4-羥基-3-甲氧基肉桂酸乙酯在擬南芥中的作用位點研究
2020-04-16 03:37:00靳麗宇張利輝
華北農學報 2020年1期
關鍵詞:除草劑
王 穩,靳麗宇,張利輝
(河北農業大學 植物保護學院,河北 保定 071001)
目前,新型除草劑的研發主要依賴于天然除草活性物質[1-2]。在天然源除草劑的研發過程中,以天然除草活性物質為探針發現新靶標已經成為新型除草劑創制的主要途徑[3]。但是,至今已經明確的除草作用靶標僅有20多種,且距離最后一種除草作用靶標HPPD被報道已有30多年[4-5]。因此,探究一種新型除草活性物質的除草作用方式具有重要意義[6-7]。
4-羥基-3-甲氧基肉桂酸乙酯是天然除草活性物質4-羥基-3-甲氧基肉桂酸的酯類衍生物之一,因其具有良好的脂溶性與除草活性且易于合成等優點,已經成為待開發的新型除草活性物質之一[8-9]。4-羥基-3-甲氧基肉桂酸乙酯對常見雜草馬唐的抑制中濃度(IC50)為129.95 mg/L,對馬齒莧芽的IC50為227.05 mg/L,對反枝莧芽的IC50為227.05 mg/L。當用1 000 mg/L的4-羥基-3-甲氧基肉桂酸乙酯莖葉噴施擬南芥24 h后,其葉片表面出現失綠黃化現象[10-13]。
河北農業大學真菌毒素實驗室張明月等[10]前期對4-羥基-3-甲氧基肉桂酸乙酯處理后的擬南芥進行了蛋白水平的分析,結果顯示1 000 mg/L的4-羥基-3-甲氧基肉桂酸乙酯莖葉噴施后的擬南芥中,共有30個差異倍數大于20的蛋白,其中只有差異蛋白gi|15224205|ref|NP_181831.1|和gi|334183674|ref|NP_001185328.1|表達量上調,因此,這2個蛋白在4-羥基-3-甲氧基肉桂酸乙酯的除草過程中起著重要作用。本試驗獲得了蛋白gi|15224205|ref|NP_181831.1|和gi|334183674|ref|NP_001185328.1|的相關基因AT2G43030和AT1G66240的T-DNA插入突變體,通過比較突變體株系與野生型株系對4-羥基-3-甲氧基肉桂酸乙酯的敏感程度,確定擬南芥AT2G43030、AT1G66240基因在4-羥基-3-甲氧基肉桂酸乙酯除草過程中的作用,為探索新的除草劑作用方式奠定了基礎[14]。
1 材料和方法
1.1 試驗材料
擬南芥(Arabidopsisthaliana)哥倫比亞野生型擬南芥(Columbia,Col-0生態型);擬南芥AT2G43030基因的T-DNA插入株系(Columbia,Col-0生態型):SALK_026275、SALK_036415、SALK_027717;擬南芥AT1G66240基因的T-DNA插入株系(Columbia,Col-0生態型):SALK_026221、CS857899、CS856641均由擬南芥信息資源數據庫-tair網站購得。
1.2 擬南芥T-DNA插入純合株系篩選
選取2~3片未抽薹擬南芥植株的蓮座葉片,無菌超純水清洗后放到1.5 mL離心管中,液氮速凍,電鉆勻漿,使用 Plant Genomic DNA Kit(TIAN GEN,中國) 試劑盒提取DNA,并用二引物法驗證純合株系。
PCR反應體系為:DNA模板2 μL,2×PCR Mix 10 μL,ddH2O 6 μL,LB與RP引物各1 μL /LP與RP引物各1 μL。PCR反應程序為:95 ℃預變性5 min;94 ℃變性30 s,55 ℃退火30 s,72 ℃延伸1 min,42個循環最后72 ℃延伸10 min。
二引物法是利用基因T-DNA插入片段的正向引物(LB)和插入位點兩側的基因序列的特異性正向引物和反向引物(LP和RP)進行PCR擴增,該方法需要2組PCR擴增,一組以LP和RP為引物,另一組以LB和RP為引物。在瓊脂糖凝膠電泳檢測PCR產物結果圖中,野生型擬南芥只有LP和RP擴增出1條目的條帶;T-DNA插入雜合株系中,LP和RP、BP和RP共擴增出2條目的條帶;T-DNA插入純和株系只有BP和RP擴增出1條目的條帶。本次試驗所用突變體株系的T-DNA插入位點及其鑒定所用引物序列見表1。

表1 擬南芥AT2G43030和AT1G66240 基因T-DNA插入株系的插入位點及其引物序列
1.3 莖葉噴霧法測定不同株系擬南芥對4-羥基-3-甲氧基肉桂酸乙酯的敏感性
藥劑配制以丙酮∶(含0.1%吐溫80)水=3∶97(V/V)為溶劑,加入化合物4-羥基-3-甲氧基肉桂酸乙酯至終質量濃度為1 000 mg/L(與iTRAQ試驗濃度相對應),使用POTTER噴霧塔噴霧處理7種供試擬南芥,使用前用甲醇和水沖洗體系,每盆噴霧量2 mL,之后放回原溫室中,24 h后進行性狀比對。
1.4 RT-qPCR檢測AT2G43030和AT1G66240基因的表達量
按1.3所述莖葉噴霧處理不同株系的擬南芥植株后,分別在15,30,45 min 3個時間段取樣,每個處理3個重復,設置空白對照。取樣后立即放入液氮中,備用。
將備用樣品電鉆勻漿,使用UNIQ-10柱式總RNA抽提試劑盒與HiScript Ⅱ Q Select RT Super Mix for qPCR(+gDNA wiper)反轉錄試劑盒進行RNA的提取和反轉錄,獲得樣品的cDNA(具體操作見說明書)。無酶水稀釋cDNA濃度至300 ng/μL,使用Trans Start Green qPCR Super Mix試劑盒測定基因的表達水平,本次試驗選用持家基因UBQ作為內參基因。
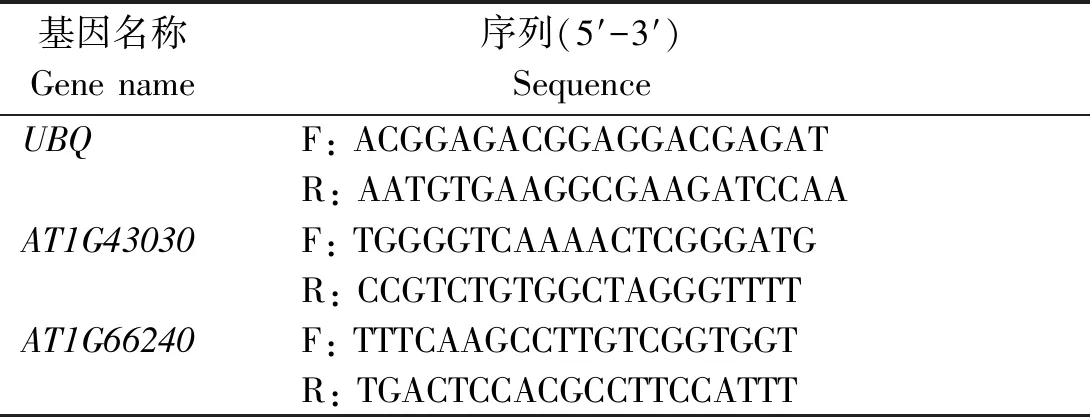
表2 擬南芥AT2G43030和AT1G66240 基因的RT-qPCR引物序列
RT-qPCR反應體系為:cDNA模板1 μL,2×Trans Star?Tip Green qPCR Super Mix 7 μL,ddH2O 1 μL,正向引物(F)和反向引物(R)各0.5 μL。RT-qPCR反應程序:95 ℃預變性30 s;95 ℃變性5 s,55 ℃退火30 s,42個循環。根據NCBI中獲得的差異基因CDS序列設計RT-qPCR所需引物,引物序列見表2。
2 結果與分析
2.1 擬南芥AT2G43030和AT1G66240基因的T-DNA插入純合株系的驗證結果
使用二引物法驗證6種T-DNA插入株系的T3植株是否為純合株系,其驗證結果如圖1所示。在瓊脂糖凝膠電泳檢測圖中,凝膠孔1,3,5,7,9,11表示插入位點兩側的基因序列的特異性引物LP和RP沒有PCR擴增產物;凝膠孔2,4,6,8,10,12表示插入片段的正向引物LB和插入位點右側序列的反向引物RP有PCR擴增產物且大小在400~800 bp,符合插入片段的大小范圍。上述結果說明了6種T-DNA插入株系的T3均為純合株系。
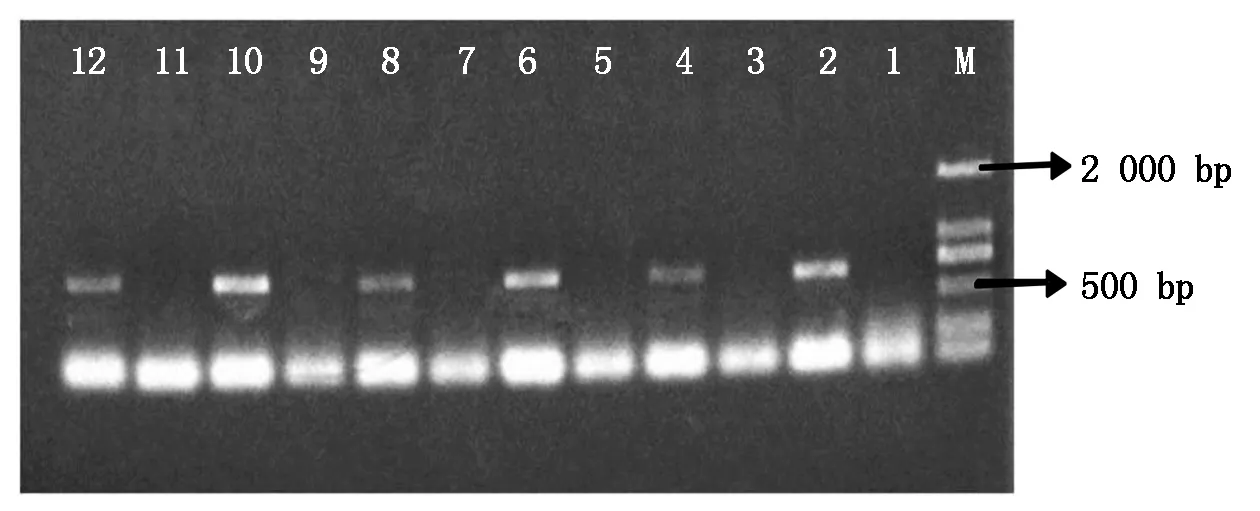
1,2.SALK_026275的純合株系驗證結果;3,4.SALK_036415的純合株系驗證結果;5,6.SALK_027717的純合株系驗證結果;7,8.SALK_026221的純合株系驗證結果;9,10.CS857899的純合株系驗證結果;11,12.CS856641的純合株系驗證結果。凝膠孔下方條帶均為引物二聚體。
1, 2. Verification results of homozygous lines of SALK_026275; 3, 4. Verification results of homozygous lines of SALK_036415; 5, 6.Verification results of homozygous lines of SALK_027717; 7, 8.Verification results of homozygous lines of SALK_026221; 9, 10.Verification results of homozygous strains of CS857899; 11, 12.Verification results of homozygous lines of CS856641. The bands below the gel holes were all primers dimers.
圖1 6種擬南芥T-DNA插入純合株系的驗證
Fig.1 Validation of sixA.thalianaT-DNAinsertion homozygous lines
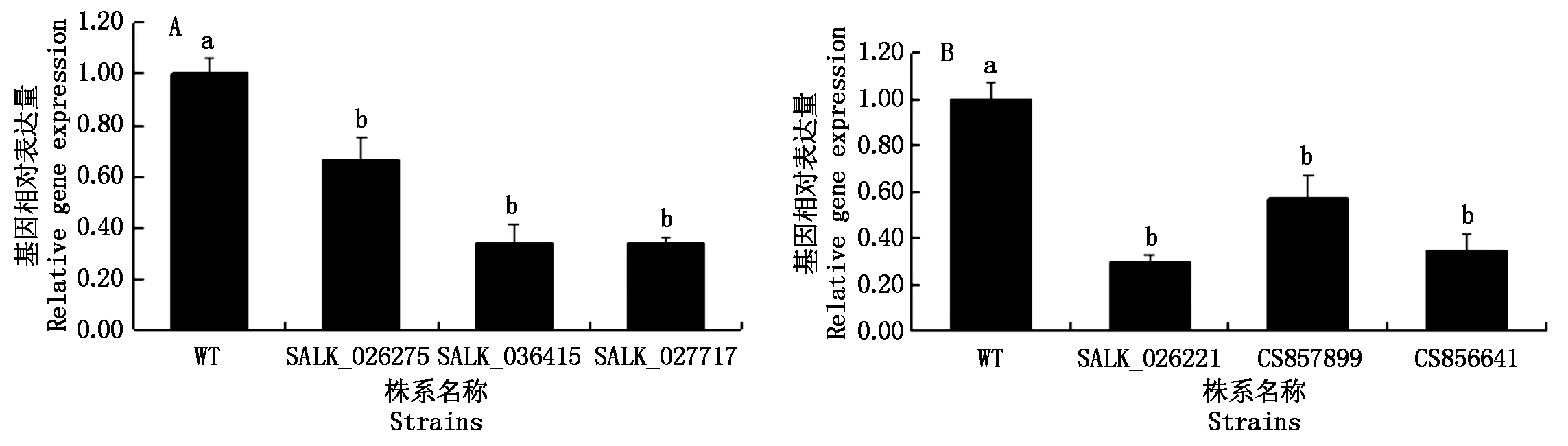
2.2 T-DNA插入純合株系內基因AT2G43030和AT1G66240表達量測定結果
利用q-PCR測定擬南芥AT2G43030和AT1G66240基因的T-DNA插入純合株系中相應基因的表達水平。如圖2所示:在AT2G43030的表達水平測定中當野生型擬南芥(WT)表達量為1.00± 0.06時,SALK_026275株系的相對表達量為0.66±0.08,SALK_036415株系的表達量為0.34±0.07,SALK_027717株系的相對表達量為0.34±0.02;在AT1G66240的表達水平測定中當野生型擬南芥(WT)相對表達量為1.00±0.07時,SALK_026221株系的相對表達量為0.30±0.03,CS857899株系的相對表達量為0.57±0.10,CS856641株系的相對表達量為0.66±0.08。結果表明:與野生型擬南芥相比,6種T-DNA插入純合株系相對應的基因表達量均降低,說明基因上的T-DNA插入會引起基因的失活且6種T-DNA插入純合株系均可用于下一步試驗中對4-羥基-3-甲氧基肉桂酸乙酯敏感性的測定。
圖中小寫字母a、b表示P=0.05水平差異顯著。

Ⅰ.擬南芥的整株受損情況;Ⅱ.擬南芥的第1對真葉受損情況;Ⅲ.擬南芥的蓮座葉受損情況;Ⅳ.擬南芥的莖生葉受損情況。
2.3 不同株系擬南芥對4-羥基-3-甲氧基肉桂酸乙酯的敏感性
1 000 mg/L的4-羥基-3-甲氧基肉桂酸乙酯莖葉噴霧處理擬南芥24 h后,其均出現黃化、皺縮、萎蔫失水等癥狀,但不同株系的受損程度不同。選取相同生長時間、不同株系的同一部位的葉片,在顯微鏡下觀察其癥狀。
如圖3所示,整株擬南芥(圖3-Ⅰ)中,野生型擬南芥(WT)癥狀最輕僅出現葉片黃化、頂端干枯現象;SALK_026275、SALK_036415、SALK_027717株系擬南芥(A1、A2、A3)和SALK_026221、CS857899、CS856641株系擬南芥(B1、B2、B3)均出現頂端干枯、葉片皺縮失水且出現水漬狀潰爛等現象,其中A1癥狀較輕,B3癥狀最嚴重,其原因可能與其插入位點有關。
擬南芥第1對真葉(圖3-Ⅱ)中,WT僅出現皺縮卷曲現象;A1、A2葉尖部分全部干枯,葉片表面失水皺縮,A3的表皮毛明顯增多且出現潰爛現象;B1、B2、B3葉表面嚴重皺縮失水、且出現黃花萎蔫現象。
在擬南芥的蓮座葉(圖3-Ⅲ)中,WT受損現象較輕,出現輕微黃化現象;A1、A2葉片卷曲,A3葉片出現嚴重皺縮失水現象;B1、B2、B3葉片失水皺縮,葉面積變小,但并未出現黃化現象。
在擬南芥的莖生葉(圖3-Ⅳ)中, WT葉柄部位黃化干枯,但葉片仍有一定生命力;A1、A2、A3均黃化、皺縮變形,且表皮毛大量增多;B1、B2、B3卷曲黃化、失水皺縮,表皮毛大量增多。綜合以上現象可知,相比野生型擬南芥(WT),AT2G43030與AT1G66240的T-DNA插入株系(A、B)對4-羥基-3-甲氧基肉桂酸乙酯更為敏感。
2.4 4-羥基-3-甲氧基肉桂酸乙酯處理不同株系的擬南芥后AT2G43030和AT1G66240基因的表達量變化
利用q-PCR測定不同株系的擬南芥體內基因AT2G43030和基因AT1G66240的相對表達量,如圖4所示。使用1 000mg/L的4-羥基-3-甲氧基肉桂酸乙酯莖葉噴霧處理不同株系的擬南芥,15,30,45 min后分別取樣測定基因AT2G43030和基因AT1G66240的相對表達量。野生型擬南芥體內基因AT2G43030和基因AT1G66240的表達量均表現不同幅度的上調,而6種T-DNA插入純合株系相關基因的表達量均未發生變化。綜合2.3中不同株系的擬南芥葉片受損癥狀,可以確定擬南芥T-DNA插入株系對4-羥基-3-甲氧基肉桂酸乙酯更為敏感是由于基因AT2G43030和基因AT1G66240的表達量降低引起的,所以擬南芥基因AT2G43030和基因AT1G66240參與擬南芥對4-羥基-3-甲氧基肉桂酸乙酯的抗逆反應。
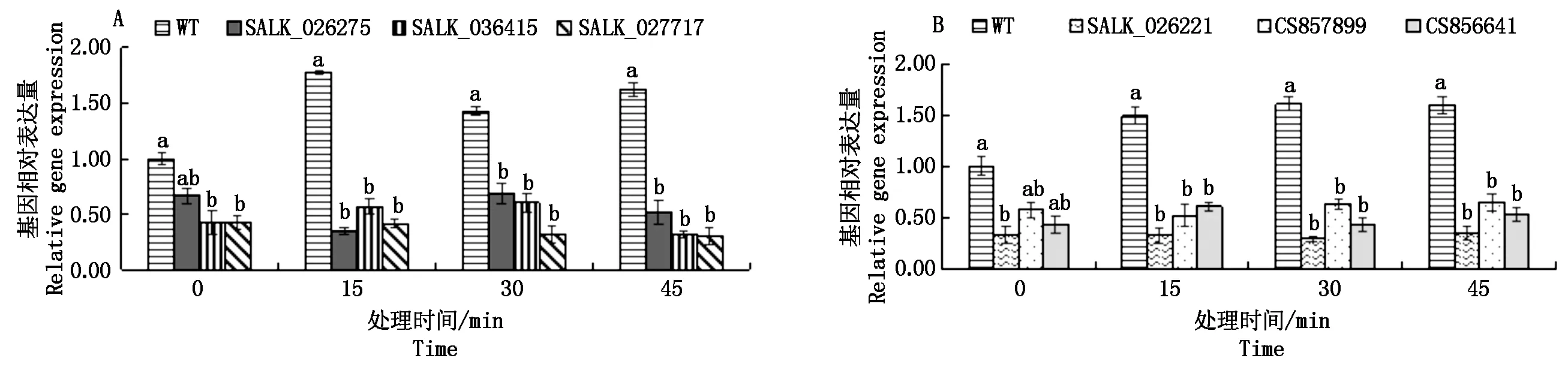
圖4 莖葉噴霧后不同株系的擬南芥在不同時間點基因AT2G43030和基因AT1G66240的表達水平比較
3 討論與結論
本研究中,莖葉噴施4-羥基-3-甲氧基肉桂酸乙酯前后基因AT2G43030和AT1G66240在T-DNA插入株系中的表達水平沒有發生變化,在野生型株系中這2個基因的表達量均上調;觀察莖葉噴施4-羥基-3-甲氧基肉桂酸乙酯24 h后不同株系的葉片損傷程度,與野生型株系相比T-DNA插入株系受損更為嚴重。因此,可以確定AT2G43030和AT1G662402個基因能夠抵御4-羥基-3-甲氧基肉桂酸乙酯對植株造成的傷害。同時,基因AT2G43030和AT1G66240的上調可能是為了彌補擬南芥某些受損基因的功能缺失。基因AT2G43030編碼50S亞單位質體核糖體蛋白,主要存在于葉綠體包膜、葉綠體基質中,與細胞的增殖有關[15];AT1G66240編碼抗氧化劑同系物1蛋白,該蛋白主要存在于擬南芥的細胞膜上,參與擬南芥的銅離子運輸過程[16],說明4-羥基-3-甲氧基肉桂酸乙酯的除草作用機制與植株的光合作用和細胞膜功能有關。
除草活性物質的作用方式及作用位點研究一直是開發新型除草劑的難點,而近些年來隨著蛋白組學、基因組學等技術的進步,以生物信息為指導、從宏觀到微觀的天然產物的作用靶標發現模式逐漸被人們所關注,該模式極大地推進了新型除草劑的研究[17-18]。唐奕教授課題組等運用以抗性基因為導向的基因組挖掘技術成功發現了一種新型天然產物除草劑aspterric acid(AA),同時,成功構建了具有AA耐受性的轉基因作物,為該類型除草劑的開發提供了可能性[19];Wallace 等[20]以DNA回旋酶為藥物靶標,靶向篩選了殺菌劑氟喹諾酮環丙沙星1,后又將其應用于除草劑,靶向合成了更具選擇性的除草活性衍生物;Wright等[21]利用RNA-sep技術測定了除草劑甲氧咪草煙對棘球藻基因表達水平的影響,為咪唑啉酮類除草劑的進一步研究奠定了基礎。本研究以前期蛋白組學的結果為依據,從基因水平解釋4-羥基-3-甲氧基肉桂酸乙酯的除草作用機制,該研究方式為精確闡釋天然活性產物的作用方式提供了新的策略,結合本研究中的天然活性產物及其作用方式研究,可為后續4-羥基-3-甲氧基肉桂酸類天然產物除草劑的發掘提供新的思路與啟示。
猜你喜歡
世界農藥(2019年3期)2019-09-10 07:04:10
今日農業(2019年15期)2019-01-03 12:11:33
現代園藝(2017年19期)2018-01-19 02:50:21
長江蔬菜(2016年10期)2016-12-01 03:05:27
獸醫導刊(2016年12期)2016-05-17 03:51:29
現代農業(2016年5期)2016-02-28 18:42:36
雜草學報(2015年2期)2016-01-04 14:58:05
種業導刊(2016年9期)2016-01-03 01:27:14
營銷界(2015年23期)2015-02-28 22:06:18
營銷界(2015年22期)2015-02-28 22:05:11
