硬頭黃竹花的形態與結構研究
2020-04-16 15:10:58王一方初彩華張月高王曙光
林業科學研究 2020年1期
李 娟,王一方,初彩華,詹 卉,張月高,王曙光*
(1. 西南林業大學 云南省高校林木生物技術重點實驗室 云南 昆明 650224;2. 西南林業大學 生命科學學院 云南 昆明 650224)
竹類植物的繁殖方式多數以無性繁殖為主,在自然條件下大部分竹種需經歷120年或更長的時間才能開花,因而竹類植物的開花現象也極少[1],這導致竹類植物通過生殖器官進行形態學、解剖學、系統分類等方面的研究皆遇到極大的阻礙,尤其是在胚胎學方面的研究更加困難。目前,國內外有關竹類植物花的解剖學已經有少量研究,但僅對少數竹種的生殖生物學進行過詳細描述,如孝順竹(Bambusa multiplex (Lour.) Raeusch. ex Schult.)、綿竹(Bambusa intermedia Hsueh et Yi)、青絲黃竹(Bambusa eutuldoides McClure var. viridi-vittata (W.T. Lin) Chia)、異葉苦竹(Pieioblastus simonii f. heteophyllus Muroi)、綠竹(Dendrocalamopsis oldhami(Munro)Keng)、毛竹(Phyllostachys heterocycla(Carr.) Mitford cv. Pubescens)、鵝毛竹(Shibataea chinensis Nakai)、巨龍竹(Dendrocalamus sinicus Chia et J.L.Sun)、雷竹(Phyllostachys praecox C.D. Chu et C. S. Chao 'Prevernalis')等竹種[2-11]。
硬頭黃竹(Bambusa rigida Keng et Keng f.)屬禾本科(Gramineae)箣竹屬(Bambusa)竹種[12]。主要分布在我國的廣東省、貴州省、四川省、江西贛州等地,其稈通直,材質硬且堅厚,可作竹筷、竹涼席等日用品開發,在用材和造紙方面前景廣闊[13-14]。以往國內外對硬頭黃竹的研究主要集中在形態與理化特性、遺傳多樣性及解剖結構等方面[15-23],缺乏對其具體花器官及胚胎發育方面的細致研究。本研究通過觀察硬頭黃竹花的基本結構,對硬頭黃竹的分類特征描述進行補充,并對硬頭黃竹雌雄配子體發育進行觀察,包括子房形態解剖結構以及雌雄配子體發育過程等,總結其胚胎發育規律,分析硬頭黃竹結實率較低的原因,為硬頭黃竹胚胎學研究積累原始資料,并為硬頭黃竹的育種工作打下基礎。
1 材料與方法
實驗所用材料于2017年4月采自西南林業大學珍稀竹種園。將不同發育階段的小穗和花枝固定于50%FAA固定液中,進行真空抽氣備用。
將固定處理后的小穗使用體視解剖鏡(Olympus HO11)進行不同發育時期的結構解剖、測量及拍照。然后再將解剖后的花藥和子房,浸蠟包埋后,旋轉切片機進行切片,切片厚度為6~8 μm。利用番紅-固綠雙重染色,加拿大樹膠封片[24]。在Nikon-ECLIPSE50顯微鏡下進行觀察和拍照,實驗數據使用Excel 軟件進行分析。
2 結果與分析
2.1 硬頭黃竹花序和小穗的形態結構
硬頭黃竹的花序軸是由不同的節和節間營養枝生長發育而來,小穗生于花枝各節,呈淡黃綠色(圖1A)。小穗底部具有潛伏芽,柄幾無,為假小穗,呈簇狀,多為2個或2個以上長在花序軸的節上(圖1A、B)。底部有3~5片由小變大的近卵形苞片包裹,苞片常發生脫落,苞片背面有紫黑色斑點,苞片一般無毛(圖1C)。小穗含有兩性花3~7朵,平均長3.75 cm,小穗軸節間平均長3.0 mm(圖1D)。開花時,往往花枝同一節的小穗開花程度不同,且同一小穗的不同小花之間開花情況和次序也不同。一般由下往上成熟程度遞減,即基部小花發育成熟最早,頂部小花發育成熟最晚。小穗軸節間清晰可見,后期會發生逐節斷落的現象(圖1E)。小穗軸的節間較扁且長度不同,通常基部節間較短,上部節間較長,且同一節間,下部較細,上部較粗(圖1F)。

圖 1 硬頭黃竹小穗的形態解剖Fig. 1 The morphological anatomy of spikelets in Bambusa rigida
2.2 硬頭黃竹的小花形態結構
硬頭黃竹小花主要由外稃、內稃、漿片、雄蕊和雌蕊五部分組成(圖2)。解剖發現,硬頭黃竹小花包括內外稃片各1枚,外稃質地較硬,平均長1.25 cm,具多脈,頂端急尖,前端邊緣具齒,一般外被纖毛,內側表面光滑(圖2A~D)。內稃對生外稃內側且短于外稃,有2脊,5脈,上部同樣被纖毛(圖2E)。漿片3枚,2枚呈半勺形,另外1枚較其它2枚略長,上部邊緣有纖毛(圖2F)。開花時,漿片吸水膨脹,內外稃張開,此時雄蕊露出小花,因此硬頭黃竹又屬于開放型花。
硬頭黃竹具1枚雌蕊,三分支羽毛狀柱頭,柱頭較長,花柱極短(圖2G)。子房呈長卵圓形,較扁,具明顯三棱,上部具絨毛,下部光滑(圖2H)。雄蕊6枚,未成熟時呈白色,形狀整齊,基部呈箭簇狀,花藥成熟時顏色逐漸變深,且不斷長大(圖2I)。花藥成熟后花絲伸出而垂懸于小花外(圖1A)。

圖 2 硬頭黃竹小花形態解剖Fig. 2 The morphological anatomy of florets in Bambusa rigida
2.3 硬頭黃竹花藥的解剖結構與發育
硬頭黃竹小穗的每朵小花具6枚花藥,每枚花藥具有4個藥室(圖3A)。花藥壁的層數隨著發育時期不同而有所變化。造孢細胞時期花藥壁完全分化,形成四層藥壁細胞,從外向內依次是表皮、藥室內壁、中層和絨氈層(圖3A)。孢原細胞發育為初生造孢細胞后又進行分裂,發育成為次生造孢細胞,此時花藥壁為完整的四層結構(圖3A)。次生造孢組織游離形成小孢子母細胞(圖3B)。在小孢子母細胞時期,絨氈層可能由于營養物質的分泌和消耗,細胞質變淡(圖3B)。接著小孢子母細胞開始減數分裂(圖3C~E),第一次減數分裂過程中發育為二分體,兩個核在同步進行第二次分裂后,會在四個核之間產生細胞壁并且形成四分體,然后形成單核小孢子(圖3F)。此時,在藥室內仍見到處于減數分解期的小孢子母細胞,這表明硬頭黃竹同一花藥室內的不同小孢子發育是不同步的(圖L)。隨著小孢子的體積逐漸增大和細胞質逐漸液泡化,形成許多小液泡,然后小液泡之間互相融合為大液泡,細胞核被大液泡擠到細胞的一端,即單核靠邊期(圖3G),此時可以在花粉壁上觀察到呈凸透鏡狀的花粉萌發孔,絨氈層發生顯著退化,幾近消失,只剩下一薄層貼著藥室內壁,藥室內壁也尚未纖維化形成纖維層。后來液泡逐漸消失,細胞質逐漸充滿整個細胞,細胞核進行一次有絲分裂,形成二細胞花粉粒,藥室內壁也發生顯著纖維化,形成纖維層(圖3H)。花粉粒隨后進一步發生沉積,形成成熟花粉粒,從而導致花粉粒內的細胞核難以觀察到(圖3I)。在花粉粒發育成熟后,藥室壁連接花絲處比較薄,且未發生纖維化,由于纖維層的作用,進而導致此處藥壁縱向裂開,以方便花粉粒釋放。在此階段,花藥壁僅剩下表皮細胞和纖維化的藥室內壁共兩層細胞(圖3I)。

圖 3 硬頭黃竹花藥解剖結構Fig. 3 The anther anatomical structure of Bambusa rigida
2.4 硬頭黃竹花藥敗育
硬頭黃竹雖然開花,但結實率極低,幾無種子可以收獲。通過對花藥進行切片觀察,可以發現花藥出現大量敗育的現象。對這些敗育的花藥進行解剖觀察分析,可歸納總結為以下幾種類型(圖4)。
第一種類型為花藥壁發育正常,能形成正常的纖維層,絨氈層亦能退化,藥室內僅個別花粉粒發育正常,但其他花粉粒發生收縮等異常現象(圖4A)。第二種類型為花藥壁發育異常,未能形成纖維層,藥室內的小孢子幾乎全為空殼花粉粒(圖4B)。第三種類型為花藥壁也未能形成纖維層,花粉粒全部收縮變形,藥室也可能會一起發生收縮(圖4C、D),該類型也可能是第二種類型的不同發育階段。第四種類型為花藥壁能形成絨氈層,且絨氈層發生退化,花藥壁也未發生纖維化,甚至藥室為空藥室,未產生任何小孢子(圖4E)。第五種類型為花藥壁未能夠正常分化,花藥室也未能產生正常的小孢子(圖4F)。
2.5 硬頭黃竹子房的解剖結構
硬頭黃竹極少開花,也未能收獲到發育成熟的果實。在采集硬頭黃竹小穗的時候,由于只有一叢竹子中的幾株開花,因此能夠采集到可用于實驗的小穗不多,加上子房極小,很難切到不同發育時期的雌配子體,因此可以觀察并用以進行描述的代表性切片較少(圖5)。硬頭黃竹子房位于雌蕊下端,子房上部外披較硬的纖毛、下部光滑,且有棱邊。縱切發現該竹種為側膜胎座,倒生胚珠(圖4A),2層珠被(圖4B)。對膨大的子房進行縱切,可偶爾發現發育中的種胚(圖4C)。但由于野外未能收獲發育成熟的果實,也未在竹叢下部的土壤中發現實生苗,因此部分受精成功的子房未能發育成正常種子的原因并不清楚。而與此形成鮮明對比的是,在校園開花慈竹(Neosinocalamus affinis)竹叢林下的土壤中可以觀察到大量存活的實生苗。

圖 4 硬頭黃竹各種發育異常的花藥Fig. 4 Various kinds of abortive anthers in Bambusa rigida
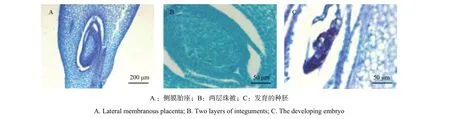
圖 5 硬頭黃竹子房Fig. 5 The ovary of Bambusa rigida
另外觀察發現,開花的硬頭黃竹竹叢中少數竹稈開花,并沒有全部竹稈一起持續開花,開花后相應的竹稈、竹叢也未死去,仍正常生長,也未再次開花。由于未見到其它地理種源的硬頭黃竹開花特征是否與此一致,因此未能明確此特征是否為該竹種的共同特征。但根據杜凡等[25]的總結,將梨藤竹屬(Melocalamus)、空竹屬(Cephalostachyum)、滇竹屬(Oxytenanthera)、思勞竹屬(Schizostachyum)、薄竹屬(Leptocanna)、牡竹屬(Dendrocalamus)、慈竹屬(Neosinocalamus)、懸竹屬(Ampelocalamus)等歸為開花后死亡類型,新小竹屬(Neomicrocalamus Keng f.)、箣竹屬(Bambusa)、大節竹屬(Indosasa)等為開花不死亡類型,箭竹屬(Fargesia)和剛竹屬(Phyllostachys)則開花死亡和開花不死亡兩種情況都有。本研究中硬頭黃竹屬于箣竹屬下的竹種為開花不死亡類型符合該結論。
3 討論
目前在竹類植物研究中,有關竹類植物的花序一般是遵循McClure[26]和耿伯介先生[27]的定義。McClure[26通過研究中國的竹種Schizostachyum的花序,在1934年首次提出“假小穗”的概念,他對竹類植物的花序進行了分類,認為竹子的花序可以分為:determinate inflorescence(有限制花序)和indeterminate inflorescence(無限制花序)兩大類,其區分標準為小穗的基部有無潛伏芽的存在。通常indeterminate inflorescence 的基部苞片內有休眠芽,可以不斷地產生新的小穗狀,此種類型為假小穗;而determinate inflorescence的基部沒有休眠芽,它的各小花是一次性進行開放的。林樹燕等[28]通過對翠竹(Pleioblastus fortunei (Van Houtte ex Munro) Nakai)、黃條金剛竹(Sasaella kongosanensis‘Aureostria-tus’)以及‘霞早’綠竹 (Bambusa oldhamii‘Xia Zao’)的花序建成過程進行了觀察,發現同一個花序中頂部的小穗首先發育,然后向基部依次發育,由不同位置小穗共同組成的花序為有限花序;而同一小穗上不同部位小花的發育順序為基部到頂部的順序依次開放,為無限制花序。因此,林樹燕等[28]認為竹類植物的花序應為混合花序。根據McClure[26]和耿伯介[27]的觀點,黃條金剛竹和翠竹的花序類型屬于有限花序即真花序,根據其定義小穗基部應沒有休眠芽。而林樹燕等[27]發現,黃條金剛竹頂部小穗發育完成后,其基部又新分化出小穗。因此林樹燕等[28]認為竹類植物小穗基部具潛伏芽,應均為有限制花序(假小穗)。本研究中硬頭黃竹小穗采集時間較早,采集時并未關注硬頭黃竹的花序,因此不予討論小穗所組成的花序方式。但由于硬頭黃竹與林樹燕等所報道的綠竹均屬同屬,因此兩者花序應該相同。在硬頭黃竹小穗內,小花由基部向頂部逐漸開放的順序,屬于無限制花序,與林樹燕等人的觀點一致。由于硬頭黃竹小穗基部具有潛伏芽,屬于傳統觀點的“假小穗”。
有關竹類植物花粉敗育、結實率低的報道并不是特別多,王雨珺等[4]在綿竹花藥中發現了大量敗育的現象,并對花藥敗育進行分析,認為花藥缺乏正常的造孢細胞、絨氈層中層發育異常、藥室收縮變形、花粉粒中空、花粉粒皺縮等多種變異類型導致花藥敗育。林樹燕[29]認為花粉敗育及萌發率低、胚囊發育異常、柱頭可授性低及自然授粉率低等情況均引起竹類植物的結實率顯著降低。龐延軍等[30]認為爬竹(Drepanostachyum scandeus (Hsuch et Yi) Keng f. ex Yi)雄蕊發育異常的現象最早出現在雄蕊原基發育時期,發育異常的雄蕊不能形成花粉囊或花粉囊內細胞高度液泡化,無法形成正常的小孢子母細胞。王曙光等[10]認為巨龍竹的小穗中存在營養競爭,致使頂端的小花先發育結實,并先一步成為營養中心,而開花瀕死的巨龍竹本身營養物質有限,進而限制其它小花的發育。除此之外,在青絲黃竹[5]、鵝毛竹[9]中均報道過多種敗育現象,這些敗育現象中與本研究中發現的硬頭黃竹藥室內的小孢子幾乎全為空殼花粉粒、花藥壁未能形成纖維層,花粉粒及藥室收縮變形及花藥壁的絨氈層退化,花藥壁未發生纖維化,甚至藥室為空藥室,未產生任何小孢子等觀察到的敗育現象相似。因此,花粉敗育可能是竹類植物敗育過程中發生共同現象,是竹類植物結實率低的共同原因。
硬頭黃竹的雌雄異熟的特殊條件導致硬頭黃竹結實率降低。雌蕊和雄蕊幾乎都不是同時成熟的,導致硬頭黃竹不能自花授粉,多是異花授粉現象。而異花授粉現象在一定程度上加大了授粉成功的難度,由此降低了硬頭黃竹的結實率。同時,硬頭黃竹的開花季節在雨季,雨季花粉粒沾水后導致其傳粉性能變差,由于其授粉的方式為風媒,因此在一定程度上也降低了硬頭黃竹的授粉幾率。在有限的范圍內,開花的竹叢少,可授粉的雌蕊又不足的情況下,必然會導致硬頭黃竹的結實率下降。
4 結論
硬頭黃竹小穗基本結構發育正常,小穗基部具潛伏芽,為假小穗。小穗開花順序由基部向頂部開放,屬于無限制花序。小穗發育正常,包含3~7朵小花,小花為開放型,具內外稃各1片,漿片3枚,雌蕊1枚,雄蕊6枚。雌蕊為三分枝羽毛狀柱頭,花柱極短。子房長卵圓形,具三棱。子房單室,側膜胎座、倒生胚珠。花粉粒為二細胞花粉,花藥發育后期出現大量敗育現象,可總結為5種敗育類型。花藥敗育是導致硬頭黃竹結實率低的主要原因。
