擬南芥核酸酶基因AtCaN2克隆及功能分析
2020-04-16 03:42:24郭坤元張欣欣
華北農(nóng)學(xué)報(bào) 2020年1期
郭坤元,張欣欣
(1.湖北省農(nóng)業(yè)科學(xué)院 中藥材研究所,國(guó)家中藥材產(chǎn)業(yè)技術(shù)體系恩施綜合試驗(yàn)站,湖北 恩施 445000;2.湖北省農(nóng)業(yè)科技創(chuàng)新中心中藥材分中心,湖北 恩施 445000;3.東北林業(yè)大學(xué),鹽堿地生物資源環(huán)境研究中心,黑龍江 哈爾濱 150040)
核酸酶存在于大多數(shù)生命體內(nèi),是一種特異性作用于磷酸二酯鍵的酶,它能夠水解核酸鏈,生成低聚核苷酸或者是單核苷酸[1],它在遺傳物質(zhì)的重組、堿基錯(cuò)配識(shí)別和切割異源DNA等多個(gè)途徑中起著重要的作用[2-6],同時(shí)有的核酸酶活性還依賴于鈣離子、鎂離子、錳離子等各種各樣的金屬離子[7]。
核酸酶在細(xì)菌和微生物里面研究的比較早,一些研究發(fā)現(xiàn),核酸酶可以用于DNA探針進(jìn)行簡(jiǎn)單快速的細(xì)菌檢測(cè),探針熒光信號(hào)與革蘭氏陰性大腸桿菌和革蘭氏陽(yáng)性金黃色葡萄球菌的數(shù)量相關(guān),該方法對(duì)活菌具有特異性,較短時(shí)間即可得到樣本信號(hào)[8];在綠膿桿菌若蟲(chóng)中的研究發(fā)現(xiàn),中腸M1區(qū)域的DNA和RNA降解率高于中腸M3區(qū)域,而中腸各區(qū)域成體核酸酶活性均較低,結(jié)果表明,不同的消化道區(qū)域都具有獨(dú)特的消化特性[9];在鏈球菌中的研究發(fā)現(xiàn),核酸酶Cas9的變異體xCas9具有較高的DNA保真度,這為進(jìn)一步研究和開(kāi)發(fā)高保真聚合酶提供了一定的研究基礎(chǔ)[10];在古細(xì)菌中,一種納米核糖核酸酶Ape0124被發(fā)現(xiàn)和報(bào)道,這種核酸酶是一種3′外切酶,特異性降解小于5個(gè)核苷酸的納米核糖核酸,生化特征研究表明,在錳離子存在條件下,Ape0124可以從3′端開(kāi)始降解ssDNA和ssRNA[11]。在昆蟲(chóng)方面,一些核酸酶也被發(fā)現(xiàn)和報(bào)道,比如在非洲象甲蟲(chóng)的轉(zhuǎn)錄組中發(fā)現(xiàn)了一些假定的dsRNA酶,其中有2種dsRNA酶在昆蟲(chóng)的腸道組織中可以特異表達(dá),RNAi試驗(yàn)證明這2 種dsRNA酶影響了dsRNA在象甲蟲(chóng)腸道中的穩(wěn)定性[12];在東亞飛蝗中,2種核酸酶LmdsRNase1和LmdsRNase4被發(fā)現(xiàn),其活性試驗(yàn)結(jié)果表明,不同生理的pH值可以影響LmdsRNase的酶活性從而改變dsRNA的穩(wěn)定性[13];有的研究對(duì)斜紋夜蛾、東亞飛蝗、美洲大蠊和無(wú)斑飛蝗4種昆蟲(chóng)的dsRNA酶活性進(jìn)行了研究,結(jié)果表明,在適宜的鎂離子濃度和高溫下,所有昆蟲(chóng)的dsRNA酶活性在堿性環(huán)境中均表現(xiàn)出較高的活性,同時(shí)所有被測(cè)昆蟲(chóng)腸道的dsRNA酶活性都比其他組織高幾百倍,這些結(jié)果有助于更全面地了解dsRNA酶的特性,并有助于在昆蟲(chóng)中開(kāi)發(fā)RNAi技術(shù)[14]。一些植物中的核酸酶也被發(fā)現(xiàn)并報(bào)道,比如利用基因芯片技術(shù)在水稻中發(fā)現(xiàn)了一種OsNAC4編碼的參與核DNA降解的核酸酶[15-16];在小麥里發(fā)現(xiàn)了一種可以凋亡細(xì)胞并降解和碎裂細(xì)胞核DNA的核酸酶[17];在花椰菜中,利用DNA-SDS-PAGE的試驗(yàn)技術(shù),通過(guò)對(duì)花椰菜種子,幼苗和花頭不同部位研究,發(fā)現(xiàn)了5種核酸酶[18]。這些研究進(jìn)一步表明核酸酶基因在植物應(yīng)對(duì)自身細(xì)胞凋亡、遺傳物質(zhì)降解及生長(zhǎng)發(fā)育中起著非常重要的作用。
本研究以模式植物擬南芥為研究對(duì)象,通過(guò)PCR擴(kuò)增和克隆測(cè)序方法克隆擬南芥核酸酶AtCaN2基因全序列,分析其序列特征,構(gòu)建其蛋白表達(dá)載體并進(jìn)行蛋白誘導(dǎo)純化及活性分析,以期更全面了解AtCaN2基因,為更詳盡探索其參與調(diào)控的分子機(jī)制提供一定的理論參考。
1 材料和方法
1.1 試驗(yàn)材料
野生型擬南芥(哥倫比亞型)、大腸桿菌 JM109、M15菌株及原核表達(dá)載體pQE-30由湖北省農(nóng)業(yè)科學(xué)院中藥材研究所資源與育種實(shí)驗(yàn)室保存,T4DNA連接酶、限制性內(nèi)切酶等購(gòu)自TaKaRa公司,RNA提取試劑盒、瓊脂糖凝膠回收試劑盒、反轉(zhuǎn)錄試劑盒和質(zhì)粒提取試劑盒等購(gòu)自天根生化科技有限公司,Ni-NTA agarose和尼龍膜等購(gòu)自生工生物工程股份有限公司,檢測(cè)His標(biāo)簽蛋白的一抗和二抗購(gòu)自碧云天生物技術(shù)研究所,其余試劑為國(guó)產(chǎn)分析純。
1.2 試驗(yàn)方法
1.2.1 基因序列克隆 取培養(yǎng)14 d大小的擬南芥材料,在液氮下研磨均勻后,按照RNA提取試劑盒說(shuō)明書(shū)的方法進(jìn)行RNA提取,然后以提取的RNA為模板,按照反轉(zhuǎn)錄試劑盒說(shuō)明書(shū)進(jìn)行反轉(zhuǎn)錄試驗(yàn)。根據(jù)GenBank數(shù)據(jù)庫(kù)中AtCaN2基因序列(序列號(hào):NC_003071.7),以擬南芥cDNA為模版,利用上游引物F:5′-ATGGGTAACGCTCTTACGTT-3′,下游引物R:5′-CCCCTATTTCCTTAGTCTAA-3′進(jìn)行PCR擴(kuò)增,退火溫度為53 ℃,同時(shí)以水為模板作為空白對(duì)照。
1.2.2 序列生物信息學(xué)分析 將獲得的AtCaN2氨基酸序列在GenBank數(shù)據(jù)庫(kù)中進(jìn)行Blast比對(duì)分析,利用DNAMAN等軟件對(duì)其與水稻(NP_001042112.1)、玉米(NP_001147648.1)、谷子(XP_004968301.1)、高粱(XP_002457283.1)、二穗短柄草(XP_003569116.1)、絨毛煙草(XP_009587089.1)、亞麻薺(XP_010508866.1)、大豆(NP_001242780.1)、薺菜(XP_006291485.1)等進(jìn)行同源性分析,并利用MEGA 5軟件構(gòu)建系統(tǒng)進(jìn)化樹(shù)。在線網(wǎng)站包括:預(yù)測(cè)相對(duì)分子質(zhì)量及等電點(diǎn)(http://web.expasy.org/protparam/),疏水性分析(http://web.expasy.org/protscale/),利用TMHMM進(jìn)行跨膜結(jié)構(gòu)域分析(http://www.cbs.dtu.dk/services/TMHMM/),信號(hào)肽分析(http://www.cbs.dtu.dk/services/SignalP),亞細(xì)胞定位分析(http://psort.hgc.jp/form.html),利用SWISS-MODEL(http://swissmodel.expasy.org/)進(jìn)行結(jié)構(gòu)域的三維同源建模。
1.2.3 AtCaN2-GFP蛋白的亞細(xì)胞定位 將AtCaN2-GFP瞬時(shí)表達(dá)載體利用聚乙二醇誘導(dǎo)法[19],轉(zhuǎn)化擬南芥葉片細(xì)胞原生質(zhì)體,原生質(zhì)體先在黑暗條件過(guò)夜培養(yǎng),然后在激光共聚焦顯微鏡下觀察GFP熒光信號(hào)位置。
1.2.4 pQE-30-AtCaN2融合表達(dá)載體的構(gòu)建 根據(jù)目的序列利用引物進(jìn)行擴(kuò)增,上游引物F:5′-GGTACCATGGGTAACGCTCT-3′,下游引物R:5′-GTCGACCTTTCCTTAGTCTA-3′,退火溫度為55 ℃。將PCR產(chǎn)物用SalⅠ和KpnⅠ與載體pQE-30連接,構(gòu)建pQE-30-AtCaN2表達(dá)載體,然后將構(gòu)建好的融合表達(dá)載體轉(zhuǎn)化M15菌株。
1.2.5 pQE-30-AtCaN2點(diǎn)突變載體的構(gòu)建 利用重疊PCR的方法對(duì)AtCaN2氨基酸序列保守區(qū)域中的天冬氨酸位點(diǎn)進(jìn)行點(diǎn)突變擴(kuò)增。第1對(duì)引物的上游引物 F:5′-ATGGGTAACGCTCTTACGTT-3′,下游引物R:5′-ATGGTGTGTCCATGTGCCAC-3′,第2對(duì)引物的上游引物F:5′-GTGGCACATGGACACACCAT-3′,下游引物R:5′-CCCCTATTTCCTTAGTCTAA-3′,然后以擴(kuò)增結(jié)果為模板,利用上一步驟1.2.4的引物進(jìn)行擴(kuò)增,后續(xù)載體構(gòu)建同1.2.4步驟。
1.2.6 融合蛋白的誘導(dǎo)、純化及Western Blot分析 小量誘導(dǎo):挑取含有pQE-30-AtCaN2質(zhì)粒的單克隆于LB液體培養(yǎng)基中振蕩培養(yǎng),待菌液的OD600達(dá)到對(duì)數(shù)中期時(shí)開(kāi)始進(jìn)行融合蛋白的誘導(dǎo),誘導(dǎo)時(shí)間分別為0,15,30,60,120,210 min,誘導(dǎo)結(jié)束后收集每個(gè)時(shí)間點(diǎn)的大腸桿菌進(jìn)行SDS-PAGE電泳;大量誘導(dǎo)及純化:根據(jù)小量誘導(dǎo)結(jié)果,利用三角瓶進(jìn)行大量誘導(dǎo),然后利用溶菌酶裂解菌液,菌液上清過(guò)Ni-NTA 樹(shù)脂進(jìn)行His-AtCaN2融合蛋白的純化,最后進(jìn)行SDS-PAGE電泳及 Western Blot分析。點(diǎn)突變?nèi)诤系鞍椎恼T導(dǎo)及純化參照大量誘導(dǎo)進(jìn)行,純化的融合蛋白用His-AtCaN2(M)表示。
1.2.7 融合蛋白酶活性分析 以λDNA為底物,向反應(yīng)緩沖液(100 mmol/L Tris-HCl pH值 7.0,10 mmol/L CaCl2)中加入1.0 μg His-AtCaN2或者His-AtCaN2(M)融合蛋白和0.5 μg λDNA,37 ℃孵育30 min,同時(shí)準(zhǔn)備對(duì)照試驗(yàn),反應(yīng)緩沖液里面只加入0.5 μg λDNA,37 ℃孵育30 min。孵育結(jié)束后,向反應(yīng)緩沖液里加入金屬螯合劑EDTA使反應(yīng)終止,然后進(jìn)行瓊脂糖凝膠電泳檢測(cè)。另外,將λDNA用HindⅢ酶切反應(yīng)后,進(jìn)行上面相同的試驗(yàn),然后利用瓊脂糖凝膠電泳檢測(cè)結(jié)果。
2 結(jié)果與分析
2.1 基因序列克隆
利用設(shè)計(jì)的引物對(duì)AtCaN2目的條帶進(jìn)行擴(kuò)增,擴(kuò)增結(jié)果如圖1所示,AtCaN2基因片段的大小約為685 bp,證明擴(kuò)增結(jié)果正確。
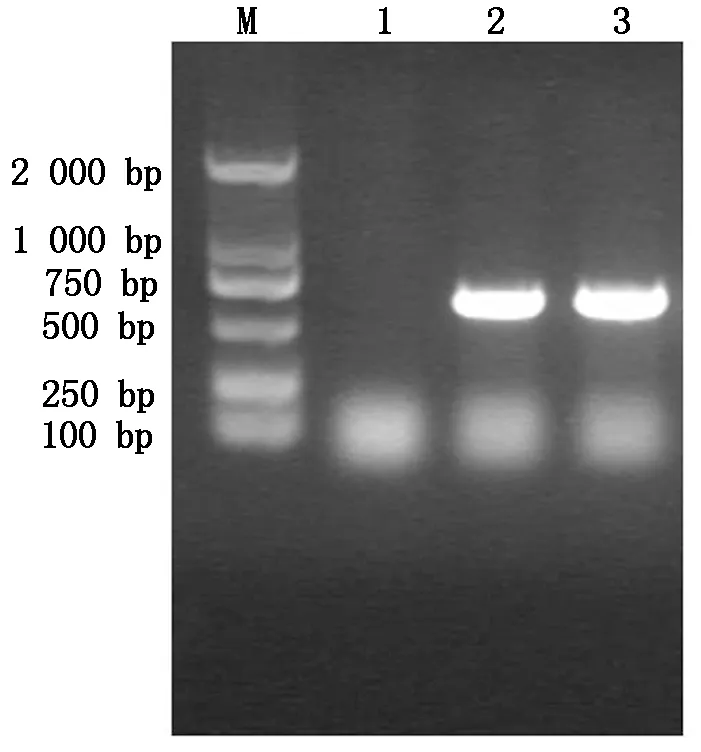
M.DNA Marker;1.空白對(duì)照;2,3.目的基因PCR擴(kuò)增。
2.2 序列生物信息學(xué)分析
利用在線網(wǎng)站(http://web.expasy.org/protparam/)預(yù)測(cè)AtCaN2蛋白的等電點(diǎn)為8.45,化學(xué)式為C1116H1788N310O335S3,共含有3 552個(gè)原子數(shù)。蛋白親/疏水性預(yù)測(cè)結(jié)果顯示,約在第40個(gè)氨基酸位置,疏水性較大,峰值為1.725,在第180個(gè)氨基酸位置,親水性較大,峰值為-2.735,整個(gè)分布區(qū)域親水性氨基酸區(qū)域多于疏水性氨基酸區(qū)域,可認(rèn)為AtCaN2為親水性蛋白(圖2-A)。SignalP 4.1 Server分析結(jié)果顯示,C值為0.143,Y值為0.124,S值為0.146,得分較低,表明AtCaN2蛋白酶沒(méi)有信號(hào)肽(圖2-B)。TMHMM跨膜結(jié)構(gòu)分析表明,AtCaN2不含跨膜結(jié)構(gòu)域(圖2-C)。亞細(xì)胞定位分析表明,AtCaN2蛋白定位于細(xì)胞質(zhì)的概率為0.45,微體的概率為0.3,線粒體基質(zhì)和溶酶體中的概率為0.1,其他細(xì)胞器的概率較低。在其整個(gè)氨基酸序列中,丙氨酸(Ala)、纈氨酸(Val)、亮氨酸(Leu)的含量最高,均超過(guò)了9%,半胱氨酸(Cys)、甲硫氨酸(Met)、色氨酸(Trp)的含量較少,不到1%,其中甲硫氨酸(Met)的含量最少,不到0.5%。蛋白質(zhì)三級(jí)結(jié)構(gòu)預(yù)測(cè)顯示它由2個(gè)連在一起的α-螺旋中心組成(圖2-D)。
2.3 氨基酸序列比對(duì)及系統(tǒng)進(jìn)化樹(shù)分析
本研究將AtCaN2與其他物種中的核酸酶氨基酸序列進(jìn)行了比對(duì),結(jié)果如圖3所示,AtCaN2與亞麻薺(Camelinasativa)的同源性較高,為87.89%,與谷子(Setariaitalic)、高粱(Sorghumbicolor)和玉米(Zeamays)的同源性較低,分別是56.42%,57.35%和56.47%,且序列中間區(qū)域含有一個(gè)高度保守的天冬氨酸D-X-D功能位點(diǎn)序列。
本研究通過(guò)BioEdit、Clustal W和Mega 5等生物軟件對(duì)AtCaN2和其他物種的核酸酶進(jìn)行了系統(tǒng)進(jìn)化樹(shù)分析,如圖4所示:可以發(fā)現(xiàn)AtCaN2與亞麻薺的親緣性最近,與谷子、玉米等禾本科植物的親緣性比較遠(yuǎn),這些結(jié)果也同氨基酸序列比對(duì)的結(jié)果表現(xiàn)出一致性。
2.4 AtCaN2-GFP蛋白的亞細(xì)胞定位
將AtCaN2-GFP融合蛋白表達(dá)載體轉(zhuǎn)化擬南芥葉片原生質(zhì)體,在視野下觀察GFP熒光信號(hào)情況,如圖5所示,AtCaN2-GFP在整個(gè)細(xì)胞質(zhì)均有表達(dá),亞細(xì)胞定位情況與預(yù)測(cè)結(jié)果比較符合。
2.5 融合蛋白的誘導(dǎo)、純化及Western Blot分析
為了檢測(cè)融合蛋白在大腸桿菌中的表達(dá)情況,本研究首先進(jìn)行了小量誘導(dǎo)表達(dá)。經(jīng)過(guò)不同時(shí)間誘導(dǎo)之后,用SDS-PAGE凝膠電泳進(jìn)行檢測(cè),檢測(cè)結(jié)果如圖6所示:在IPTG誘導(dǎo)下,融合蛋白開(kāi)始表達(dá),并且隨著誘導(dǎo)時(shí)間的延長(zhǎng),融合蛋白的表達(dá)量也逐漸增加,同時(shí)結(jié)果還發(fā)現(xiàn)誘導(dǎo)120,210 min融合蛋白的表達(dá)量差別不大,這表明pQE-30-AtCaN2載體構(gòu)建成功,可以成功地進(jìn)行融合蛋白的誘導(dǎo)表達(dá)。
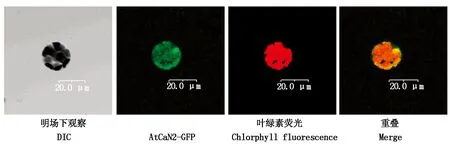
圖5 AtCaN2-GFP亞細(xì)胞定位
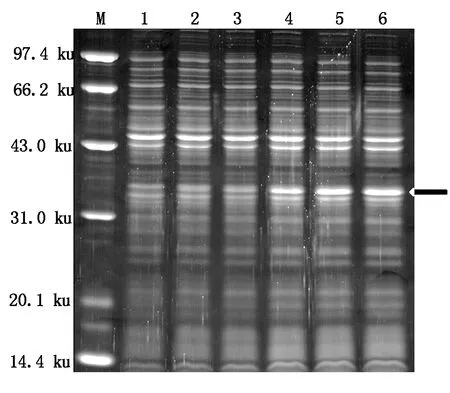
M.蛋白質(zhì)Marker;1-6.誘導(dǎo)0,15,30,60,120,210 min。
根據(jù)小量誘導(dǎo)表達(dá)結(jié)果,本研究對(duì)融合蛋白進(jìn)行大量誘導(dǎo)表達(dá)及純化,結(jié)果如圖7-A所示:得到了大小約為35 ku的純化的His-AtCaN2融合蛋白(泳道2),本研究進(jìn)一步對(duì)His-AtCaN2融合蛋白進(jìn)行了Western Blot分析,如圖7-B所示:檢測(cè)到信號(hào)(泳道4),這些結(jié)果說(shuō)明His-AtCaN2融合蛋白正確翻譯表達(dá),另外純化出來(lái)的蛋白也沒(méi)有出現(xiàn)結(jié)構(gòu)上的斷裂或者降解的情況。
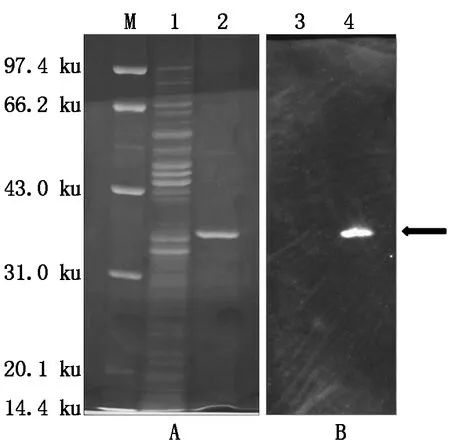
A:M.蛋白質(zhì)Marker;1.未誘導(dǎo)的His-AtCaN2蛋白;2.純化的His-AtCaN2蛋白;B:3,4.Western Blot分析。
2.6 點(diǎn)突變?nèi)诤系鞍椎恼T導(dǎo)和純化
本研究對(duì)保守區(qū)域天冬氨酸位點(diǎn)突變后的His-AtCaN2融合蛋白進(jìn)行誘導(dǎo)和純化,結(jié)果如圖8所示:小量誘導(dǎo)和大量誘導(dǎo)均有點(diǎn)突變后的His-AtCaN2(M)融合蛋白表達(dá),且純化后得到了點(diǎn)突變His-AtCaN2(M)較單一的條帶(箭頭所指位置)。
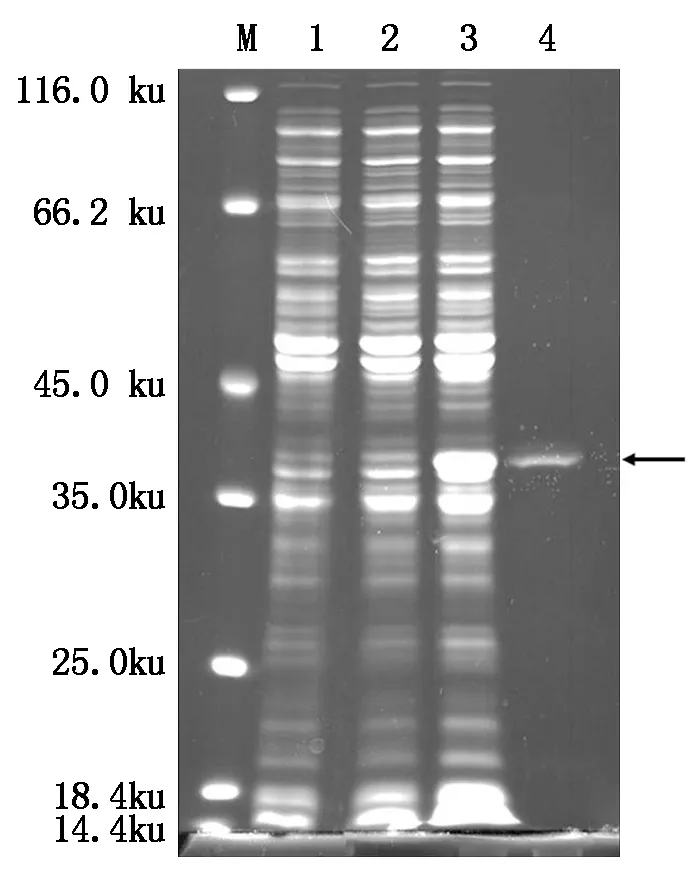
M.蛋白質(zhì)Marker;1.未誘導(dǎo)的蛋白;2.His-AtCaN2(M)融合蛋白小量誘導(dǎo);3.His-AtCaN2(M)融合蛋白大量誘導(dǎo);4.His-AtCaN2(M)融合蛋白純化。
M.Protein Marker;1.Uninduced protein;2.His-AtCaN2(M)fusion protein small quantity induction; 3.His-AtCaN2(M)fusion protein large quantity induction; 4.His-AtCaN2(M)fusion protein purification.
圖8 點(diǎn)突變?nèi)诤系鞍椎恼T導(dǎo)和純化
Fig.8 Induction and purification of pointmutation fusion protein
2.7 融合蛋白酶活性分析
本研究以λDNA為底物,對(duì)核酸酶His-AtCaN2的活性進(jìn)行檢測(cè),由圖9可以看出,在孵育30 min后,添加His-AtCaN2融合蛋白的泳道,底物λDNA大部分降解,核酸酶表現(xiàn)出較高的活性,而添加His-AtCaN2(M)融合蛋白的泳道,底物λDNA基本上未降解,核酸酶表現(xiàn)出沒(méi)有活性(圖9-A);同樣以HindⅢ酶切后的λDNA為底物時(shí),添加His-AtCaN2融合蛋白的泳道,底物大部分降解,核酸酶同樣表現(xiàn)出較高的活性,而添加His-AtCaN2(M)融合蛋白的泳道,底物λDNA基本上未降解(圖9-B)。
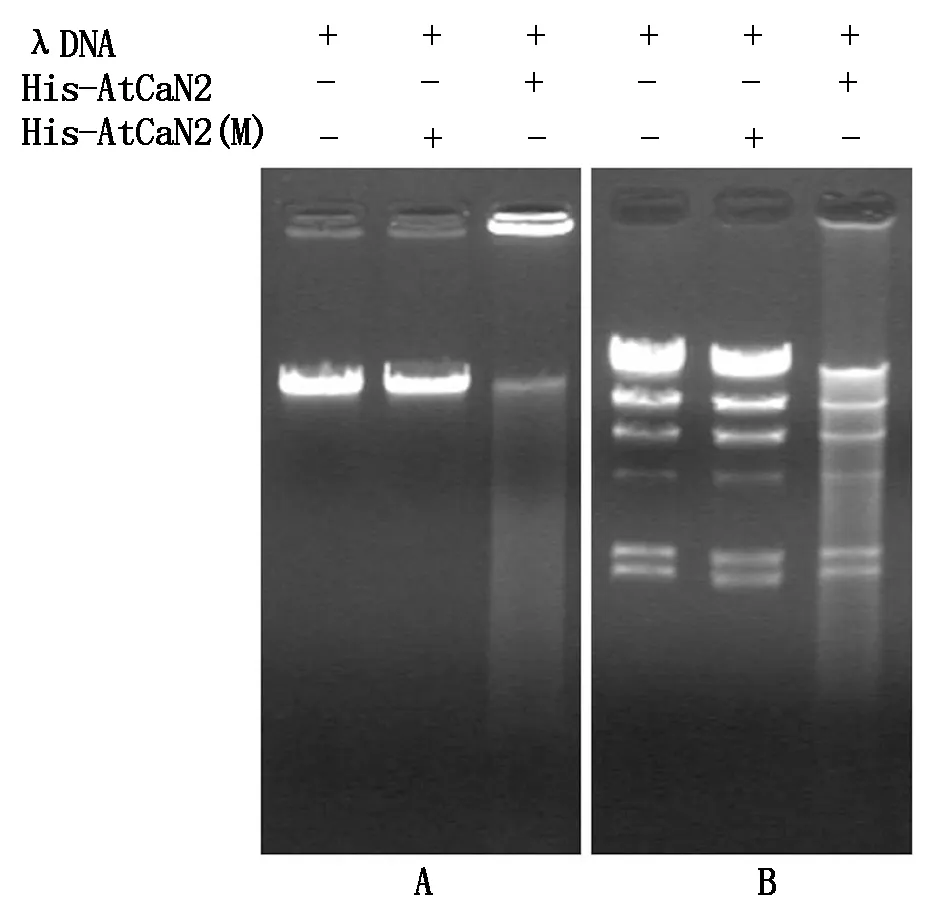
A.底物λDNA;B.HindⅢ酶切反應(yīng)后的λDNA為底物。
3 討論
核酸酶存在于大多數(shù)生命體內(nèi),它在遺傳物質(zhì)修飾、堿基識(shí)別和異源DNA降解等生命過(guò)程中起著非常重要的作用[1-6],同時(shí)一些核酸酶還依靠外源二價(jià)金屬離子來(lái)表現(xiàn)它們的酶活性[7,11,14]。本研究從擬南芥中克隆了一種核酸酶AtCaN2,序列比對(duì)結(jié)果表明,它與亞麻薺的同源性較高,為87.89%,與谷子、高粱和玉米的同源性較低,分別是56.42%,57.35%和56.47%,且序列中間區(qū)域含有一個(gè)高度保守的天冬氨酸D-X-D功能位點(diǎn)序列;預(yù)測(cè)AtCaN2的等電點(diǎn)為8.45,沒(méi)有跨膜結(jié)構(gòu)域,不含信號(hào)肽,是一個(gè)親水性蛋白;蛋白質(zhì)三維結(jié)構(gòu)顯示它是由2個(gè)連在一起的α-螺旋中心組成;進(jìn)化樹(shù)分析表明,AtCaN2與亞麻薺的親緣性最近,與谷子、玉米等禾本科植物的親緣性比較遠(yuǎn);亞細(xì)胞定位結(jié)果表明,其定位于細(xì)胞質(zhì),與預(yù)測(cè)結(jié)果相符合;蛋白誘導(dǎo)和純化得到大小約為35 ku的His-AtCaN2融合蛋白,Western Blot分析表明融合蛋白正確翻譯表達(dá),且沒(méi)有出現(xiàn)結(jié)構(gòu)上的斷裂或者降解的情況;酶活性分析表明,AtCaN2同一些報(bào)道一樣,對(duì)核酸底物具有降解作用[11,15,17,20],且保守的天冬氨酸D-X-D序列是重要的功能位點(diǎn)。總之,AtCaN2在植物中具有重要的功能和調(diào)控作用,本研究結(jié)果為下一步在植物體內(nèi)研究該基因的相關(guān)機(jī)理作用提供了基礎(chǔ)。
猜你喜歡
中學(xué)生數(shù)理化·中考版(2022年8期)2022-06-14 06:55:24
今日農(nóng)業(yè)(2021年19期)2022-01-12 06:16:36
中老年保健(2021年11期)2021-08-22 03:15:44
中學(xué)生數(shù)理化(高中版.高考數(shù)學(xué))(2021年1期)2021-03-19 08:28:38
民用飛機(jī)設(shè)計(jì)與研究(2020年4期)2021-01-21 09:15:02
無(wú)線電工程(2020年11期)2020-10-29 01:25:46
現(xiàn)代出版(2020年3期)2020-06-20 07:10:34
電子制作(2018年18期)2018-11-14 01:48:24
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
中國(guó)中醫(yī)藥現(xiàn)代遠(yuǎn)程教育(2014年11期)2014-08-08 13:23:44
