長(zhǎng)軸與短軸間的比值)有關(guān)。朱振鵬等(2013)研究認(rèn)為,揚(yáng)州鵝種蛋的蛋形指數(shù)在1.32~1.46時(shí)其孵化率最高;張楠楠等(2014)研究發(fā)現(xiàn),肉雜雞種蛋的蛋形指數(shù)在1.25~1.30和1.31~1.37 時(shí)孵化效果較好,而在1.19~1.24時(shí)孵化效果較差。但針對(duì)家蠶卵形指數(shù)與其卵孵化率間的相關(guān)研究至今未見報(bào)道。【擬解決的關(guān)鍵問題】調(diào)查家蠶Nistari品種野生型正常形卵(Nistari+)及其新突變體紡錘形卵(Nistari-)的卵形指數(shù)、孵化率、受精率和轉(zhuǎn)青死卵率,分析家蠶卵形指數(shù)對(duì)孵化率的影響規(guī)律,以及孵化酶基因(BmHE I和BmHE II)、幾丁質(zhì)酶基因(Chitinase)和周期蛋白基因(Period)在不同卵形間的表達(dá)規(guī)律,為闡明家蠶卵形指數(shù)與孵化率的相關(guān)性及其分子機(jī)理打下基礎(chǔ)。
1 材料與方法
1. 1 試驗(yàn)材料
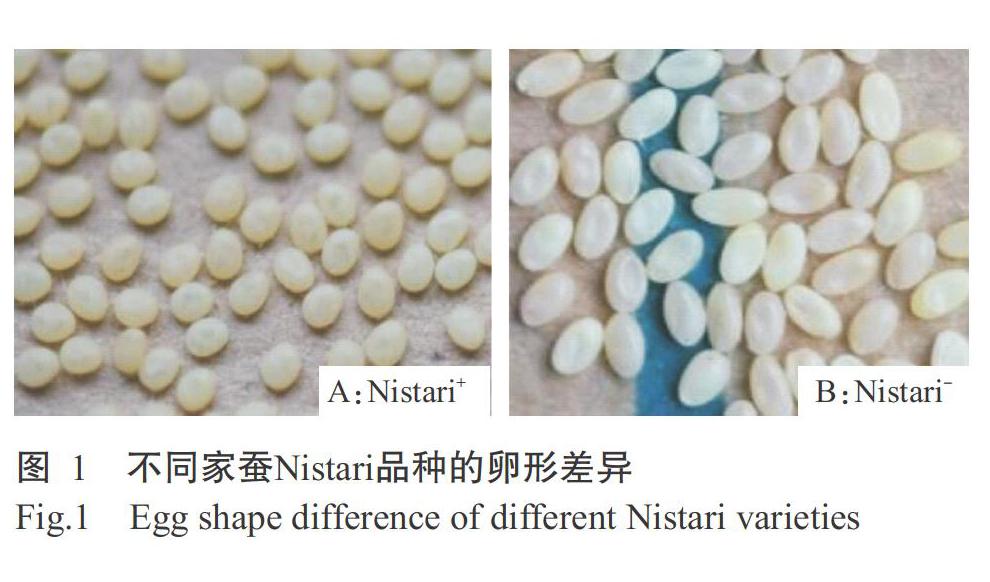
家蠶Nistari品種的野生型正常形品種Nistari+(圖1-A)及其新突變體紡錘形品種Nistari-(圖1-B)由江蘇科技大學(xué)/中國農(nóng)業(yè)科學(xué)院蠶業(yè)研究所保存提供。其中,Nistari+品種的蠶卵呈橢圓形,Nistari-品種的蠶卵呈紡錘形。2個(gè)品種的蠶卵孵化后,幼蟲在25 ℃、相對(duì)濕度(65±5)%、12 h光照/12 h黑暗的條件下桑葉飼養(yǎng),上蔟結(jié)繭、化蛹、羽化產(chǎn)卵和蠶卵催青等按常規(guī)管理?xiàng)l件進(jìn)行操作。
1. 2 品種間卵形指數(shù)及孵化率調(diào)查
以正常產(chǎn)卵的框制種Nistari+和Nistari-蠶卵為材料,以蛾圈為單位,每個(gè)蛾圈調(diào)查50粒卵,調(diào)查10個(gè)蛾圈。在Leica體視顯微鏡下測(cè)量蠶卵的縱徑和橫徑,計(jì)算卵形指數(shù)。調(diào)查后的蠶種按常規(guī)催青條件進(jìn)行催青,待蠶卵完全孵化后,統(tǒng)計(jì)不同品種蛾圈內(nèi)的總卵數(shù)、孵化卵數(shù)、受精卵數(shù)和轉(zhuǎn)青死卵數(shù),計(jì)算蠶卵的孵化率、受精率和轉(zhuǎn)青死卵率。3次重復(fù),取平均值,比較Nistari+和Nistari-品種間的卵形指數(shù)和孵化率差異。
卵形指數(shù)=縱徑長(zhǎng)度(長(zhǎng)軸)/橫徑長(zhǎng)度(短軸)
孵化率(%)=孵化蠶卵數(shù)/受精卵數(shù)×100
受精率(%)=受精卵數(shù)/總卵數(shù)×100
轉(zhuǎn)青死卵率(%)=(受精卵數(shù)-孵化卵數(shù))/受精卵數(shù)×100
1. 3 品種內(nèi)卵形指數(shù)及孵化率調(diào)查
以正常產(chǎn)卵Nistari+和Nistari-散卵為材料,每個(gè)供試品種取3000粒卵,按1.2的方法計(jì)算卵形指數(shù)后放入預(yù)先標(biāo)記好的不同卵形指數(shù)小組內(nèi),每組100粒卵。調(diào)查后的蠶種按常規(guī)催青條件進(jìn)行催青,待蠶卵完全孵化后,按1.2的方法調(diào)查不同卵形指數(shù)小組內(nèi)的總卵數(shù),統(tǒng)計(jì)孵化卵數(shù)、受精卵數(shù)和轉(zhuǎn)青死卵數(shù),計(jì)算蠶卵的孵化率、受精率和轉(zhuǎn)青死卵率。3次重復(fù),取平均值,分別分析Nistari+蠶卵及Nistari-蠶卵不同卵形指數(shù)與孵化率、受精率和轉(zhuǎn)青死卵率的關(guān)系,并進(jìn)行相關(guān)分析。
1. 4 4個(gè)孵化相關(guān)基因表達(dá)差異比較
以肌動(dòng)蛋白3(Actin3)為內(nèi)參基因,比較蠶卵胚胎后期孵化相關(guān)基因(BmHE I、BmHE II、Chitinase和Period)的表達(dá)差異。所用引物及其序列信息見表1,均委托生工生物工程(上海)股份有限公司合成。以Nistari+和Nistari-蠶卵催青第7、8、9和10 d(蠶卵胚胎發(fā)育后期)的散卵各100 mg為材料,按TRIzol試劑盒說明提取總RNA,獲得的蠶卵總RNA用適量DEPC水溶解,并稀釋至500 ng/?L。取1 ?g RNA反轉(zhuǎn)錄合成cDNA第一鏈,具體步驟按照First Strand cDNA Synthesis Kit試劑盒(TaKaRa公司)說明進(jìn)行操作。以cDNA為模板進(jìn)行實(shí)時(shí)熒光定量PCR檢測(cè),反應(yīng)體系50.0 ?L:2×Ultra SYBR Mixture 25.0 ?L,上、下游引物(10 ?mol/L)各1.0 ?L,cDNA模板2.0 ?L,ddH2O補(bǔ)足至50.0 ?L。擴(kuò)增程序:95 ℃預(yù)變性2 min;95 ℃ 15 s,59 ℃ 30 s,72 ℃ 30 s,進(jìn)行30個(gè)循環(huán)。
1. 5 統(tǒng)計(jì)分析
試驗(yàn)數(shù)據(jù)采用Excel 2016進(jìn)行統(tǒng)計(jì)整理,運(yùn)用SPSS 17.0進(jìn)行差異顯著性分析,并以O(shè)rigin 8.0制圖。
2 結(jié)果與分析
2. 1 Nistari+和Nistari-蠶卵的卵形指數(shù)及其孵化率
由表2可知,Nistari+蠶卵的平均卵形指數(shù)(1.26)極顯著低于Nistari-蠶卵(1.73)(P<0.01,下同);Nistari+蠶卵的卵形指數(shù)最大值為1.29、最小值為1.22,而Nistari-蠶卵的卵形指數(shù)最大值為1.81、最小值為1.65。Nistari+和Nistari-蠶卵的孵化率分別為96.22%和51.33%,二者差異極顯著。可見,平均卵形指數(shù)較小有利于Nistari+蠶卵孵化,平均卵形指數(shù)較大不利于Nistari-蠶卵孵化。
2. 2 Nistari+蠶卵卵形指數(shù)與孵化率、受精率和轉(zhuǎn)青死卵率的關(guān)系
由表3可知,第1~4組Nistari+蠶卵的卵形指數(shù)分別在1.22~1.23、1.24~1.25、1.26~1.27和1.28~1.29,其中以第4組最高、第1組最低;第2組Nistari+蠶卵的孵化率最高,為97.39%,極顯著高于其他3個(gè)組別;第1、第3、第4組Nistari+蠶卵的孵化率分別為95.88%、96.93%和94.68%,組間差異均達(dá)極顯著水平。第2組Nistari+蠶卵的受精率最高,為98.84%,極顯著高于其他3個(gè)組別;第2組Nistari+蠶卵的轉(zhuǎn)青死卵率最低,僅為2.61%,極顯著低于其他3個(gè)組別。可見,卵形指數(shù)為1.24~1.25更有利Nistari+蠶卵受精和孵化,其受精率和孵化率均最高。
2. 3 Nistari-蠶卵卵形指數(shù)與孵化率、受精率和轉(zhuǎn)青死卵率的關(guān)系
由表4可知,第1~4組Nistari-蠶卵的卵形指數(shù)分別為1.65~1.68、1.69~1.72、1.73~1.76和1.77~1.81,其中以第4組最高、第1組最低;第4組Nistari-蠶卵的孵化率最高,為56.52%,極顯著高于其他3個(gè)組別;第1、第2和第3組Nistari-蠶卵的孵化率分別為50.43%、47.87%和50.48%,其中第1組與第3組的差異不顯著(P>0.05,下同),但二者極顯著高于第2組。第4組Nistari-蠶卵的受精率最高(74.87%),極顯著高于其他3個(gè)組別;第2組Nistari-蠶卵的受精率最低(54.59%),極顯著低于其他3個(gè)組別。第4組Nistari-蠶卵的轉(zhuǎn)青死卵率最低,為43.48%,極顯著低于其他3個(gè)組別。可見,卵形指數(shù)在1.77~1.81更有利于Nistari-蠶卵受精和孵化,受精率和孵化率均最高。
2. 4 卵形指數(shù)與蠶卵孵化率、受精率和轉(zhuǎn)青死卵率的相關(guān)分析結(jié)果
根據(jù)統(tǒng)計(jì)學(xué)正態(tài)分布規(guī)律及參考禽蛋相關(guān)蛋形指數(shù)與孵化率的相關(guān)分析方法(韋偏等,2019;黃得純等,2020),采用SPSS 17.0中的最小二乘法進(jìn)行擬合回歸方程相關(guān)分析。由表5可知,卵形指數(shù)與Nistari+蠶卵孵化率、受精率和轉(zhuǎn)青死卵率所對(duì)應(yīng)的回歸方程分別為:y=-2350x2+5878.2x-3578.4、y= -2500x2+6267.6x-3829.9和y=2350x2-5878.2x+3578.4。卵形指數(shù)與Nistari-蠶卵孵化率、受精率和轉(zhuǎn)青死卵率所對(duì)應(yīng)的回歸方程分別為:y=1185.6x2-4045.5x+3499.3、y=2780.9x2-9492.6x+8156.6和y=-1185.6x2+4045.5x-3499.3。綜上所述,蠶卵孵化率、受精率和轉(zhuǎn)青死卵率與卵形指數(shù)存在一定的相關(guān)性,且受卵形指數(shù)影響。
2. 5 Nistari+和Nistari-蠶卵胚胎發(fā)育后期孵化相關(guān)基因的表達(dá)差異
在常規(guī)條件下進(jìn)行催青,蠶卵在第11 d孵化。選取蠶卵胚胎發(fā)育后期即催青第7、8、9和10 d的Nistari+和Nistari-蠶卵,調(diào)查比較BmHE I、BmHE II、Chitinase和Period等孵化相關(guān)基因的表達(dá)情況。結(jié)果(圖2)顯示,Nistari+和Nistari-蠶卵的BmHE I和BmHE II基因相對(duì)表達(dá)量在胚胎發(fā)育后期至孵化前均呈逐漸上升趨勢(shì),說明BmHE I和BmHE II基因參與蠶卵孵化過程;Nistari+蠶卵的BmHE I基因相對(duì)表達(dá)量在催青第7~10 d均極顯著高于Nistari-蠶卵(圖2-A),Nistari+蠶卵的BmHE II基因相對(duì)表達(dá)量在催青第7~8 d表現(xiàn)為顯著高于Nistari-蠶卵(P<0.05,下同),在催青第9~10 d則表現(xiàn)為極顯著性高于Nistari-蠶卵(圖2-B)。可見,家蠶的BmHE I和BmHE II基因均在蠶卵胚胎發(fā)育后期(孵化前)大量合成,且Nistari+蠶卵的合成量遠(yuǎn)高于Nistari-蠶卵,可能是導(dǎo)致Nistari-與Nistari+品種間孵化率極顯著差異的主要原因。
由圖3-A可看出,在催青第7、8、9和10 d,Nistari+蠶卵的Chitinase基因相對(duì)表達(dá)量(分別為0.002、0.004、0.008和0.035)均極顯著高于Nistari-蠶卵(分別為0.001、0.002、0.004和0.010),Nistari+蠶卵的Chitinase基因相對(duì)表達(dá)量在孵化前1 d(催青第10 d)達(dá)最高值(0.035),遠(yuǎn)高于催青第7~9 d的Nistari+蠶卵Chitinase基因相對(duì)表達(dá)量。說明Nistari+蠶卵的Chitinase基因在孵化前1 d大量合成,且極顯著高于Nistari-蠶卵,可能是導(dǎo)致Nistari-蠶卵與Nistari+蠶卵孵化率差異的原因之一。
由圖3-B可看出,在催青第7、8、9和10 d,Nistari+蠶卵的Period基因相對(duì)表達(dá)量(分別為0.0082、0.0500、0.0253和0.0125)均極顯著高于Nistari-蠶卵(分別為0.0060、0.0039、0.0227和0.0080),Nistari+蠶卵的Period基因相對(duì)表達(dá)量在孵化前2 d(催青第9 d)達(dá)最高值(0.0253),遠(yuǎn)高于催青第7、8和10 d的Nistari+蠶卵Period基因相對(duì)表達(dá)量。說明Nistari+蠶卵的Period基因在孵化前2 d大量合成,且極顯著高于Nistari-蠶卵,推測(cè)這也是導(dǎo)致Nistari-蠶卵與Nistari+蠶卵孵化率差異的原因之一。
高海青,唐瑩,顧海洋,楊斌. 2019. 影響蠶種孵化率的因素及解決途徑[J]. 江蘇蠶業(yè),41(1-2):15-17. [Gao H Q,Tang Y,Gu H Y,Yang B. 2019. Influence factors on hatching rate of silkworm eggs and its solutions[J]. Jiangsu Sericulture,41(1-2):15-17.]
高頌,劉立會(huì),劉仕平,王偉,陸改,夏慶友. 2012. 家蠶GH18家族幾丁質(zhì)酶的系統(tǒng)進(jìn)化和BmChi的時(shí)期表達(dá)分析[J]. 蠶業(yè)科學(xué),38(3):418-423. [Gao S,Liu L H,Liu S P,Wang W,Lu G,Xia Q Y. 2012. Phylogenetic analysis of GH18 family chitinases and temporal expressional profile of BmChi in the silkworm,Bombyx mori[J]. Science of Sericulture,38(3):418-423.]
黃得純,呂敏芝,馮鑫,黎振杰,張綺瓊,吳日富,甘仕棠. 2020. 東濤雞雞蛋品質(zhì)測(cè)定及相關(guān)性分析[J]. 黑龍江畜牧獸醫(yī),(2):41-44. [Huang D C,Lü M Z,F(xiàn)eng X,Li Z J,Zhang Q Q,Wu R F,Gan S T. 2020. Quality determination and correlation analysis of Dongtao chicken eggs[J]. Heilongjiang Animal Science and Veterinary Medicine,(2):41-44.]
雷芳. 2010. 淺談?dòng)绊懶Q種質(zhì)量的因素及對(duì)策[J]. 四川蠶業(yè),(3):52-53. [Lei F. 2010. Influence factors on the quality of silkworm eggs and its countermeasures[J]. Sichuan Sericulture,(3):52-53.]
李燕飛,李乙,蔣玉蓮,韋紅群,何珊珊,莫云霞. 2018. 廣西現(xiàn)行家蠶品種兩廣二號(hào)原種隨時(shí)孵化技術(shù)[J]. 南方農(nóng)業(yè)學(xué)報(bào),49(4):787-793. [Li Y F,Li Y,Jiang Y L,Wei H Q,He S S,Mo Y X. 2018.Instantaneous hatching technique for silkworm variety Liangguang 2 stock in Guangxi[J]. Journal of Southern Agriculture,49(4):787-793.]
劉振興. 2014. 夜間光照對(duì)黑腹果蠅睡眠和生存能力影響及黑腹果蠅鐘蛋白Timeless與Period相互作用的調(diào)控機(jī)理[D]. 北京:中國農(nóng)業(yè)大學(xué). [Liu Z X. 2014. Effects of light interruption on sleep and viability and interaction between clock proteins Timeless and Period in Drosophila melanogaster[D]. Beijing:China Agricultural University.]
潘明德,席全忠,張波,田超文. 2019. 蠶種孵化的影響因素及應(yīng)對(duì)措施[J]. 現(xiàn)代農(nóng)業(yè)科技,(23):213. [Pan M D,Xi Q Z,Zhang B,Tian C W. 2019. Influence factors on hatching process of silkworm eggs and its countermeasures[J]. Modern Agricultural Science and Technology,(23):213.]
申建梅,陳炳翰,黃丹青,何夢(mèng)琪,胡黎明. 2018. 小菜蛾幾丁質(zhì)酶基因的克隆及表達(dá)分析[J]. 環(huán)境昆蟲學(xué)報(bào),40(1):173-179. [Shen J M,Chen B H,Huang D Q,He M Q,Hu L M. 2018. Cloning and expression profiling of a Chitinase gene from Plutella xylostella[J]. Journal of Environmental Entomology,40(1):173-179.]
束梅影. 2013. 家蠶BmN培養(yǎng)細(xì)胞晝夜節(jié)律生物鐘基因的表達(dá)特征[D]. 蘇州:蘇州大學(xué). [Shu M Y. 2013. Expression characteristics of the circadian clock genes in BmN cell line[D]. Suzhou:Soochow University.]
陶卉,徐世清. 2015. 鱗翅目昆蟲晝夜節(jié)律的授時(shí)機(jī)制研究進(jìn)展[J]. 生命科學(xué),27(11):1355-1363. [Tao H,Xu S Q. 2015. Research progress in the circadian rhythm mechanism of lepidopteron[J]. Chinese Bulletin of Life Scien-ces,27(11):1355-1363.]
王英,司馬楊虎,宋艷,朱曉蘇,徐世清. 2008. 昆蟲生物鐘基因及其分子作用機(jī)制研究進(jìn)展[J]. 江蘇蠶業(yè),30(1):9-14. [Wang Y,Sima Y H,Song Y,Zhu X S,Xu S Q. 2008. Research progress on insect circadian clock genes and their molecular mechanisms[J]. Jiangsu Sericulture,30(1):9-14.]
韋偏,祖盤玉,張福平. 2019. 貴州黃雞蛋品質(zhì)測(cè)定和相關(guān)性分析[J]. 貴州畜牧獸醫(yī),43(2):4-6. [Wei P,Zu P Y,Zhang F P. 2019. Quality determination and correlation analysis of Guizhou yellow eggs[J]. Guizhou Journal of Animal Husbandry and Veterinary Medicine,43(2):4-6.]
張達(dá)燕. 2013. 基于光節(jié)律誘導(dǎo)的家蠶孵化節(jié)律及分子機(jī)制研究[D]. 蘇州:蘇州大學(xué). [Zhang D Y. 2013. Research on hatching rhythm and its molecular mechanism of Bombyx eggs under induction condition of circadian photoentrainment[D]. Suzhou:Soochow University].
張楠楠,檀曉萌,郝二英,賈淑庚,陳輝. 2014. 蛋形指數(shù)對(duì)肉雜雞孵化率的影響[J]. 家禽科學(xué),(8):14-16. [Zhang N N,Tan X M,Hao E Y,Jia S G,Chen H. 2014. Effect of egg shape index on hatching rate of broiler chickens[J]. Poultry Science,(8):14-16.]
張艷. 2020. 亞洲小車蝗的產(chǎn)卵、孵化行為與染菌后蝗蝻體溫響應(yīng)的研究[D]. 蘭州:甘肅農(nóng)業(yè)大學(xué). [Zhang Y. 2020. Study on oviposition,egg hatching behavior of Oedaleus decorus asiaticus and its nymph body temperature response infected by Metarhizium anisopliae[D]. Lanzhou:Gansu Agricultural University.]
趙振華,黎壽豐,黃華云,張靜,丁余榮,薛龍崗. 2010. 蛋重與蛋形指數(shù)對(duì)邵伯雞種蛋孵化性能的影響[J]. 中國家禽,32(19):64-65. [Zhao Z H,Li S F,Huang H Y,Zhang J,Ding Y R,Xue L G. 2010. Effect of egg weight and egg shape index on the hatching performance of Shaobo chic-ken breeding eggs[J]. Chinese Poultry,32(19):64-65.]
朱振鵬,孫曉先,桑丹丹,龔道清. 2013. 蛋重和蛋形指數(shù)對(duì)揚(yáng)州鵝種蛋孵化率的影響[J]. 飼料博覽,(8):6-8. [Zhu Z P,Sun X X,Sang D D,Gong D Q. 2013. Influence of egg weight and egg shape index on egg hatchability of Yangzhou goose[J]. Feed Review,(8):6-8.]
Bowles V M,Young A R,Barker S C. 2008. Metalloproteases and egg-hatching in Pediculus humanus,the body(clothes) louse of humans(Phthiraptera:Insecta)[J]. Parasitology,135(1):125-130.
Lu F H,Tang S M,Shen X J,Wang N,Zhao Q L,Zhang G Z,Guo X J. 2010. Molecular cloning and characterization of hatching enzyme-like gene in the silkworm,Bombyx mori[J]. Molecular Biology Reports,37(3):1175-1182.
Royer V,F(xiàn)raichard S,Bouhin H. 2002. A novel putative insect chitinase with multiple catalytic domains:Hormonal regulation during metamorphosis[J]. The Biochemical Journal,366(3):921-928.
Sauman I,Tsai T,Roca A L,Reppert S M. 1996. Period protein is necessary for circadian control of egg hatching behavior in the silkmoth Antheraea pernyi[J]. Neuron,17(5):901-909.
Tang S M,Wu J,Zhao X H,Wang H Y,Qiu Z Y,Shen X J,Guo X J. 2012. Molecular cloning and characterization of hatching enzyme-like geneII (BmHELII) in the silkworm,Bombyx mori[J]. Biochemical and Biophysical Research Communications,419(2):194-199.
Tao H,LI X,Qiu J F,Liu H J,Zhang D Y,Chu F,Sima Y h,Xu S Q. 2017. The light cycle controls the hatching rhythm in Bombyx mori via negative feedback loop of the circadian oscillator[J]. Archives of Insect Biochemistry & Physiology,96(2). doi:10.1002/arch.21408.
Yasumasu S,Iuchi I,Yamagami K. 2010. cDNAs and the genes of HCE and LCE,two constituents of the medaka hatching enzyme[J]. Development,Growth & Differentiation,36(3):241-250.
Young A R,Mancuso N,Meeusen E N,Bowles V M. 2000. Characterisation of proteases involved in egg hatching of the sheep blowfly,Lucilia cuprina[J]. International Journal for Parasitology,30(8):925-932.
(責(zé)任編輯 蘭宗寶)
