干旱脅迫和復水處理后梭梭轉錄因子的轉錄組分析
2020-04-16 03:43:54何江峰王力偉房永雨王蘊華劉紅葵
華北農學報 2020年1期
關鍵詞:差異
何江峰,王力偉,2,房永雨,王蘊華,王 朝,劉紅葵
(1.內蒙古自治區農牧業科學院 生物技術研究中心,內蒙古 呼和浩特 010031;2.內蒙古大學 生命科學學院,內蒙古 呼和浩特 010020;3.內蒙古農業大學 生命科學學院,內蒙古 呼和浩特 010010)
梭梭(Haloxylonammodendron)屬黎科(Chenopodiaceae),為超旱生小喬木,呈高大灌木狀,植株高度一般在1~4 m,個別植株可高達8~10 m,亦稱鹽木、瑣瑣樹、梭梭柴。梭梭是肉蓯蓉(CistanchedeserticolaY.C.Ma)的寄主,肉蓯蓉是名貴的中藥材,在醫學上具有抗腫瘤、保肝、補腎、通便、抗老年癡呆、抗輻射等獨特功能,其寄生在梭梭的根端[1-2]。梭梭長期生長在荒漠環境中,形成了抗旱、抗風沙、耐高溫、耐寒、耐鹽堿和耐貧瘠等特性。因此,梭梭是荒漠化地區主要的防風固沙樹種,也可作為駱駝、羊適口性飼料,是荒漠草場中主要的木本飼料樹種之一[3]。梭梭具有重要的經濟價值、生態價值和飼用價值,是我國的二級保護植物,國內外許多學者已從梭梭抗逆的形態學、解剖學、生理學、生態學及分子生物學等方面進行了相關研究。
梭梭的生存環境極其嚴酷,其主要分布地區常年干旱少雨,年降水量不足50 mm,相對濕度低于40%,蒸發量高于降水量的30~100倍,生存環境的極端溫度最高可達40 ℃以上,最低為-40 ℃以下,地表溫度可達70 ℃以上[4]。為了適應極端環境,梭梭逐步形成了自身的極端抗旱機制,但梭梭的抗旱機理、抗旱遺傳機制和抗旱相關組學的研究相對較少,前人研究發現,部分轉錄因子及與干旱脅迫相關,例如,AP2-EREBP轉錄因子(TINY、CBF1、Ptis、AtEBP、DREB1、DREB2)主要調節植物對低溫、干旱及高鹽[5]等的分子應答反應;bZIP轉錄因子在擬南芥基因組、豆科植物基因組、水稻基因組中參與對各種逆境脅迫的響應[6-7];擬南芥中3個NAC基因(ANAC019、ANAC055和ANAC072)的過表達能夠顯著提高植株對干旱的抗性[8];許多NAC基因過表達能夠顯著提高水稻植株的脅迫抗性,卻不會減緩植株的正常生長[9-10];bHLH基因也參與水稻應對干旱脅迫的分子調控過程[11]。在改良植物的抗逆分子育種中,導入或改良轉錄因子較導入或改良個別功能基因在增強植物抗逆性方面更為有效。因此,本研究主要對干旱脅迫和復水處理下梭梭的轉錄因子進行分析,挖掘與干旱相關的轉錄因子,為闡明梭梭抗旱機理奠定理論基礎,保護及開發我國抗逆植物基因資源。
1 材料和方法
1.1 材料及處理條件
梭梭種子收集于內蒙古自治區阿拉善左旗。將梭梭種植在塑料花盆中(直徑:15 cm,高:18 cm),盆內土壤均勻,濕度及質量均一致。梭梭幼苗在植物培養室種植,室內溫度為24/20 ℃(16 h光照 / 8 h 暗處理),相對濕度為30%,光子通量密度為350 μmol/(m2·s)。本研究隨機單株種植25盆梭梭幼苗,包括1個正常澆水組(對照組,CK)、干旱脅迫組(土壤相對含水量為30%~45%,LMS;土壤相對含水量為20%~25%,MWS;土壤相對含水量為10%~15%,SWS)和旱后復水處理組(土壤相對含水量為在重度水分脅迫組上增加10%~15%,RSWS)。干旱脅迫組、復水處理組和對照組的種植均設置5個生物學重復,每天在固定時間測質量、控水、補水并記錄待幼苗生長90 d之后,分別從CK、LWS、MWS、SWS 以及重度脅迫90 d之后復水處理7 d的RSWS的植株上采集等量的根、莖和葉混合,液氮速凍,置于-80 ℃冰箱中儲存備用,提取各處理的總RNA。
1.2 總 RNA 提取
材料總RNA的提取采用 RNAprep Pure Plant Plus KIT(DP441,TIANGEN北京生化科技有限公司)試劑盒。用 DNase酶(2212,大連寶生物工程有限公司)對樣品中的 gDNA 進行消化處理。RNA完整性和純度質量經1.2%非變性瓊脂糖凝膠電泳和 Agilent 2100 分析系統檢測。
1.3 RNA測序和分析方法
總RNA提取按照文獻[12]的方法,利用 Solexa/Illumina進行測序及深圳華大基因公司的Illumina系統軟件進行數據分析。RPKM值用于基因表達水平的分析,利用Cluster軟件[13],以歐氏距離為距離矩陣計算公式,對基因和試驗條件同時進行等級聚類分析,聚類結果用Java Treeview顯示,利用WEGO軟件對差異基因做GO功能分類統計[14],pathway顯著性富集分析以KEGG pathway為單位,應用超幾何檢驗,找出與整個基因組相比后差異表達基因中顯著性富集的pathway基因和基因組的代謝途徑[15]。
1.4 de novo 拼接及序列分析
用Trinity[16]做轉錄組 de novo 組裝,并通過 reads overlap 關系得到 Contig。然后,將 clean reads 比對回Contig,通過paired-end reads 能確定來自同一轉錄本的不同 Contig 以及這些 Contig 之間的距離,Trinity 將 Contig 連在一起,最后得到兩端不能再延長的序列,即為 Unigene。組裝得到的 Unigene,首先使用 Tgicl 將其去冗余并進一步拼接,然后再對這些序列進行同源轉錄本聚類,最終得到非冗余 Unigene。用 Tophat v2.0.9(Broad Institute)將拼接得到的非冗余 Unigene 與已公布的同種屬基因組信息和轉錄組數據進行比對、注釋,用已獲取各測序樣本中包含的轉錄因子編碼基因信息。
1.5 差異表達基因的篩選
樣本中 Unigene 的表達量以公式化校正的 FPKM(Fragments per kb per million fragments)[17]值衡量。2個樣本間差異表達基因(Differential expression genes,DEGs)的篩選及分析使用DEGSeq R package軟件,以FDR(False discovery rate)≤0.001,且Log2(fold change,FC)≥1作為篩選閾值進行差異表達基因的篩選。
1.6 差異表達基因 Real-time PCR 驗證
從測序結果中隨機挑選 9 個轉錄因子編碼基因,用熒光實時定量 PCR 法驗證轉錄組測序數據的有效性和準確性。引物設計用 Primer 5 軟件,序列見表1。以18S為內參基因,用2-ΔΔCt計算,并校正目標基因在不同處理下的表達量,每個基因3次生物學重復。10 μL反應體系中包括5 μL SYBR Premix Ex Taq(TaKaRa,TaKaRa Clontech,Japan),5 μmol/L上游引物,5 μmol/L下游引物和20 ng 模板cDNA。所有反應體系加樣都在96孔板進行,并在Lightcycle-480(羅氏)熒光定量PCR儀上完成,反應程序:95 ℃ 5 min;后按95 ℃ 15 s,55 ℃ 2 min,72 ℃ 1 min,共40個循環;72 ℃延伸2 min。18S基因為擴增體系的內參基因,擴增產物的表達水平按 2-ΔΔCt方法計算[18]。每個樣品都有3次技術重復,結果都以平均值±標準誤(s)(n= 3)計算。
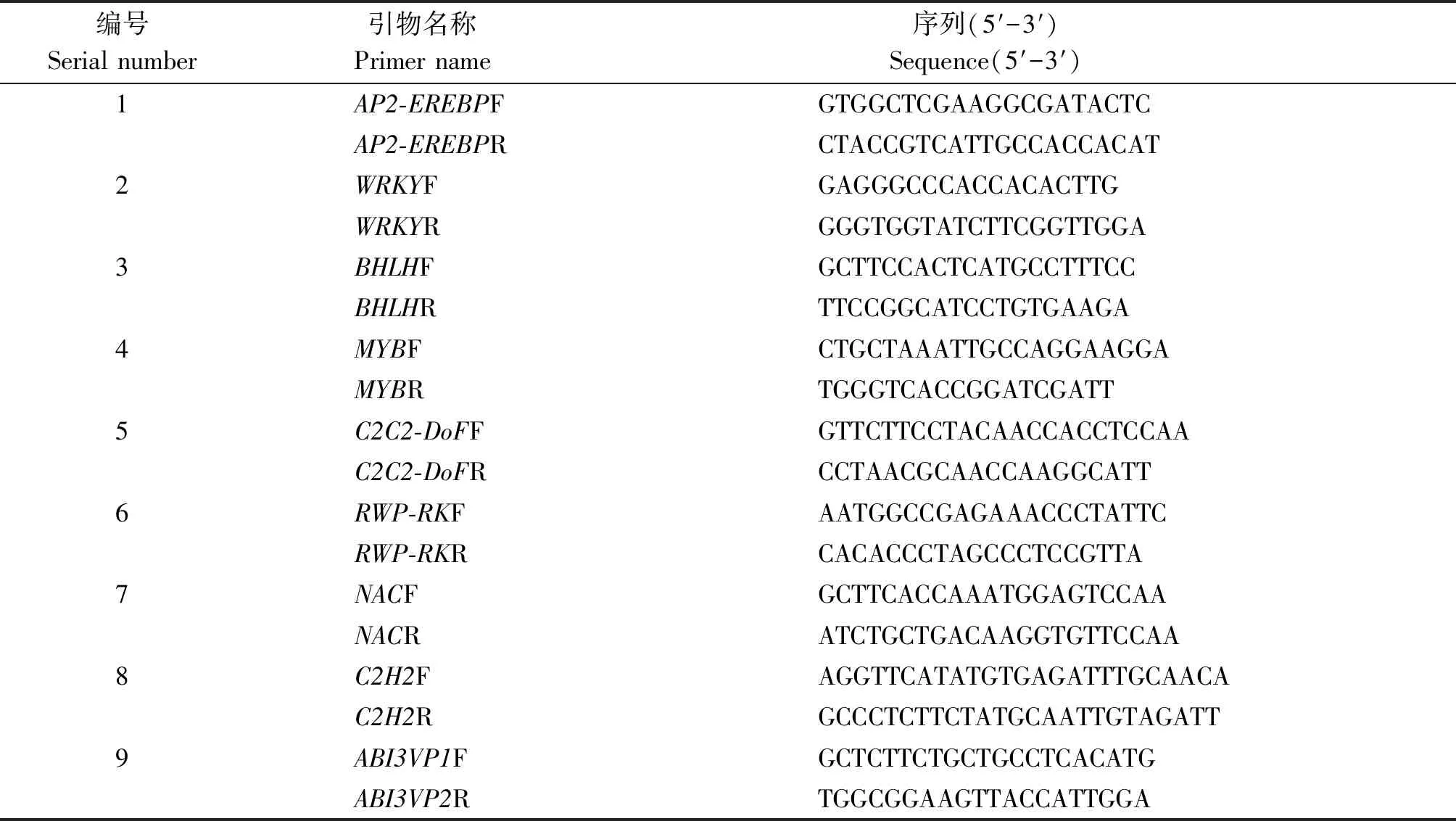
表1 實時定量PCR引物
2 結果與分析
2.1 轉錄組數據統計分析
以梭梭幼苗為材料,對CK、LWS、MWS、SWS和RSWS處理組進行轉錄組測序和表達譜分析。對測序數據進行拼接、比對和整理,經de novo 拼接、去冗余、同源轉錄本聚類后,最終在CK、LWS、MWS、SWS和RSWS 5個組中得到74 641條非冗余Unigene,平均長度780 bp,N50長度1 537 bp。 將CK、LWS、MWS、SWS和RSWS混合樣本拼接得到的非冗余Unigene進行比對、注釋,共檢測到56個轉錄因子家族,1 307個轉錄因子編碼基因;在CK、LWS、MWS、SWS和RSWS組分別檢索到 934,978,994,1 003,991 條轉錄因子編碼基因。LWS、MWS、SWS分別與CK組相比,RSWS與SWS相比,差異表達轉錄因子均以下調表達方式為主,CK-vs-SWS組中下調轉錄因子數目最多,達112條,約占該組差異表達轉錄因子編碼基因總數的59.3%(表2)。
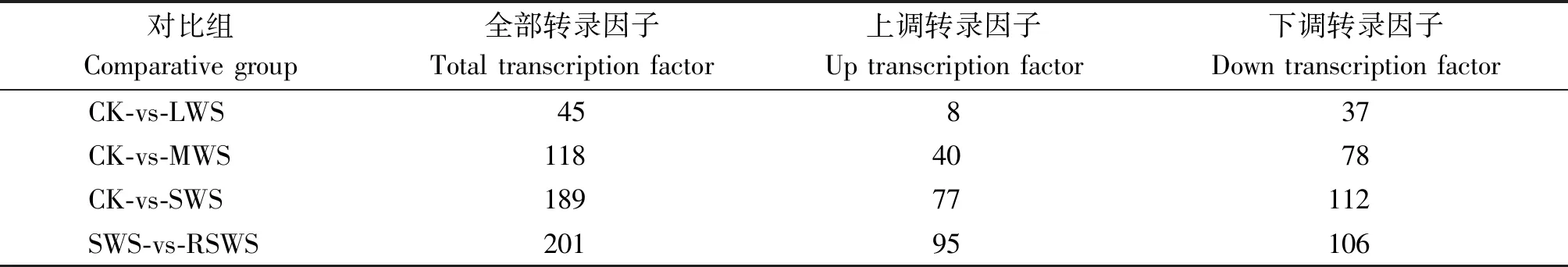
表2 差異表達轉錄因子
2.2 差異表達轉錄因子家族統計分析
獲得4個對比組轉錄因子基本數量信息后,進一步統計分析轉錄因子家族及其包含成員的數量關系。LWS、MWS、SWS分別與CK組對比,RSWS與SWS相比,表3分析表明,其中,AP2-EREBP、WRKY、bHLH、MYB、NAC和ABI3VP1 家族富集的DEGs 超過該組差異表達基因總數的50%以上,其中AP2-EREBP、WRKY、MYB和bHLH家族轉錄因子較多。由圖1-A-C結果顯示,AP2-EREBP家族包含的下調差異表達基因,分別為6個(CK-vs-LWS)、17個(CK-vs-MWS)和 16個(CK-vs-SWS)DEGs,約占各組DEGs總數的75%,81%,59%;WRKY家族包含的下調差異表達基因,分別為6個(CK-vs-LWS)、12個(CKvs-MWS)和 2個(CK-vs-SWS)DEGs,約占各組DEGs總數的100%,100%,33%;bHLH家族包含的下調差異表達基因,分別為5個(CK-vs-LWS)、4個(CK-vs-MWS)和 16 個(CK-vs-SWS)DEGs,約占各組DEGs總數的100%,57%,84%;MYB家族包含的下調差異表達基因,分別為5個(CK-vs-LWS)、9個(CK-vs-MWS)和 10 個(CK-vs-SWS),約占各組DEGs總數的100%,64%,45%。圖1-D結果顯示,AP2-EREBP家族、NAC家族和WRKY家族包含的上調差異表達基因分別占各自DEGs總數的67%,86%,75%。以上表達規律顯示,MYB和WRKY家族基因在LWS組中沒有上調表達,但在MWS和SWS中其上調表達數逐漸增加。綜合說明,RSWS組中偏向通過增加 AP2-EREBP、NAC家族和WRKY家族編碼基因的上調表達數來實現梭梭幼苗對水分刺激的應答調節;而干旱組(LWS、 MWS和 SWS)更偏向于下調AP2-EREBP和bHLH 的DEGs數量來實現其對干旱脅迫的響應。
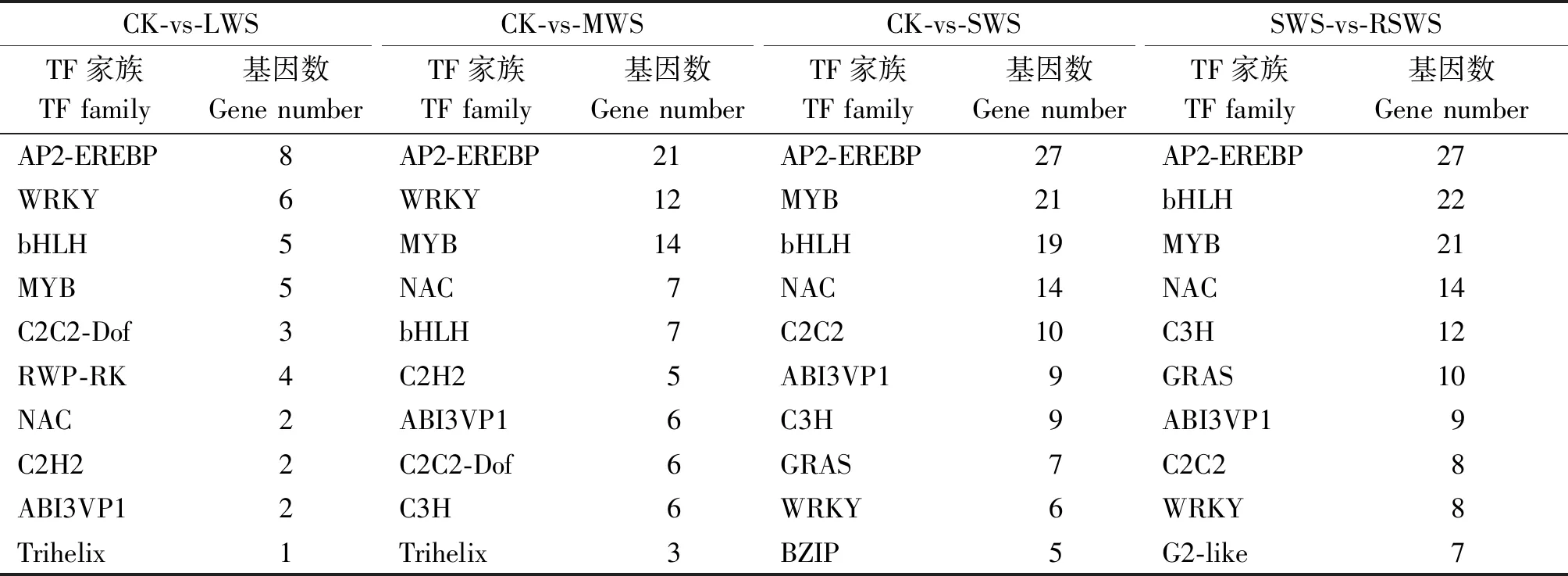
表3 干旱和復水組各轉錄因子家族中的基因表達數
2.3 差異表達轉錄因子表達量分析
從4個對比組中挑選了表達量(FC)變化幅度最大的10個上調和下調基因,并對這些基因的表達規律進行統計分析。結果表明:差異表達基因Tify(CL5087)和NAC(Unigene28108)在干旱脅迫組(LWS、MWS和SWS)和復水組(RSWS)中均上調表達,表達量(FC)變幅為1.23~12.24,2.89~11.37;AP2-EREBP(Unigene2016)、AP2-EREBP(Unigene442)和AP2-EREBP(Unigene160)在干旱脅迫組(LWS、 MWS和SWS)中下調,在復水組(RSWS)中又上調表達。表達量變幅為-4.65~12.22,-8.31~7.38,-6.99~6.85。bHLH(CL2529)和E2F-DP(CL4870)在干旱脅迫組(LWS、 MWS和SWS)中下調表達,變化幅度分別為-14.17~-3.73,-11.89~-4.56;在復水組中卻不表達。RWP-RK(CL8167)、DBP(CL6537),在干旱脅迫組(LWS、MWS和SWS)和復水組(RSWS)中均上調表達。BZIP(Unigene23554)在干旱脅迫組(LWS、 MWS和SWS)和復水組(RSWS)中均下調表達。綜合統計數據結果顯示,AP2-EREBP、NAC、E2F-DP、bHLH 和BZIP等5類轉錄因子對干旱和水分刺激應答顯著(表 4)。
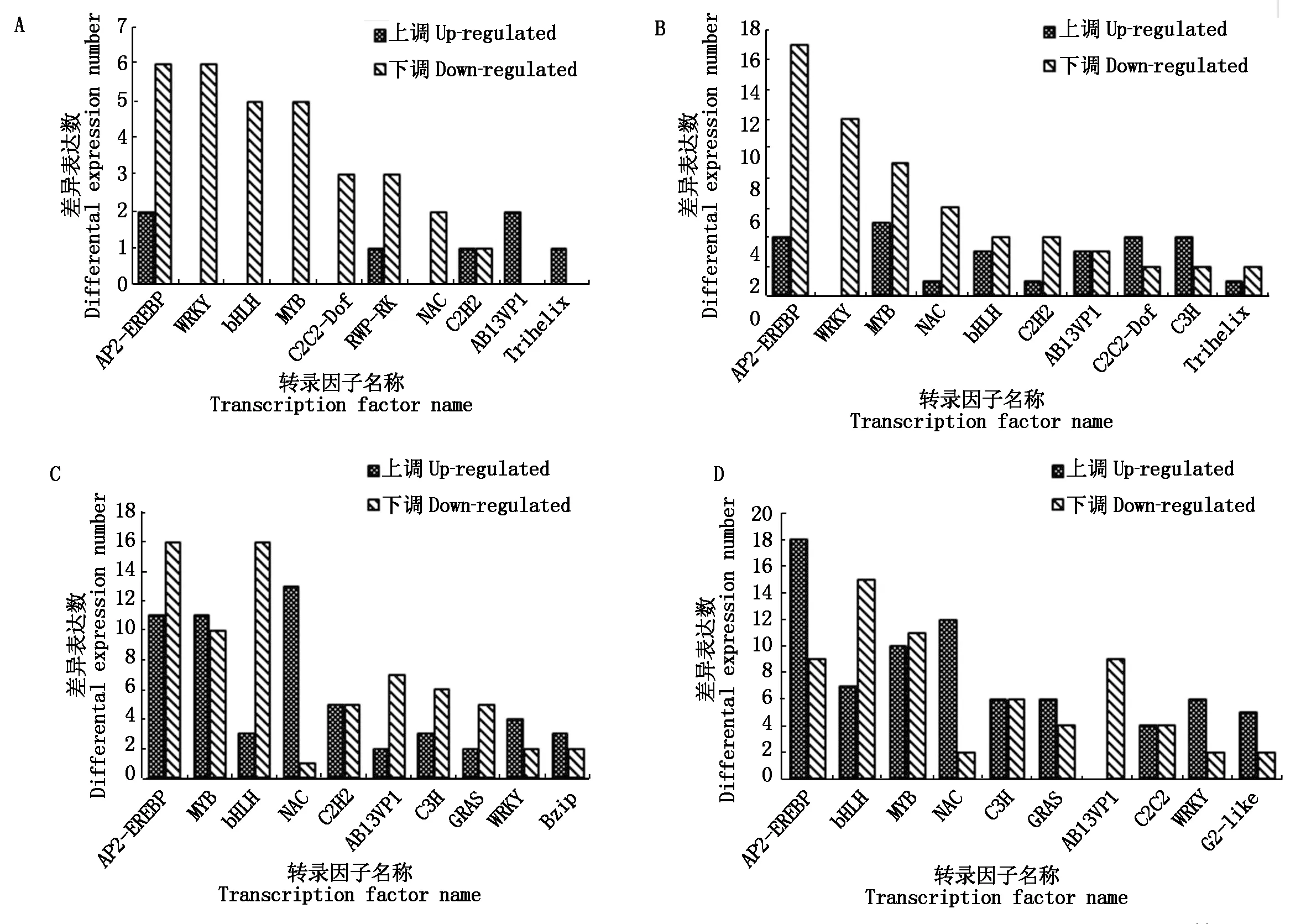
A.CK-vs-LWS的差異表達基因; B.CK-vs-MWS的差異表達基因;C.CK-vs-SWS的差異表達基因; D.RSWS-vs-SWS的差異表達基因。
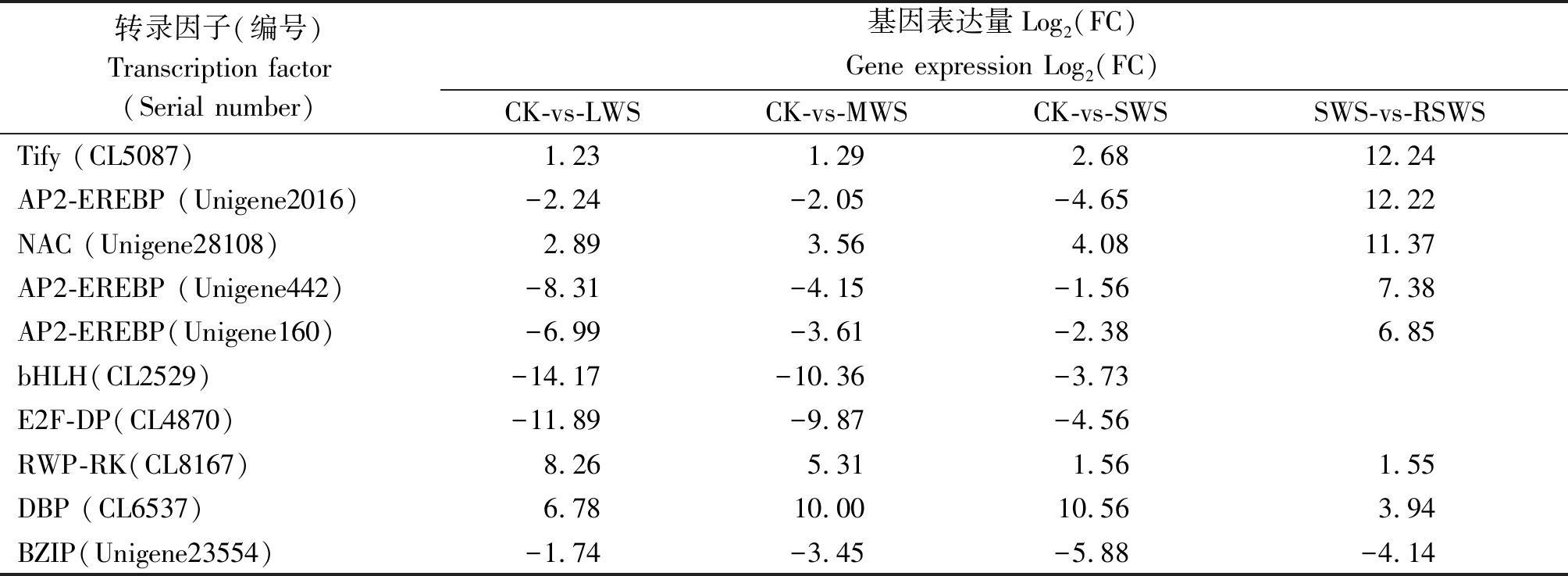
表4 對比組差異表達轉錄因子表達量分析
注:Log2(FC)≥1時,基因表達顯著。
Note:Gene expression is significant when Log2(FC)≥1.
2.4 Real-time PCR 驗證
從測序數據中隨機選取了9個轉錄因子編碼基因,用Real-time PCR 方法驗證測序數據的有效性。圖2顯示,干旱和水分刺激應答相關的AP2-EREBP、WRKY、bHLH、MYB、C2C2-Dof、RWP-RK、NAC、C2H2和ABI3VP1編碼基因在處理前后表達量變化均極顯著。干旱脅迫組和復水組測序數據與用熒光定量PCR 法得到的基因表達數據之間的相關性分別為0.947 8(P<0.01),0.967 2(P<0.01),說明干旱脅迫和復水處理后產生的差異表達基因數據有效。
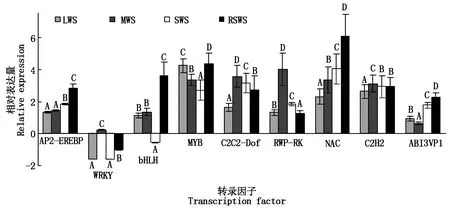
均值±s,樣本均為3個重復;不同大寫字母表示統計結果在1%水平上有顯著差異。
3 討論與結論
本研究通過對干旱脅迫和復水處理下梭梭轉錄因子應答的分子機理進行研究,發掘其中的關鍵轉錄因子,對培育抗旱能力強、水分利用效率高的新作物品種具有重要的意義。研究表明,轉錄因子(Transcription factors,TFs)受脅迫刺激誘導產生,并將信號傳遞、放大,能夠通過調控靶基因轉錄效率從多個層面降低脅迫對植物的傷害,對植物在逆境下的生長發育起到了至關重要的調節作用,尤其是AP2-EREBP、 E2F/DP、MYB、NAC、WRKY、bHLH和BZIP等家族,是信號轉導通路上的關鍵調控因子,對干旱脅迫響應和水分刺激應答發揮了重要的調節作用[19-23]。本試驗測序結果中,各對比組中 AP2-EREBP、MYB、bHLH、NAC、WRKY和ABI3VP1等轉錄因子編碼基因的數量占主要比例,說明這幾類轉錄因子對梭梭干旱脅迫和水分刺激應答調控作用極顯著,其中,AP2-EREBP、WRKY、MYB和bHLH是所有56個家族中差異表達轉錄因子成員最多的蛋白家族。AP2-EREBP轉錄因子要調節植物對低溫、干旱及高鹽[23]等的分子應答反應。本研究中 AP2-EREBP是干旱和復水處理下數量豐富、表達量變化最顯著的一類轉錄因子,且AP2-EREBP(Unigene2016)、AP2-EREBP(Unigene442)和AP2-EREBP(Unigene160),在干旱脅迫下顯著下調,而在復水組中顯著上調,與 bHLH 轉錄因子類似,干旱脅迫處理下誘導更多的下調ERF轉錄因子表達,說明這類轉錄因子發揮調控作用的模式可能與bHLH類似。本研究結果還表明,干旱處理和復水處理后下調的bHLH編碼基因數量均大于上調數,推測bHLH編碼基因主要通過下調其表達調控機體應答脅迫過程。張子佳等[11]對水稻 22 個bHLH基因在PEG 和 ABA 脅迫下的表達譜進行了分析,結果表明,其中一些bHLH基因在響應干旱脅迫過程中受ABA負調控。此外,不同的bHLH基因參與應答環境脅迫的調節方式不同,表明參與 PEG 和 ABA 脅迫應答的bHLH基因具有不同的分子路徑或模式。因此,梭梭中bHLH轉錄因子的進一步研究已在該課題組將繼續進行。
Liao等[24]從研究的大豆基因中獲得了 156 個 MYB 家族基因,利用酵母單雜交的方法篩選出了40個左右與逆境相關的基因,將這些基因轉入擬南芥進行功能驗證,發現轉 GmMYB177 提高了擬南芥耐旱性。丁震乾等[25]在陸地棉中克隆了 MYB 轉錄因子基因GhRAX3,發現其表達在干旱脅迫 0.5 h 后即表現為顯著上調,且 48 h 內持續高表達水平;而抑制GhRAX3的表達,則加快了棉花植株的失水率、細胞質膜過氧化和細胞受損程度,降低了棉花對干旱脅迫的耐受性。目前,發現的大多數 MYB 類轉錄因子基因均表現為提高植物抗旱能力,但亦有例外,如 Zhou 等[26]的研究任務,將JcMYB001轉入擬南芥后,其過表達反而增加了擬南芥對干旱的敏感性。本研究中的MYB轉錄因子在干旱脅迫組與對照組相比,其下調差異表達基因較其他轉錄因子多,MYB家族中上調基因數隨著干旱脅迫程度的增加而增加。說明本研究結果也證實了MYB家族的轉錄因子對干旱脅迫具有正調控和負調控性。
綜合以上結果推測,發揮主要調控作用的轉錄因子家族或成員間可能存在一定的共表達性或相互關聯性,各轉錄因子間協同作用對干旱和水分刺激做出精細應答調控。植物對干旱脅迫和水分刺激應答過程受眾多分子和細胞通路的調控[27]。轉錄因子能特異與順式作用元件結合激活或抑制下游靶基因表達,參與脅迫信號傳遞網絡,其“閥門作用”的重要性不言而喻。本研究借助高通量測序技術,從整體層面概要性的分析了干旱和干旱后復水處理下梭梭幼苗轉錄因子種類及初步的表達規律,結果將為干旱脅迫應答調控網絡、有目的地篩選關鍵調控基因和分子設計育種等研究提供基礎理論數據。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
