油松谷胱苷肽轉(zhuǎn)移酶Tau1單體結(jié)構(gòu)穩(wěn)定性研究
2020-04-21 03:59:22王曉霞楊海靈毛建豐王曉茹
林業(yè)科學(xué)研究 2020年2期
王曉霞,楊海靈,毛建豐,王曉茹
(北京林業(yè)大學(xué)生物科學(xué)與技術(shù)學(xué)院,北京 100083)
谷胱苷肽轉(zhuǎn)移酶(Glutathione S-transferases,GSTs; EC 2.5.1.18)是一類具有多種功能的蛋白質(zhì)超家族。最早發(fā)現(xiàn)GSTs能夠催化還原型谷胱苷肽( GSH)的巰基與異源生物和內(nèi)生生物來源的各種不同結(jié)構(gòu)化合物的親電子基團(tuán)結(jié)合形成R-SG復(fù)合物,增加這些化合物的疏水性,使其易于排出細(xì)胞膜,分解之后排出體外,從而達(dá)到解毒的作用[1];其后發(fā)現(xiàn)GST還具有連接、信號轉(zhuǎn)導(dǎo)等多種生理功能[2],并將具有GSH依賴的轉(zhuǎn)移酶、過氧化物酶、異構(gòu)酶和氧化還原酶以及轉(zhuǎn)運功能的蛋白定義為GST蛋白超家族成員[3]。基于基因的結(jié)構(gòu)、蛋白序列的相似性、關(guān)鍵氨基酸位點的保守性、底物專一性以及免疫交叉反應(yīng)等特征可將GST超家族成員分為多個亞家族[4]。植物 GST可分為10 類:Tau、Phi、Lambda、DHAR、Theta、Zeta、EF1Bγ、TCHQD、Iota和 Hemerythrin[5-7],其中,Tau類GST不僅是植物特有的亞類,也是成員最多的亞類,參與保護(hù)細(xì)胞免受各種生物和非生物脅迫[8-11],以及光信號下細(xì)胞伸長和開花的調(diào)控[12]。
Tau GST都是以二聚體形式發(fā)揮其功能的,單個亞基大約23~30 kDa[13]。每一個亞基都包含獨立的催化部位,包括一個GSH特異結(jié)合位點(G位點),位于蛋白N端及C端結(jié)構(gòu)域形成的V型區(qū)域表面,和一個結(jié)合疏水底物的位點(H位點),主要位于蛋白C端,G位點與H位點由5~10個氨基酸殘基的linker區(qū)連接。G和H位點在結(jié)合和催化底物反應(yīng)時會發(fā)生構(gòu)象變化,具有一定程度的可變性[14]。因此,為了保持GST蛋白催化活性,需要穩(wěn)定G位點和H位點的空間結(jié)構(gòu)。
本課題組前期的研究顯示,裸子植物油松(Pinus tabulaeformis)中一個 Tau類 GST基因(PtGSTU1)在油松不同組織部位均表達(dá),其編碼的蛋白對典型的 GST底物 1-氯-2,4-二硝基苯( CDNB) 和 7-氯 -4-硝 基 苯 并 -2-氧 雜 -1,3-二 唑(NBD-Cl)具有高活性和較高的親和力,其蛋白的Ser13、Lys40、Ile54、Glu66和Ser67為谷胱苷肽結(jié)合(G-位點)殘基,對底物特異性和GSH的親和力具有重要作用,同時這些位點的替換能夠影響蛋白的穩(wěn)定性[15-16]。本研究中,筆者進(jìn)一步探討PtGSTU1的單體穩(wěn)定性,根據(jù)蛋白三維結(jié)構(gòu)模擬預(yù)測出在單體結(jié)構(gòu)穩(wěn)定性中具有關(guān)鍵作用的氨基酸位點,利用定點突變對這些位點進(jìn)行氨基酸分子替換,通過酶學(xué)功能檢測,解析這些位點對蛋白穩(wěn)定性的影響。
1 材料和方法
1.1 結(jié)構(gòu)模擬
本研究選取了一個油松Tau類谷胱苷肽轉(zhuǎn)移酶為研究對象,利用PtGSTU1[16]的蛋白序列(GenBank No. AAT69969)進(jìn)行結(jié)構(gòu)模擬。利用PtGSTU1蛋白序列在 Protein Data Bank(https://www.rcsb.org/)數(shù)據(jù)庫中搜索結(jié)構(gòu)模板,使用默認(rèn)參數(shù)利用SwissModel[17]進(jìn)行結(jié)構(gòu)模擬。
1.2 構(gòu)建PtGSTU1突變體
以含有PtGSTU1編碼序列的重組載體為模板,通過PCR進(jìn)行定點突變。對于Arg 18突變體,利用誘變正向引物(R18X -F)和反向引物(PtGSTU1-R)進(jìn)行PCR擴增。PCR反應(yīng)混合物含有10 pmol的每種引物,1 U的pfx Taq DNA聚合酶(Invitrogen Life Technologies,USA),0.3 mmol·L-1的各種 dNTP(Amersham Pharmacia Biotech,USA),1 mmol·L-1MgSO4和 1~3 ng 的模板 DNA,反應(yīng)體系為50 μL。優(yōu)化的PCR條件為:94℃下變性2 min,94℃ 15 s,55℃退火30 s和68℃延伸60 s,反應(yīng)30個循環(huán),最終延伸2 min。對于103位突變體,進(jìn)行兩輪PCR,第一輪PCR分別用正向引物(PtGSTU1-F)和誘變反向引物(D103A-R)、反向引物(PtGSTU1-R)和誘變正向引物(D103AF)分別進(jìn)行擴增,獲得2個片段。用GFX PCR DNA and Gel Band Purification Kit(Amersham Pharmacia Biotech,USA)純化第一輪反應(yīng)產(chǎn)物。第一輪PCR產(chǎn)物在目的突變位點產(chǎn)生短片段重疊區(qū)段。第二輪PCR使用正向(PtGSTU1-F)和反向(PtGSTU1-R)引物,以第一輪2個重疊的PCR產(chǎn)物為模板進(jìn)行拼接。PCR反應(yīng)條件與上述相同,所有引物見表1。對兩個突變位點的PCR最終產(chǎn)物使用GFX PCR DNA and Gel Band Purification Kit(Amersham Pharmacia Biotech)從1%瓊脂糖凝膠中回收PCR產(chǎn)物中的DNA片段。純化的DNA片段和表達(dá)載體pET30a(Novagen,Madison,WI)分別用相應(yīng)的限制性內(nèi)切酶消化后進(jìn)行重組。所得質(zhì)粒用于轉(zhuǎn)化大腸桿菌BL21。重組載體利 用 BigDye( Applied Biosystems, Foster City,CA)進(jìn)行測序驗證。

表 1 引物序列Table 1 Primer sequences
1.3 重組PtGSTU1和突變體的表達(dá)和純化
將含有重組pET30a質(zhì)粒的大腸桿菌BL21過夜培養(yǎng),然后以1:100稀釋,震蕩培養(yǎng)直至光密度(A600)達(dá)到0.5;加入0.1 mmol·L-1IPTG誘導(dǎo)蛋白表達(dá),并分別在37、25、20℃下繼續(xù)培養(yǎng)8 h;4℃以6 500 g離心10 min收集菌體,重懸于結(jié)合緩沖液(20 mmol·L-1磷酸鈉,0.5 mol·L-1NaCl,20 mmol·L-1咪唑,pH值7.0)中,低溫超聲破碎;然后將勻漿在4℃下以10 000 g離心10 min。通過SDS-PAGE分析所得沉淀和上清液。
確定成功表達(dá)可溶蛋白后,剩余的上清液上樣到已用結(jié)合緩沖液預(yù)平衡的Ni Sepharose High Performance柱(Amersham Pharmacia Biotech)上。用洗脫緩沖液(20 mmol·L-1磷酸鈉,0.5 mol·L-1NaCl,500 mmol·L-1咪唑,pH 值 7.0)洗脫與 Ni Sepharose High Performance柱結(jié)合的目的蛋白質(zhì)。使用PD-10柱(Amersham Pharmacia Biotech)對蛋白溶液進(jìn)行脫鹽,脫鹽緩沖液為10 mmol·L-1Tris-HCl緩沖液(pH值7.0)。
1.4 酶學(xué)活性及熱穩(wěn)定性測定
根據(jù) Habig等[1]描述的測定方法,測定GST對CDNB(1-chloro-2,4-dinitrobenzene)、ECA(ethacrynic acid)和4-NPA(4-nitrophenyl acetate)底物的催化活性。利用Ricci等[18]所述方法,測定對 NBD-Cl( 7-chloro-4-nitrobenzo-2-oxa-1,3-diazole)的催化活性。所有測定均在25℃下進(jìn)行。根據(jù)Lowry等[19]的方法計算蛋白質(zhì)濃度。使用梯度濃度(0.1 至 1 mmol·L-1)的 GSH(glutathione)和 1 mmol·L-1的CDNB測定對 GSH的米氏常數(shù)(Km)和酶被底物飽和時的反應(yīng)速度(Vmax)。使用梯度濃度(0.1至 1 mmol·L-1)的 CDNB和1 mmol·L-1的 GSH測定對 CDNB的米氏常數(shù)(Km)和酶被底物飽和時的反應(yīng)速度(Vmax)。將數(shù)據(jù)繪制為雙倒數(shù)Lineweaver-Burk圖以確定Km和Vmax值。
從25~65℃,以5℃間隔的溫度梯度溫育樣品15 min后測定對CDNB的底物活性。分別對野生型(Wild type)和突變體 D103A(0.45 mg·mL-1蛋白溶于 10 mmol·L-1Tris-HCl,pH 值 7.0)繪制熱穩(wěn)定性曲線。
2 結(jié)果與分析
2.1 PtGSTU1單體結(jié)構(gòu)分析
用PtGSTU1蛋白序列搜索蛋白質(zhì)結(jié)構(gòu)數(shù)據(jù)庫,獲得最優(yōu)模板為一個節(jié)節(jié)麥Tau類GST晶體(TaGSTU4,PDB id為1 gwc)。以TaGSTU4晶體結(jié)構(gòu)為模板進(jìn)行同源建模,獲得模擬的PtGSTU1蛋白三維結(jié)構(gòu)(圖1A)。結(jié)果發(fā)現(xiàn),PtGSTU1單體具有典型的GST三維結(jié)構(gòu),具有N端和C端兩個結(jié)構(gòu)域,其中,N端及C端結(jié)構(gòu)域形成的V字型區(qū)域表面主要為第一底物GSH的特異結(jié)合位點(G位點),C端活性口袋包含結(jié)合疏水底物的位點(H位點),G位點與H位點由一段短肽連接。
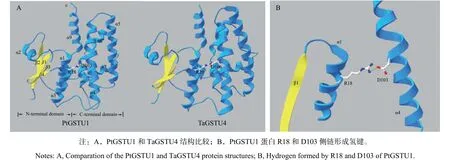
圖 1 PtGSTU1蛋白結(jié)構(gòu)Fig. 1 Structure of PtGSTU1
蛋白結(jié)構(gòu)模型顯示,N端結(jié)構(gòu)域一個精氨酸(R)位點能夠與C端結(jié)構(gòu)域一個天冬氨酸(D)形成氫鍵(圖1B)。將PtGSTU1與不同植物、不同類型GST蛋白進(jìn)行序列比對(圖2),發(fā)現(xiàn)PtGSTU1蛋白N端結(jié)構(gòu)域的βαβαββα結(jié)構(gòu)在Tau類GST中非常保守,而C末端的結(jié)構(gòu)可變性較大。第18位的精氨酸在所有GST中均保守,而第103位的天冬氨酸僅在Tau類GST中保守。

圖 2 植物GST蛋白序列比對Fig. 2 Alignment of plant GST proteins
2.2 PtGSTU1及其突變體蛋白的表達(dá)
為了探討PtGSTU1蛋白R18和D103位之間氫鍵對蛋白結(jié)構(gòu)穩(wěn)定性的影響,筆者對這2個位點分別單獨進(jìn)行了定點突變。將18位精氨酸分別突變?yōu)楫惲涟彼幔↖)、丙氨酸(A)、色氨酸(W)、賴氨酸(K)、谷氨酸(E)和組氨酸(H),構(gòu)建了R18I、R18A、R18W、R18K、R18E和R18H六個突變體蛋白,并將103位天冬氨酸突變?yōu)楸彼幔ˋ)構(gòu)建D103A突變體。這些用于替換的氨基酸殘基側(cè)鏈具有不同的極性和空間位阻。
對野生型PtGSTU1和所有突變體進(jìn)行蛋白的表達(dá)檢測(圖3),分別在37、25、20℃進(jìn)行蛋白的誘導(dǎo)表達(dá),對收集的菌體進(jìn)行破碎,破碎后分離細(xì)胞沉淀物和上清,進(jìn)行SDS-PAGE檢測。檢測結(jié)果顯示:野生型和突變體蛋白分子量在32 kDa左右,與預(yù)期一致。在37℃下,6個R18突變體均為包涵體,而野生型和D103A在上清中表達(dá)。在25℃和20℃下,R18A在上清檢測到微弱的表達(dá),其他5個R18突變體均為包涵體,野生型和D103A在上清中表達(dá)。
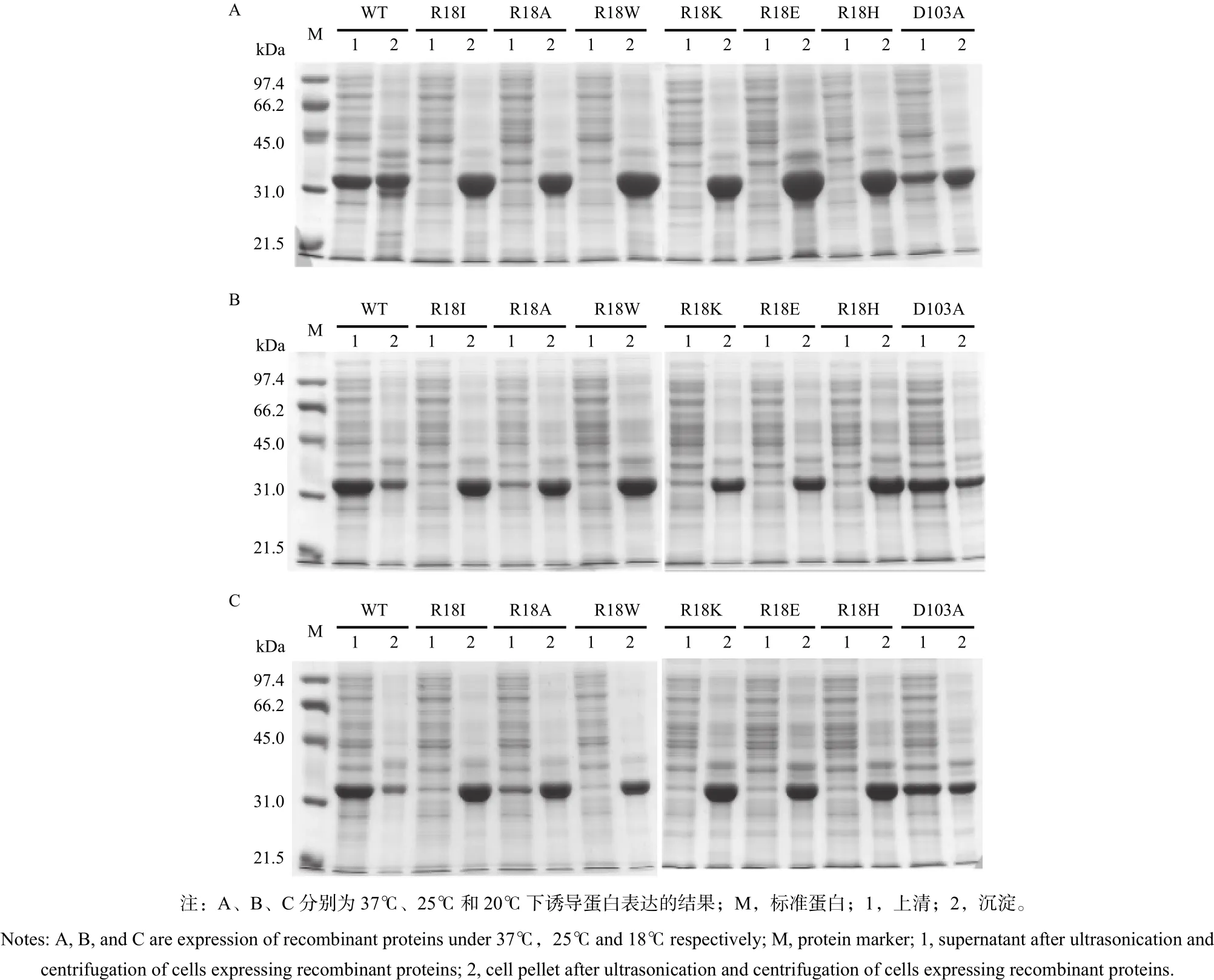
圖 3 PtGSTU1及其突變體蛋白表達(dá)Fig. 3 Expression of the wild type and mutants of PtGSTU1
在細(xì)胞沉淀中的蛋白多為由膜包裹的高密度、不溶性蛋白質(zhì)顆粒,因此,6個R18突變體無法獲得高純度的具有正確折疊的可溶蛋白。為了檢測D103突變體的酶學(xué)功能變化,筆者分別對野生型和D103A突變體蛋白進(jìn)行純化,利用Ni離子親和柱純化上清中表達(dá)的蛋白,以野生型蛋白為參照,純化后,重組蛋白均顯示單一條帶,在SDSPAGE上具有相同的遷移率(圖4)。
2.3 PtGSTU1及其突變體酶學(xué)功能測定
PtGSTU1蛋白R18位6個突變體的表達(dá)檢測顯示:僅R18A有微量蛋白能夠在上清中表達(dá),但由于表達(dá)量過低,純化后無法獲得高濃度可溶蛋白進(jìn)行酶學(xué)功能的檢測。為了檢測突變體是否還具有GST催化活性,筆者以含有pET30a載體的Bl21誘導(dǎo)表達(dá)后的上清液為陰性對照,測定R18A重組蛋白表達(dá)上清對CDNB的催化活性,結(jié)果顯示:R18A具有微弱的催化活性(OD340= 0.02)。
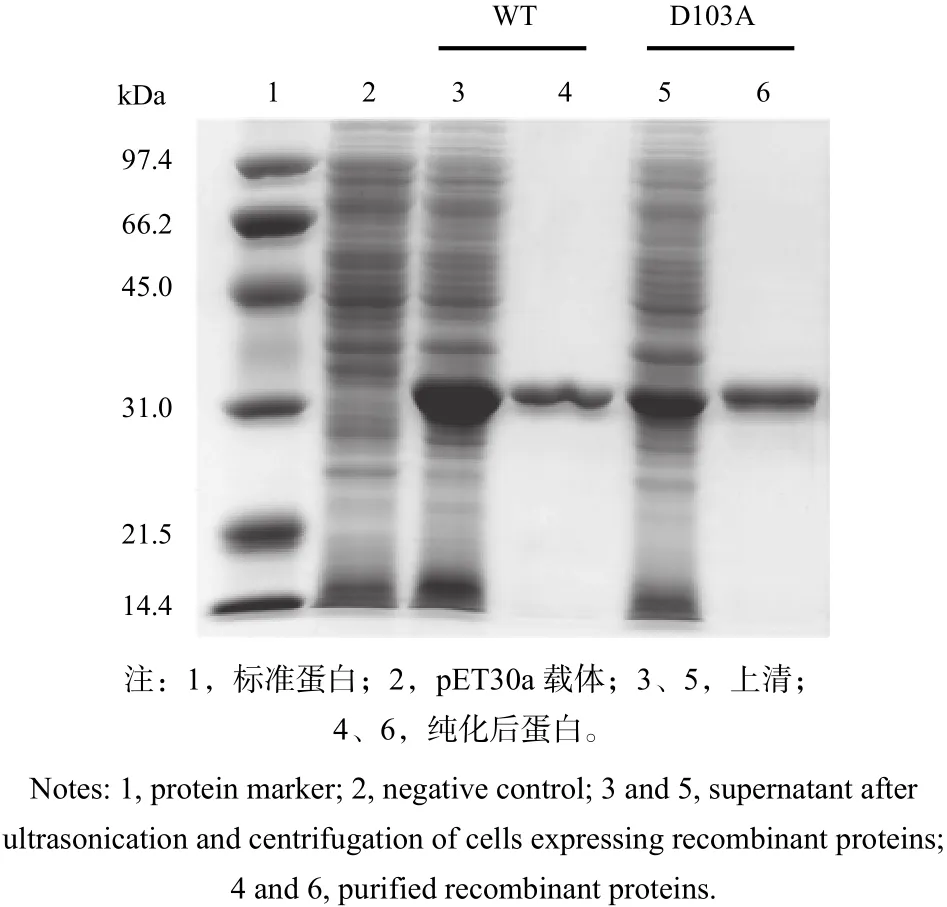
圖 4 PtGSTU1及D103A突變體蛋白純化Fig. 4 Expression of the wild type and mutant D103A of PtGSTU1
對純化后的D103A重組蛋白進(jìn)行酶學(xué)活性檢測發(fā)現(xiàn):103位天冬氨酸突變?yōu)楸彼岷螅鋵?種底物的活性明顯降低,對CDNB的催化活性僅為(1.70 ± 0.26) μmol·min-1·mg-1,與野生型相比降低了12.2倍;對NBD-Cl的催化活性為(0.45 ±0.08) μmol·min-1·mg-1, 與 野 生 型 相 比 降 低 了10.7倍,ECA降低1.1倍,對4-NPA幾乎檢測不到催化活性(表2)。
CDNB是GST經(jīng)典底物,廣泛用于GST活性的檢測,且PtGSTU1-D103A對CDNB的催化活性最高。我們使用CDNB和GSH作為底物測定PtGSTU1-D103A的動力學(xué)常數(shù)。發(fā)現(xiàn)PtGSTU1-D103A對兩個底物的米氏常數(shù)(Km)均有明顯增加,分別為 1.06 ± 0.25 mmol·L-1和 0.87± 0.17 mmol·L-1。這意味著與野生型相比,PtGSTU1-D103A對底物的親和力明顯降低。底物飽和時,PtGSTU1-D103A催化GSH和CDNB的反應(yīng)速率(Vmax)降低了至少9倍。相應(yīng)的,對底物的催化效率(kcat/Km)也明顯降低(表3)。
2.4 PtGSTU1及其突變體熱力學(xué)穩(wěn)定性分析
為了進(jìn)一步驗證R18和D103這2個氨基酸位點間氫鍵對Tau類GST蛋白單體結(jié)構(gòu)的影響,筆者分別對野生型和PtGSTU1-D103A蛋白的熱力學(xué)穩(wěn)定性進(jìn)行檢測。野生型蛋白在25~50℃之間保留了90%以上的催化活性,這表明其在50℃以下是穩(wěn)定的;而PtGSTU1-D103A突變體蛋白隨溫度升高,催化活性快速降低,在30℃時僅剩約40%的催化活性,預(yù)示著其蛋白熱穩(wěn)定性較低(圖5)。
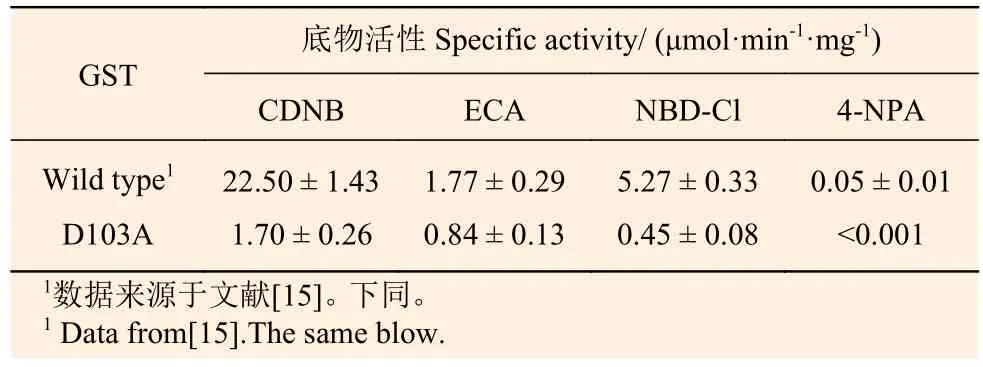
表 2 PtGSTU1野生型和D103A突變體蛋白底物活性Table 2 Specific activities of the wild type and D103A mutant of PtGSTU1 towards different substrates. The values shown are means ± S.D., calculated from three replicates
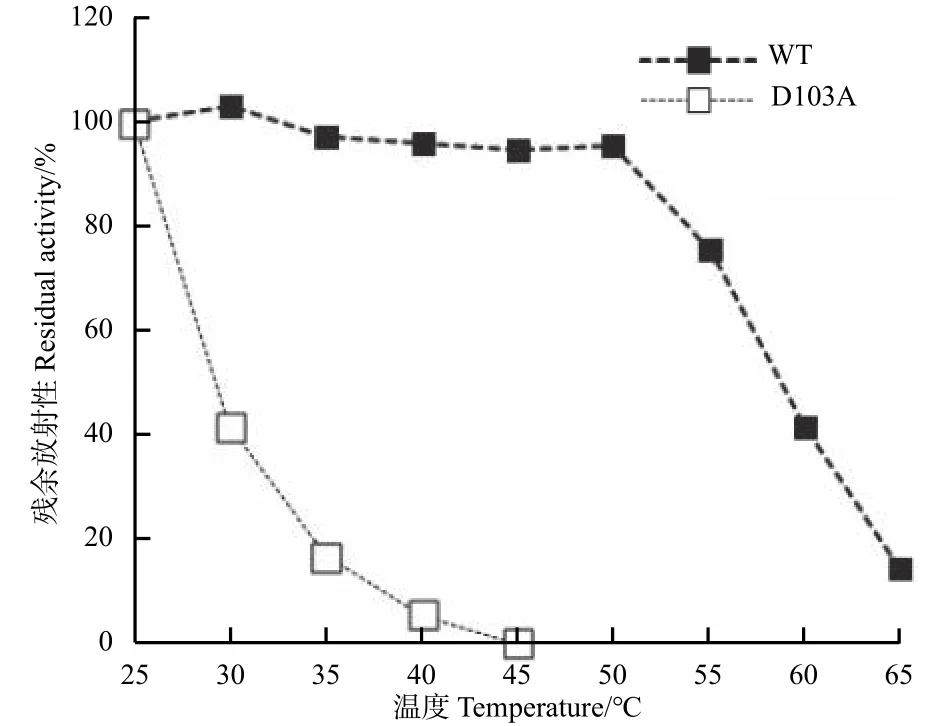
圖 5 PtGSTU1及D103A突變體蛋白的熱力學(xué)穩(wěn)定性Fig. 5 Thermal stability of the recombinant wild type and mutant D103A of PtGSTU1
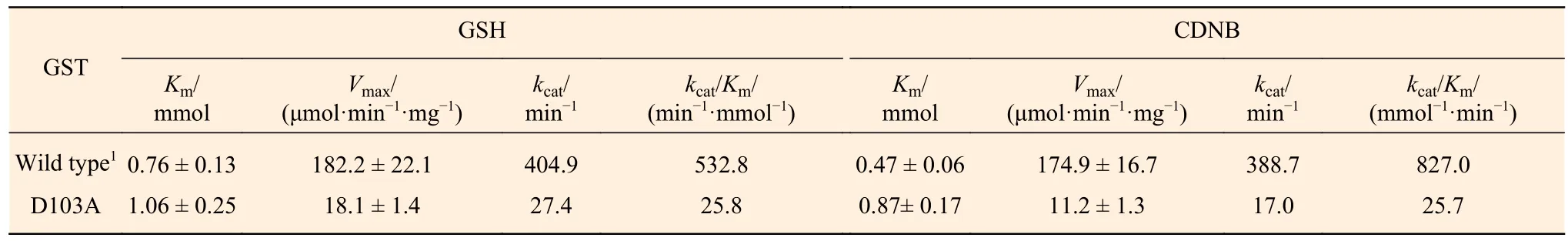
表 3 PtGSTU1野生型和D103A突變體蛋白動力學(xué)。Table 3 Steady-state kinetic constants of the wild type and D103A mutant of PtGSTU1. Data are means ± S.D, calculated from three replicates.
3 討論
基于與節(jié)節(jié)麥TaGSTU4蛋白晶體結(jié)構(gòu)的比較,筆者推測PtGSTU1的18位精氨酸和103位天冬氨酸能夠形成氫鍵,這個氫鍵穩(wěn)定了蛋白N端和C端的空間結(jié)構(gòu),使蛋白能夠形成正常的折疊。PtGSTU1的催化口袋位于N端和C端的空間結(jié)構(gòu)之間[16],第18位精氨酸位于α1螺旋上,這一位點在植物不同類型GST序列中均保守。PtGSTU1蛋白G-site活性位點13位的絲氨酸對GST蛋白活性至關(guān)重要[15-16],位于α1螺旋末端,α1螺旋空間結(jié)構(gòu)的改變將直接影響G位點與第一底物GSH的結(jié)合。
精氨酸為堿性氨基酸,其帶正電,具有-(CH2)3-NH-CNH-NH2的長側(cè)鏈,能在空間上最大限度接近位于C端α4螺旋上的103位天冬氨酸,與天冬氨酸側(cè)鏈上的-COOH形成N-H…O型氫鍵。將18位的精氨酸突變?yōu)橘嚢彼釙r(R18K),雖然其側(cè)鏈具有4個碳原子長鏈,能夠接近C端結(jié)構(gòu)域,但由于極性降低(R,pKa=12.0;K,pKa=10.5),影響了氫鍵形成。R18H突變體的18位精氨酸側(cè)鏈被組氨酸咪唑基取代,不僅極性降低(H,pKa=6.08),且側(cè)鏈長度變短。此外,當(dāng)突變?yōu)楫惲涟彼幔↖)和丙氨酸(A)時,側(cè)鏈上的氨基被甲基取代,同時影響了該位點的電負(fù)性和空間結(jié)構(gòu)。突變?yōu)楣劝彼幔‥)時,側(cè)鏈為-CH2-CH2-COOH,該位點由堿性氨基酸變?yōu)閹ж?fù)電的酸性氨基酸,破壞氫鍵形成。當(dāng)突變?yōu)樯彼幔╓)時,β-吲哚基增加了-N-H與103位天冬氨酸形成氫鍵的位阻。對突變體蛋白的表達(dá)分析顯示,無論將18位精氨酸突變?yōu)閴A性還是酸性氨基酸,均會影響蛋白的正確折疊,因此,所有R18突變體蛋白均表達(dá)為包涵體,盡管R18A在上清中檢測到微弱表達(dá),其活性也遠(yuǎn)低于野生型。這預(yù)示著在植物GST中,18位精氨酸可能是穩(wěn)定單體結(jié)構(gòu)的關(guān)鍵氨基酸之一。
與在植物中極為保守的18位精氨酸相比,103位天冬氨酸僅在Tau類GST中保守。將天冬氨酸-CH2-COOH側(cè)鏈替換為-CH3時,雖然失去了強極性的氧原子,但依然能夠形成正確的蛋白折疊;但其底物活性相較于野生型明顯降低,對底物的親和力降低了將近1倍,催化效率降低近10倍,預(yù)示著這一位點的突變可能影響了活性位點的空間結(jié)構(gòu)。同時,熱力學(xué)穩(wěn)定性檢測也驗證了,D103A突變體的穩(wěn)定性明顯低于野生型。說明,盡管103位天冬氨酸能夠參與形成氫鍵,但它的突變并不會使蛋白完全失活,而是明顯降低其的催化能力和穩(wěn)定性。這不僅印證了GST蛋白C端結(jié)構(gòu)域的多變性,同時預(yù)示著C端結(jié)構(gòu)域中可能存在其他氨基酸位點能夠與18位精氨酸形成氫鍵,從而穩(wěn)定蛋白單體折疊結(jié)構(gòu)。
4 結(jié)論
本研究通過對油松PtGSTU1野生型和突變體的酶學(xué)功能檢測發(fā)現(xiàn), N 末端 18 位精氨酸(Arg18)和 C 末端 103 位天冬氨酸(Asp103)形成的氫鍵對穩(wěn)定蛋白單體結(jié)構(gòu)具有重要作用。由于植物中GST蛋白N端結(jié)構(gòu)域具有保守性,C端結(jié)構(gòu)域具有多變性,Arg18的突變對單體穩(wěn)定性的影響大于Asp103 ,表明C端結(jié)構(gòu)域中可能存在其他氨基酸位點能夠與18位精氨酸形成氫鍵,從而穩(wěn)定蛋白空間結(jié)構(gòu)。
