miR-195通過抑制小膠質細胞自噬促進神經術后持續性疼痛的研究
2020-04-10 11:42:04范宗峙馬剛史國棟閆廷飛
實用骨科雜志 2020年3期
范宗峙,馬剛,史國棟,閆廷飛
(1.中國人民解放軍聯勤保障部隊第988醫院骨科,河南 鄭州 450042;2.第二軍醫大學附屬長征醫院骨科,上海 200003)
神經疼痛廣泛發生于神經損傷過程中,多由背根神經節的軸突損傷和異位/自發活動的損傷放電引起[1]。該病發病率較高,在普通人群中的發生率約為7%,中國約有1 600萬人罹患神經疼痛并且療效有限。已有的證據顯示中樞神經系統小膠質細胞的激活在神經疼痛形成和維持中起到重要作用[2],同時小膠質細胞的自噬過程參與其免疫炎癥反應的調節[3]。我們前期研究發現,miR-195在脊髓神經損傷模型中表達上調[4]。基于背根神經節小膠質細胞與中樞神經系統小膠質細胞功能相似[5],我們檢查了脊髓神經結扎模型(spinal nerve ligature model,SNL)術后脊髓小膠質細胞miR-195及自噬水平,以及miR-195模擬物對小膠質細胞的自噬過程及炎癥因子的影響,以初步探究在miR-195大鼠術后持續性疼痛中發揮的作用。
1 資料與方法
1.1 實驗材料
1.1.1 實驗動物 成年雄性SD大鼠購于中國科學院(上海),年齡3~4個月,體重(260±10)g。根據實驗動物研究標準的規定,在恒溫24℃、晝夜交替環境中飼養,自由攝食、飲水。按隨機數字表分成2組(n=18):脊髓神經結扎模型組(SNL組)和假手術組(Sham組)。
1.1.2 主要試劑與儀器 兔源抗LC3多克隆抗體、兔抗大鼠p62多克隆抗體、HRP標記的山羊抗兔IgG(法國Fermentas公司);SYBR Green PCR試劑盒、M-MLV逆轉錄試劑盒(北京天根生化科技公司);Percoll細胞分離液(北京索萊寶科技有限公司);ECL化學發光試劑盒(廣州捷倍斯生物科技有限公司);RIPA細胞裂解液、Trypsin-EDTA細胞消化液,DMEM-H基礎培養基,胎牛血清(美國Gibco公司);牛血清白蛋白,TRIzol reagent(美國Invitrogen公司);異氟醚,miR-195模擬物。細胞濾網(BD Biosciences,Alphen aan de Rijn,The Netherlands)、EPPENDORF低溫高速離心機;iQ5 Bio-Rad iCycler實時檢測系統;TANON凝膠成像分析儀,自動的動態足部觸覺測量器(Ugo Basile model 37400,Milan,Italy)。
1.2 方法
1.2.1 大鼠脊髓神經結扎模型制作 實驗動物給予異氟醚(2.5%)麻醉后,暴露背部脊柱橫截移除左側L6。分離出L5段神經,用4-0的絲線緊密結扎背根神經節的遠端,縫合切口。術后3 d后肌肉注射青霉素預防感染。假手術組模型給予相同的手術處理但是不結扎神經。
1.2.2 脊髓小膠質細胞的分離 術后第1天、第3天、第5天、第10天隨機處死4只大鼠,分離腰部膨大部位。將脊髓在4 mL預冷的Hanks’液中磨碎,采用70 μm的細胞濾網過濾并在400 g離心7 min,轉移上清獲得細胞懸液。15 mL離心管中分別緩慢加入3 mL的75% Percoll、3 mL的50% Percoll,3 mL的5% Percoll及2 mL的PBS。緩慢將細胞懸液加入分離液,10°C條件下1 000 g離心20 min,在50%和75%分界處的細胞即為小膠質細胞。收集后用預冷的PBS清洗,以含有1% BSA的PBS重懸備用。
1.2.3 實時定量逆轉錄PCR檢測miR-195水平 取部分上述小膠質細胞,嚴格按照說明書要求采用TRIzol reagent提取細胞總核糖核酸(ribonucleic acid,RNA),M-MLV逆轉錄試劑盒進行逆轉錄。以U6和β-actin分別作為miR-195和mRNAs的內參,使用SYBR Green PCR試劑盒和Rotor-Gene RG-3000A進行實時定量PCR反應,2-ΔΔCt法進行基因表達水平的定量分析。每個樣本分析3次。
1.2.4 Western blot檢測LC3與p62蛋白的表達 取部分1.2.2所述方法獲得的小膠質細胞,以β-actin為內參進行Western blot檢測。PBS洗滌后加入細胞裂解液于冰上渦旋裂解30 min。15 000 r/min離心裂解物15 min后收集上清液,取30 μL上述樣本分別加入4×SDS-PAGE上樣緩沖液煮沸5 min,上樣至120 g/L SDS-PAGE膠電泳。結束后,將凝膠上的蛋白電轉移至聚偏二氟乙烯(polyvinylidene fluoride,PVDF)膜,以含50 g/L脫脂奶粉的磷酸鹽吐溫緩沖液(phosphate buffered saline tween-20,PBST)封閉3 h。根據分子量標記剪下LC3與p62對應位置的PVDF膜,分別轉移至兔源抗LC3多克隆抗體與兔抗大鼠p62多克隆抗體(1︰1 000),37℃孵育2 h,PBST清洗3次,加入HRP標記的山羊抗兔IgG(1︰4 000),37℃孵育1 h,PBST清洗3次,增強化學發光法(enhanceed chemi-luminescence,ECL)化學發光法顯色曝光。
1.2.5 鞘內給藥與效應分析 如1.2.1所述方法進行分組手術后經腰椎穿刺進行鞘內給藥。將32號的探針插入脊柱的L5和L6之間,手術當天到術后4 d隔日給予大鼠miR-195模擬物以及2-O-甲基化修飾的miR-195抑制劑3 μg。采用自動的動態足部觸覺測量器測試機械刺激縮足反應閾值。最大值為50 g,達到最大值會自動斷開。每個爪子的閾值為3次連續測量的平均值,每次測量的時間間隔至少為10 min。對觸摸痛進行檢測時給予大鼠后爪足底部50 μL丙酮。每個后爪交替給予2次,每次連續給藥的間隔為5 min。丙酮給予2 min后檢測反應。連續檢測3 d后處死大鼠,Percoll密度梯度離心法分離脊髓小膠質細胞,Western Blot檢測LC3、p62及促炎因子白細胞介素-1β(interleukin-1β,IL-1β)、腫瘤壞死因子-α(tumor necrosis factor-α,TNF-α)和誘導型一氧化氮合酶(inducible nitric oxide synthase,iNOS)。

2 結 果
2.1 脊髓神經結扎引起小膠質細胞miR-195表達升高 為探究脊髓神經結扎對大鼠脊髓小膠質細胞miR-195表達的影響,我們建立了大鼠脊髓神經結扎模型,并通過Percoll密度梯度離心法分離脊髓小膠質細胞,采用逆轉錄實時定量PCR檢測miR-195的表達水平。結果表明在術后第1天、第3天、第5天、第10天時SNL組miR-195的表達水平均高于Sham組,并且SNL組內miR-195的表達水平呈現逐漸升高的趨勢,表明脊髓神經結扎是脊髓小膠質細胞miR-195表達的刺激因素(見圖1)。
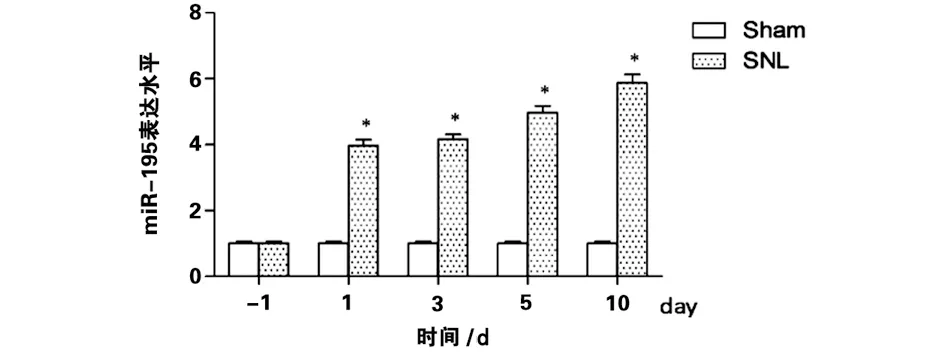
注:*代表與sham組(對照組)相比差異具有統計學意義
2.2 脊髓神經結扎引起小膠質細胞自噬水平降低 為探究脊髓神經結扎對大鼠脊髓小膠質細胞自噬水平的影響,我們檢測了自噬體膜標記蛋白LC3以及多功能泛素結合蛋白p62。LC3Ⅰ酯化為LC3BⅡ后才能錨定于自噬體膜上,故而LC3BⅡ的表達升高是自噬功能活化的標志之一;而p62通過連接LC3和泛素介導蛋白降解,其表達量與自噬水平呈負相關。SNL后的第1天、第3天、第5天、第10天時LC3-Ⅱ/LC3-Ⅰ的表達下調,同時p62表達水平升高,提示SNL后小膠質細胞自噬過程受到抑制(見圖2)。
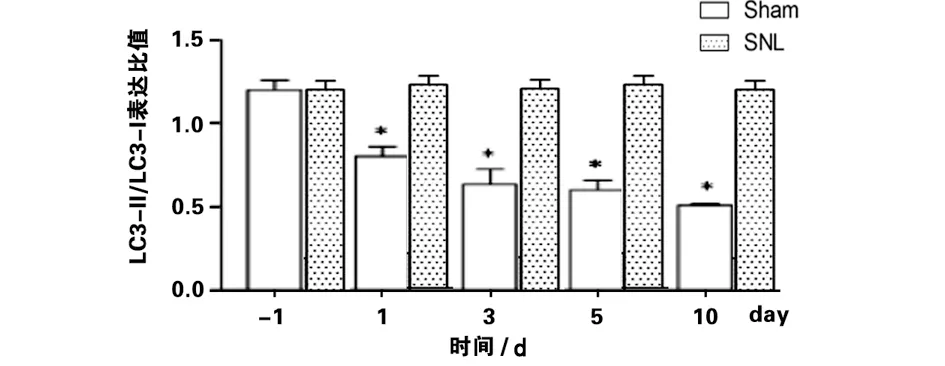
a 自噬體膜標記蛋白LC3-Ⅱ/LC3-Ⅰ表達比值
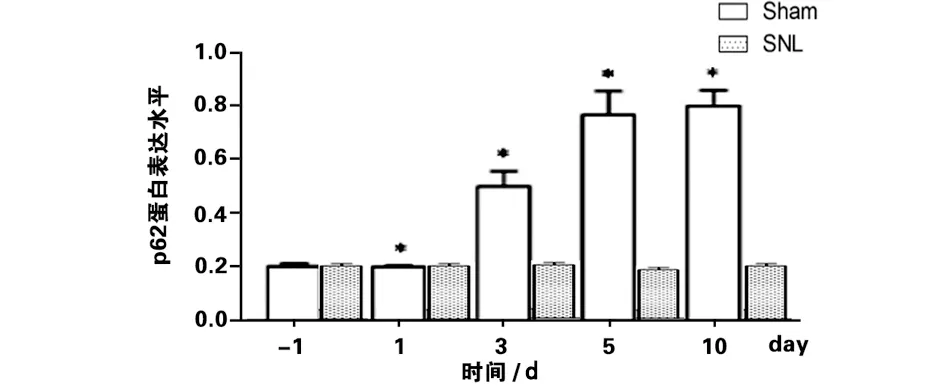
b 多功能泛素結合蛋白p62表達水平
2.3 miR-195的模擬物對大鼠行為的影響 為明確miR-195在脊髓神經結扎引起大鼠神經痛中的作用,我們于在體條件下給予miR-195的模擬物,觀察其對大鼠行為學水平的影響。結果顯示鞘內給予miR-195的模擬物可縮短機械刺激縮足閾值并對熱刺激敏感,且這種效應維持在注射后至少3 d(見圖3)。
2.4 miR-195模擬物引起小膠質細胞炎癥因子增加與自噬抑制 兩組大鼠鞘內給予miR-195的模擬物后,我們采用Western Blot法檢測脊髓小膠質細胞炎癥因子IL-1β,TNF-α和iNOS表達水平及自噬標記蛋白LC3、p62表達水平。結果表明鞘內給予miR-195模擬物誘使大鼠脊髓背角小膠質細胞的炎癥因子表達增加,與此同時LC3-Ⅱ/LC3-Ⅰ的表達下調而p62表達水平升高,提示miR-195模擬物抑制小膠質細胞自噬(見圖4)。

a 機械刺激縮足閾值 b 熱刺激縮足閾值
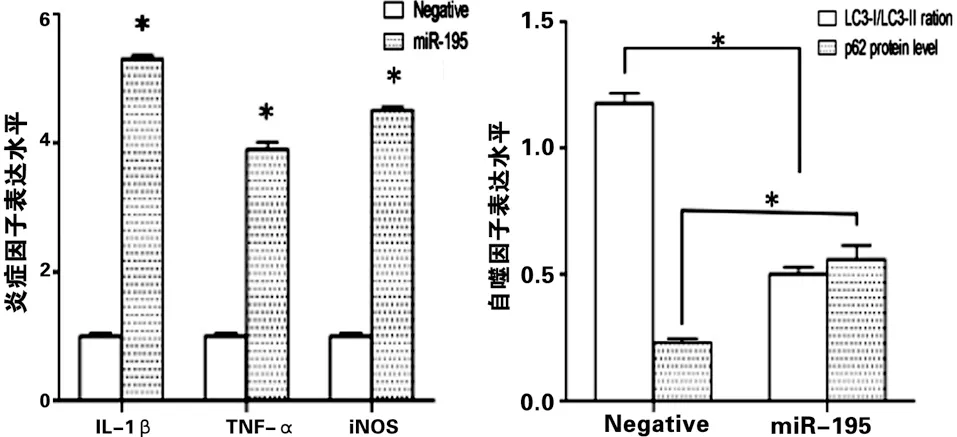
a 炎癥因子表達水平 b 脊髓小膠質細胞的自噬水平
注:*代表與對照組相比差異具有統計學意義
圖4 miR-195的模擬物引起小膠質細胞炎癥因子表達增加與自噬抑制
3 討 論
小膠質細胞是定位于中樞神經系統的巨噬細胞,是脊髓水平炎癥因子的主要來源,而這些促炎因子在神經疼痛的形成和維持中發揮著重要作用[6]。在坐骨神經炎性病變、慢性壓迫性神經損傷、部分坐骨神經結扎和脊神經結扎等各種慢性神經疼痛模型中,痛覺過敏的出現均伴有小膠質細胞的顯著活化[7]。尹顯和等[8]建立的術后持續性疼痛大鼠模型中,脊髓小膠質細胞活化水平于術后3~7 d明顯升高,術后12 d恢復到術前水平。因此我們利用大鼠建立了脊髓神經結扎模型,著力探究脊髓小膠質細胞在持續性疼痛中的分子機制。
近年來的研究表明,miRNAs可能是神經免疫和神經疼痛的重要調節因子[9、10]。Aldrich等[11]發現miR-96,miR-182,miR-183在成年大鼠的背根神經節中高表達,脊髓神經結扎后這些miRNAs的表達大幅度下調,認為它們可能通過轉錄調節疼痛相關基因的表達導致慢性疼痛。Favereaux等[12]則觀察到神經疼痛動物中miR-103表達下降,鞘內給予miR-103可成功的緩解疼痛,提示miRNAs可能成為治療慢性疼痛的新靶點。而我們的前期研究發現,大鼠脊髓神經結扎模型中后脊髓背角miR-195的表達增加,且增加的峰值及持續時間與術后痛覺過敏程度保持一致,提示miR-195可能參與術后持續性疼痛[4]。據此,我們分離了結扎后脊髓小膠質細胞并檢測其miR-195表達水平,結果顯示術后2~14 d miR-195表達水平均顯著高于假手術組并且持續升高,證明脊髓背角miR-195表達的增加主要來源于小膠質細胞。
Zhou等[13]發現缺血腦損傷后通過激活小膠質細胞自噬可抑制免疫炎癥反應,抑制小膠質細胞自噬可增加損傷后炎癥反應。Ma等[14]認為小膠質細胞的自噬有助于其活化和隨后的炎癥反應,然而具體機制仍不明確。基于細胞自噬對于免疫炎癥反應的調節功能,我們采用LC3與p62兩個自噬標志分子作為指標,檢測了脊髓結扎術后小膠質細胞的自噬水平。本研究中,L5脊髓神經結扎后脊髓背角中小膠質細胞自噬水平明顯下降,且持續至術后至少10 d,與miR-195的表達水平呈負相關。為探究miR-195表達升高與自噬抑制的關系,我們采用了miR-195模擬物對術后大鼠進行鞘內注射,并檢測炎癥因子與自噬水平,以及大鼠行為學的變化以衡量神經疼痛程度。有趣的是,miR-195模擬物顯著抑制自噬水平并且促進炎癥因子的表達,與此同時大鼠械刺激縮足閾值降低并對熱刺激敏感,表明miR-195可能通過抑制細胞自噬,從而促進了炎癥因子的表達,參與術后神經疼痛的產生和維持。
總之,本研究初步證明miR-195通過調節小膠質細胞的自噬過程影響其炎癥反應,在神經損傷后持續性疼痛中發揮重要作用,這為研究臨床術后持續性疼痛的治療提供了新的思路和藥物作用靶點。此后,我們將進一步探究miR-195抑制小膠質細胞自噬的機制,以更深入的闡明神經損傷后持續性疼痛的分子機制。
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年12期)2019-05-21 02:55:32
海峽科技與產業(2016年3期)2016-05-17 04:32:12
學苑創造·A版(2015年11期)2016-01-14 09:03:27
