黑斑側褶蛙蝌蚪斷尾后的補償生長和發育研究
2020-04-20 13:22:46樊曉麗林植華
生態學報 2020年6期
關鍵詞:生長
樊曉麗, 林植華
麗水學院生態學院, 麗水 323000
動物賴以生存的棲息地環境條件通常是可變的。當遭遇食物短缺、低溫脅迫、有毒物質、捕食者等不良條件時,各種動物往往出現暫時的生長停滯或負增長的現象[1- 4]。為了彌補不利環境條件對早期生長發育所產生的消極影響,許多生物在達到特定發育階段之前,往往發生補償生長(Compensatory growth)。所謂補償生長是指動物經過一段時間的生長抑制或其他各種不良環境條件脅迫后,一旦恢復有利生長條件而出現生長加速的現象[5]。根據補償程度的不同,可分為不能補償生長、部分補償生長、完全補償生長和超補償生長四類[6]。具體途徑包括延長幼體生長期[7],提高食物同化率[8],提高生長率[9],改變營養物分配策略[10],降低代謝成本[11],或對不同組織有選擇地分配生長[12]等補償方式,是動物適應自然環境變化而采取的特殊生存策略。昆蟲[12]、魚類[9, 11]、兩棲類[2- 4]、爬行類[10]等生物在一定條件下都會出現補償生長機制。
野外和實驗室內均發現,食蚊魚(Gambusiaaffinis)、克氏原螯蝦(Procambarusclarkiion)、蜻蜓幼蟲(Dragonfly larvae)等水生動物在捕食過程中,往往會造成無尾類蝌蚪的尾不同程度損傷[13- 17]。反捕食理論推測一些無尾類蝌蚪通過引誘捕食者攻擊其彩色明亮尾,而避免對頭部和軀干這兩個身體重要部分的致命攻擊[18- 23]。蝌蚪通過尾鰭損傷可直接獲得生存利益(成功逃避捕食者),但需要付出一定的生長發育代價。一方面,無尾類蝌蚪的游泳能力與其尾部形態特征密切相關,如較長較窄的尾能夠顯著提高蝌蚪的游泳速度[24- 25],當尾損傷嚴重到一定程度(如損傷程度超過30%),就直接造成一定的運動代價(即游泳能力下降),從而使得身體的其他部位更容易受到攻擊,最終降低存活率[13, 19, 26]。另一方面,嚴重的尾損傷可能會導致蝌蚪受傷組織再生能耗的增加,生長受阻,從而使其生長率下降[19, 27]。
捕食風險會隨著時間推移而發生變化,例如,食蚊魚等受口裂限制,蝌蚪個體越大,被捕食的風險越低[17]。補償生長可用來反映生物對早期捕食風險誘導所引起的生長抑制在后續一段時期內的發展變化[11, 28]。因此,確認補償生長的存在是捕食理論和建模的關鍵問題,因為它意味著當捕食風險逐漸消失時,與逃避捕食有關的生長損失可能被抵消。已有的研究表明,補償生長更有可能發生在個體發育經歷棲息地變化的物種中[11, 29],生活史復雜的無尾兩棲類幼體對環境條件變化高度敏感[30- 31],是研究補償生長可塑性的適應意義以及其對生活史的影響的很好模型。
黑斑側褶蛙(Pelophylaxnigromaculatus)隸屬于兩棲綱(Amphibia)無尾目(Anura)蛙科(Ranidae)側褶蛙屬(Pelophylax),是我國的廣布種,通常在每年的3月下旬出蟄,繁殖期為4—6月,主要產卵于稻田、池塘等靜水域的岸邊淺水處[32- 33]。其繁殖水體中共同生活蜻蜓幼蟲、食蚊魚、克氏原螯蝦等各類捕食者,和尾部受傷的黑斑側褶蛙蝌蚪。本研究通過捕食者食蚊魚誘發黑斑側褶蛙蝌蚪形成不同程度斷尾,通過實驗室內分組長期飼養,比較不同程度斷尾組蝌蚪的體長與尾的變化、游泳能力、變態時間與大小等差異,旨在探討:(1) 蝌蚪斷尾損失后的生長軌跡變化,(2) 斷尾損失對蝌蚪游泳能力的影響,(3) 早期階段的斷尾受損對變態時間與大小的影響,(4) 為無尾類蝌蚪的斷尾補償生長機制和種群野外保護提供一定的基礎理論數據。
1 材料與方法
1.1 實驗動物的采集
2014年4月2日雨后清晨,在浙江麗水學院(28°27′N,119°53′E)校園內的持久性池塘中采集3窩黑斑側褶蛙蝌蚪的部分卵團,帶回兩棲爬行動物實驗室,置于塑料箱(700×500×400 mm3,200 mm水深)中自然孵化,待蝌蚪長至能自由游泳的Gosner26- 27期[34]時(Nikon XTS30解剖顯微鏡下鑒定)進行實驗。在同一水體中,用撈網人工捕獲5條食蚊魚帶回實驗室飼養備用。
1.2 實驗設計與管理
2014年4月7日20:00,隨機撈取100只大小相似的黑斑側褶蛙蝌蚪放入同一只塑料箱內,同時放入5條食蚊魚。12 h后,根據斷尾程度將蝌蚪分成全尾組、1/3斷尾組和1/2斷尾組,每組各20只,分別單獨飼養在塑料碗內(口徑120 mm,水深30 mm),每48 h用黑斑側褶蛙粉質飼料按每只蝌蚪體重的20%充足喂食,每周遞增,避免食物水平對蝌蚪發育的影響[2]。每96 h用曝氣24 h后的自來水進行完全換水,保證水位一致,同時按順序調換塑料碗的位置以減少位置形成的溫度差異影響。
每48 h連續跟蹤拍攝測定10次形態。將蝌蚪放入底下有標尺的培養皿中,用Sony DSC-T100數碼相機拍攝其俯視圖,在電腦上用ImageJ 1.44p軟件測定照片中蝌蚪的體長(從吻端到泄殖孔的距離)和尾長(從尾基部至末端的距離)。
以出現兩只前肢(42期)作為蝌蚪的變態標準,及時撈出變態個體,稱取體重,量取體長,測定形態后,單獨放移入塑料碗(保留5 mm水)內,碗口用網孔為1 mm的塑料網罩住,以避免幼蛙逃逸,待尾巴完全消失形成幼蛙(45期),重新測取體重、體長、前肢長、后肢長、頭寬[35]。以實驗開始到蝌蚪出現兩只前肢所經歷的天數作為變態時間。整個實驗在25℃氣侯室(± 0.1℃)內進行,光周期設為12L∶12D。
1.3 蝌蚪游泳能力與局部形態特征的測定
在實驗的第26 d,通過水迷宮視頻跟蹤分析系統(Morris型,安徽正華生物儀器設備有限公司)每組蝌蚪的游泳能力。測試時,先讓每只測試蝌蚪在游泳池內(水溫設為25℃)適應1 min后,再用柳樹枝條輕輕觸碰蝌蚪尾基部使其快速游動,由水迷宮內置軟件記錄蝌蚪在1 min內的疾游速(mm/s)、平均游速(mm/s)和總路程(mm)。
運動測定結束后,測定每只蝌蚪的尾側面局部形態特征。將每只蝌蚪移入外側貼有厘米紙的透明有機玻璃圍成的長方體(80×30×60 mm3)中,拍攝其側面圖后用相同軟件測定出蝌蚪的體長(從吻端到泄殖孔的距離)、體高(軀干最高處)、尾肌高(尾基部的肌肉高度)、尾高(尾部最高處)和尾長(從尾基部至末端的距離)[25]。
1.4 數據分析
用Statistica統計軟件包完成所有數據的統計分析。統計分析前,檢驗數據正態性(Kolmogorov-Smirnov test)和方差同質性(F-max test)。經檢驗,數據無需轉換符合參數統計的條件。用單因素方差分析(One-way ANOVA)、單因素多變量方差分析(One-way MANOVA)及后續的Tukey′s檢驗處理和比較相應的數據。描述性統計值用平均值標準誤(Mean±SE)表示,顯著性水平設置為=0.05。
2 結果
2.1 斷尾后蝌蚪的體長和尾長生長變化
食蚊魚處理后,不同實驗組黑斑側褶蛙蝌蚪的體長和尾長的描述性統計見圖1。One-way ANOVA顯示,3個實驗組間的蝌蚪經歷來自食蚊魚捕食風險后的體長差異顯著(F2, 53=7.33,P<0.002),全尾組和1/3斷尾組蝌蚪的體長顯著大于1/2斷尾組蝌蚪的體長,全尾組和1/3斷尾組之間的蝌蚪體長差異不顯著,全尾組、1/3斷尾組和1/2斷尾組蝌蚪之間形成顯著的尾長梯度差異(F2, 53=403.15,P<0.001)。到第19天時,三個實驗組蝌蚪體長出現反向的顯著差異(F2, 52=4.31,P<0.018),1/2斷尾組蝌蚪體長顯著大于全尾組蝌蚪體長,1/3斷尾組與全尾組和1/2斷尾組之間的蝌蚪體長差異均不顯著。三個實驗組之間蝌蚪尾長的差異不顯著(F2,52=0.19,P=0.828)。

圖1 不同斷尾處理組黑斑側褶蛙蝌蚪的生長變化Fig.1 The growth change of Pelophylax nigromaculata tadpoles from different tail treatments
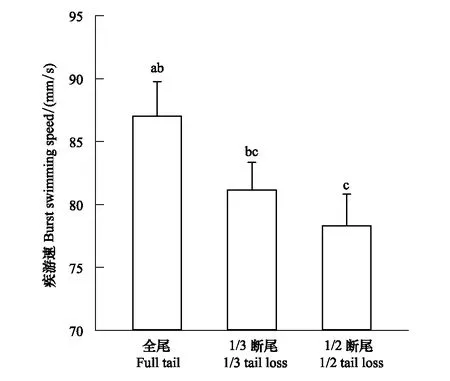
圖2 不同斷尾處理組黑斑側褶蛙蝌蚪的疾游速 Fig.2 Burst swimming speed of Pelophylax nigromaculata tadpoles from different tail treatments
2.2 蝌蚪的游泳速度與形態特征
One-way ANOVA顯示,第26天黑斑側褶蛙蝌蚪的疾游速組間差異顯著(圖2,F2, 52=4.89,P<0.045),全尾組蝌蚪的疾游速顯著大于1/2斷尾組蝌蚪的疾游速,1/3斷尾組蝌蚪的疾游速與全尾組和1/2斷尾組之間差異不顯著。One-way MANOVA顯示,不同實驗組之間的蝌蚪1 min內的游泳總路程、平均游速、蝌蚪的體長、體高、尾肌高、尾高和尾長差異不顯著(Wilks′ λ=0.82,df=8, 98,P=0.283,描述性統計見表1)。
2.3 斷尾處理對變態時間和大小的影響
所有實驗組中黑斑側褶蛙蝌蚪剛變態前后(Gosner 42- 46期)形態特征的描述性統計見表2。One-way MANOVA顯示,實驗組個體在變態時間、變態前后幼體的體重、體長、尾長、前肢長、后肢長、頭寬等差異均不顯著(Wilks′ λ=0.73,df=8, 90,P=0.063)。
3 討論
動物個體發育早期所經歷的環境條件會對后續發育階段的個體適合度和表現產生強烈影響,處于不良環境壓力下的生物如何補償生長對于理解生活史策略的多樣性和進化是至關重要的[3]。通過補償生長,最大限度地減少發育早期所經歷的不良環境壓力對生物體個體產生的負面影響[3]。
3.1 斷尾后黑斑側褶蛙蝌蚪體長和尾長的補償生長響應
本實驗結果表明,在遭遇捕食者捕食斷尾后,全尾組和1/3斷尾組蝌蚪的體長顯著大于1/2斷尾組,而前兩者無明顯差異。造成這一情況,可能是由于1/2斷尾程度造成蝌蚪個體大量失血和組織液流失或者抽樣誤差造成。而1/3的斷尾對于蝌蚪個體而言,可能造成的傷害較小,導致其體長與全尾組沒有顯著差異。已有研究表明,藍尾石龍子(Eumeceselegans)有1/3以上的身體總脂肪含量在尾部,僅次于軀干的脂肪含量,故尾部是其重要的能量儲存器官[36]。當蜥蜴斷尾后主要利用來源于糖原和油脂的能量,先在斷尾截面利用組織液和血液凝結封閉傷口并結痂,隨后啟動尾再生機制,但尾再生會消耗大量的能量[37]。
在蝌蚪恢復生長的第19天時,1/2斷尾組蝌蚪體長顯著大于全尾組蝌蚪體長,出現了明顯的反向差異現象,而1/3斷尾組蝌蚪居中,且靠近1/2斷尾組蝌蚪體長。這表明了經過一段無捕食壓力時期后,這兩組斷尾組蝌蚪的生長率明顯增加,且1/2斷尾組蝌蚪的提高幅度更大些,表明斷尾組蝌蚪發生了補償生長。當捕食風險(食蚊魚)消失后,且食物條件優越,蝌蚪的覓食活動并不受限于時空,大大增加了對其生長的能量投入,可能擁有更高的生長率或消化效率,從而降低了逃避捕食者所帶來的生長代價,提高生長速率[38- 40],這與R.temporaria蝌蚪生長軌跡不同[2]。

表1 黑斑側褶蛙蝌蚪的平均游速與形態特征的描述性統計

表2 黑斑側褶蛙蝌蚪變態前后的形態變化
本研究結果表明,1/2組和1/3斷尾組蝌蚪的尾經過捕食風險移除19天后恢復生長到全尾組蝌蚪的尾長相同長度。蝌蚪迅速地重新長出完整地尾部,對于蝌蚪修復捕食者對其尾部所造成的傷害至關重要[41- 42]。
3.2 斷尾處理對蝌蚪游泳能力及尾局部形態特征的影響
關于無尾兩棲類蝌蚪游泳能力的已有研究表明,蝌蚪通常不擅長于長時間地持續游泳,當逃避捕食者時,往往采取提高瞬時加速度或增加快速轉彎頻次的游泳方式[19, 25, 42- 44]。本實驗結果表明,第19天,不同實驗組蝌蚪的尾長差異不顯著,但1/2斷尾組蝌蚪的疾游速顯著低于全尾組蝌蚪的疾游速。可能是由于斷尾蝌蚪對傷口進行了修復,經過一段時間,長出再生尾,導致蝌蚪疾游速的降低。也有可能是再生尾的形態比原尾有所改變,增大了水流對尾部的阻力,使蝌蚪的游速減小。又或者1/2的斷尾對蝌蚪尾部的神經造成了嚴重傷害,在尾部再生的過程中,神經組織無法完全恢復,從而減弱了尾部的運動功能,降低了疾游速。已有研究表明,石龍子科(Scincidae)蜥蜴在斷尾后運動速度下降19%—35%,鞭尾蜥科(Teiidae)和角蜥科(Phrynosomatidae)物種下降32%—42%,蜥蜴科(Lacertide)部分物種下降28%—48%[36, 45]。閆東娟等人(2015)研究了寬鰭鱲(Zaccoplatypus)和鯽魚(Carassiusauratus)尾鰭缺失后的恢復生長對其快速啟動游泳能力的影響,實驗表明經過20 d的恢復生長,尾鰭面積恢復到切除前的62%—64%,兩者間尾鰭恢復生長的速度差異不顯著;雖然兩種魚尾鰭面積沒有完全恢復,但它們的快速啟動運動能力卻得到了全面恢復[46]。這與本實驗結果有所區別,可能是由于魚類游動時主要依靠尾鰭和身體的擺動。對魚類而言,雖然尾鰭在運動方面起著很大的作用,但身體的擺動作用也不容小覷,故游泳能力有了一定的分配,恢復起來較為容易[46]。本研究結果與關于蝌蚪的已有大多數研究保持一致,蝌蚪的游動主要依靠尾鰭,當尾嚴重受損時,即使恢復原來的形態,在功能上也有了一定的缺失[26]。例如,鎮海林蛙(Ranazhenhaiensis)蝌蚪的游泳速度與尾長度呈正相關,即尾長是鎮海林蛙蝌蚪游泳速度的關鍵性因素,超過1/3程度尾損傷會給鎮海林蛙蝌蚪帶來一定的運動代價[26]。
3.3 斷尾處理不影響變態時間和大小
無尾類生活史中變態時間與大小是其兩個重要適合度指標。越來越多的證據表明,補償生長更有可能發生在個體發育經歷棲息地變化的物種中[29- 30]。因為變溫動物在變態或遷徙時個體變小會造成顯著的死亡率代價[29],較大的變態個體活到成體的機會更多,且繁殖成功率更高[30, 47]。
本研究結果表明,3個實驗組蝌蚪的變態時間差異不顯著,即斷尾后,蝌蚪飼養在消除不利條件的環境下,能與對照組蝌蚪在同一時間內完成變態發育,且變態前后幼體在形態特征上差異均不顯著。這表明當捕食壓力移除后,蝌蚪可通過補償生長消除前期的生長損失對變態時間與大小的持續影響。例如,經歷食物短缺的R.temporaria蝌蚪在恢復低劑量和高劑量飼養后,蝌蚪生長軌跡在變態開始前趨于同一水平,對食物限制所致的生長速率降低表現出完全的代償性生長[2];Fejervaryacancrivora蝌蚪從鹽度脅迫中釋放出來的高鹽度組與低鹽度組蝌蚪擁有相似的變態時間與大小,表現出完全的生長補償[48];瑞典林蛙的北方和南方種群受溫度和食物誘導停止生長后,均可通過延長蝌蚪期表現出追趕式生長(Catch-up growth)響應,變態時的體重與對照組種群相同;追趕式生長是指生物在某一特定生命周期最終達到在沒有經歷限制生長條件下的相同大小[5, 7, 48]。但是,Hector等[7]研究了發育早期的食物限制對褐樹蛙蝌蚪(Litoriaewingii)變態前后的形態與適合度相關行為特征的影響,結果發現,與對照組蝌蚪相比,限定喂食組蝌蚪將延長5天變態,形成較大變態個體。也有學者發現,如果將變態前期的R.temporaria蝌蚪飼養在食物缺乏或高生理濃度的皮質酮環境中,則蝌蚪在發育階段發育緩慢,出現前肢時體重明顯低于對照組的蝌蚪,而在變態后的生長過程中,體重的增加明顯高于對照組蝌蚪,出現追趕式生長的現象[49]。嚴格意義的補償生長是指經過一段時間的限制生長后,通過生理或行為改變直接提高生長率。例如,可通過攝入食物的消化和同化來提高增長率,消化道越長,食物運送時間越長,那么消化效率就會越高[8-9, 38, 50]。
綜上所述,黑斑側褶蛙蝌蚪能夠在變態前調整生長軌跡補償早期捕食風險造成的生長損失,斷尾損失并不影響變態時間與大小,斷尾超過一半的蝌蚪經過補償生長后仍要付出一定的運動代價。
致謝:姜娜同學在野外采集和實驗室飼養及原始數據收集方面提供幫助,特此致謝。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
