一株木蹄層孔菌SO3高產漆酶發酵工藝及部分酶學特性研究
2020-04-27 04:48:48謝玉清代金平楊新平古麗努爾艾合買提王志方王小武
天然產物研究與開發 2020年2期
謝玉清,陳 競,代金平,楊新平,古麗努爾·艾合買提,王志方,王小武,馮 蕾*
1新疆農業科學院微生物應用研究所;2新疆特殊環境微生物重點實驗室,烏魯木齊830091
漆酶最早于1883 年在漆樹的汁液中發現,后來研究表明漆酶廣泛存在于自然界中,如植物、真菌、細菌、動物及昆蟲中[1]。漆酶屬于藍色氧化酶家族,是一種含銅的多酚氧化酶,以分子氧作為最終電子受體,是重要的木質纖維降解酶之一,還能催化降解多種芳香族化合物,特別是酚類[2]。因而漆酶作為一種綠色生物催化劑在生物質能源、紙漿生物漂白、染料脫色、廢水處理、食品加工等領域具有廣闊的應用前景[3]。
真菌漆酶比細菌漆酶、植物漆酶等具有更好的熱穩定性、金屬離子耐受性及更高的底物催化氧化性,在工農業及環境領域的應用中得到了較高的關注[4]。木蹄層孔菌屬擔子菌綱的白腐菌目,是漆酶最主要的生產者之一[5]。真菌漆酶的生產模式包括固態發酵和液體發酵,工業生產基本以液體發酵為主,而高酶活發酵工藝的優化及漆酶穩定性的研究是工業化生產漆酶的關鍵[6]。本研究對新疆阿勒泰山區分離篩選到的一株木蹄層孔菌(Fomesfomentarius)SO3菌產漆酶的發酵條件及部分酶學特性進行研究,為真菌漆酶的規模化生產奠定基礎。
1 材料與方法
1.1 實驗材料與試劑
1.1.1 實驗菌種
木蹄層孔菌(Fomesfomentarius)SO3為本實驗室從采自新疆阿勒泰山的菌種中分離篩選獲得。
1.1.2 培養基
產酶基礎培養基MF[7-9]:麥麩25 g,葡萄糖10 g,酒石酸銨1.84 g,NaCl 1.0 g、KH2PO42 g,琥珀酸鈉1.18 g, VB110 mg,聚山梨酯-80 0.5 g,微量元素溶液70 mL,加水定容至1 000 mL,pH調至5.5。
微量元素組成:MgSO4·7H2O 3.0 g,MnSO4·H2O0.5 g,ZnSO4·7H2O 0.1 g,CuSO4·7H2O 0.1 g,CaCl2·2H2O 0.1 g,KAl(SO4)2·12H2O 10 mg,H3BO310 mg,NaMnO4·2H2O 10 mg,加水定容至1 000 mL。
1.1.3 酶檢測試劑
0.1 mol/L NaAc-HAc緩沖液(pH 4.0),0.5 mmol/L 2,2-聯氮-二(3-乙基-苯并噻唑-6-磺酸)二銨鹽(ABTS),漆酶(laccase)。
1.2 實驗方法
1.2.1 漆酶活力測定
采用ABTS[2,2-azino-bis(3-ethylbenzthiazoline-6-sulfonic acid)]法[10-12]。反應體系組成為:0.1 mol/L NaAc-HAc緩沖液(pH4.0)1.95 mL、0.5 mmol/L 連氮-二(3-乙基苯并噻唑-6-磺酸)(ABTS)2.00 mL和適當稀釋的酶液50 μL, 28 ℃啟動反應。并在3 min內連續測定反應液420 nm(ε=3.6×104mol/L/cm)處吸光值的增加值。該條件下,每分鐘使1 μmol/L的ABTS氧化所需的酶量定義為1個活力單位(U)。
漆酶活力計算公式:漆酶活力(U/L)=(n×△A×106×V1)/(3.6×104×3×V2)
其中:n為酶液稀釋倍數;V1為反應總體積;V2為反應酶液體積;△A為3 min內反應液在420 nm處吸光度的變化值;3.6×104為ABTS氧化態的摩爾吸光系數(mol/L/cm)。
1.2.2 產漆酶發酵條件的研究
1.2.2.1 不同碳源對發酵產漆酶的影響
采用單因素試驗設計[13],在基礎培養基MF的基礎上進行碳源替換,分別以0.5%的麥芽糖、蔗糖、甘油、葡萄糖、羧甲基纖維素鈉、淀粉、麥麩作為碳源,裝液量50 mL/瓶,種子菌齡為5天,接種量5%,pH調至5.5,發酵7天后檢測漆酶活性。
1.2.2.2 不同氮源對發酵產漆酶的影響
采用單因素試驗設計,在基礎培養基MF的基礎上進行氮源替換,分別以0.5%干酪素、酵母粉、蛋白胨、硝酸銨、硫酸銨、磷酸二氫銨、酒石酸銨作為氮源,裝液量50 mL/瓶,種子菌齡為5天,接種量5%,pH調至5.5,發酵7天后檢測漆酶活性。
1.2.2.3 不同金屬離子添加對發酵產酶的影響
以MF為產漆酶基礎培養基,分別添加濃度為0、0.1、0.5、1.0、2.0 mmol/L的Cu2+、Mg2+、Fe2+、Mn2+、Zn2+、Ca2+、Co2+,培養6天,測定漆酶的活性(U/L)。
1.2.2.4 不同誘導劑對漆酶活性的影響[14]
以MF為基礎產漆酶培養基,pH調至5.5,裝液量50 mL/瓶,種子菌齡為5天,接種量5%,接種后第二天添加不同誘導劑,誘導劑分別為藜蘆醇、ABTS、沒食子酸、單寧酸及愈創木酚,終濃度為0.025 mmol/L。分別在第3、5、7、9、11天測定酶活。
1.2.2.5 正交實驗優化培養基
在MF基礎產酶培養基基礎上,各選擇兩種最優的碳、氮源進行組合,設計正交實驗,選用L9(34)正交實驗,具體組合如表1。

表1 培養基組成因素水平表
1.2.3 漆酶部分酶學特性研究
1.2.3.1 SO3漆酶的最適反應溫度和熱穩定性
在pH4.0的NaAc-HAc緩沖液中,在預試驗基礎上,分別在20、30、40、50、60、70、80 ℃條件下測定酶活,設定在50 ℃條件下測定的SO3漆酶活性為100%,確定漆酶最適反應溫度;在上述不同反應溫度下繼續保溫10 min后,再次測定漆酶活性,以反應起始的酶活為對照,計算殘留酶活,確定其熱穩定性。
1.2.3.2 SO3漆酶的最適反應pH
分別用pH為2.0、3.0、4.0、5.0、6.0、7.0、8.0的Na2HPO4-檸檬酸緩沖液配制反應底物,測定漆酶活力。
1.2.3.3 SO3漆酶pH穩定性
將酶液加入pH分別為3.0、4.0、5.0、6.0、7.0、8.0的Na2HPO4-檸檬酸緩沖液配制的底物,放入40 ℃的水浴中保溫24 h,測定酶活。
1.2.3.4 不同金屬離子對SO3漆酶穩定性的影響
用pH4.0的NaAc-HAc緩沖液配制FeCl2、MnSO4、NaCl、CoCl2、ZnSO4、CaCl2、MgSO4、CuSO4、KCl、CdCl2、CrCl3、PbCl2、AgNO3、BaSO4溶液,使其終濃度為4 mmol/L,在最適反應溫度下,以不添加上述金屬離子的反應體系作為對照,計算漆酶的相對活力。
1.2.4 SO3漆酶蛋白性質研究
1.2.4.1 SO3分泌蛋白SDS-PAGE分析
按最優的發酵條件獲得漆酶發酵液,10 000 rpm、4 ℃、離心20 min,經過了30%~80%的硫酸銨分段鹽析、Macro-Prep DEAE弱陰離子交換層析和Bio-Gel P-60 凝膠過濾層析系列純化,純化產物進行SDS-PAGE電泳分析[15]。
1.2.4.2 SO3分泌蛋白雙向電泳、漆酶等電點測定及主要蛋白質譜分析[16]
采用Bio-Rad蛋白質雙向電泳系統測定木蹄層孔菌(F.fomentarius)SO3漆酶的等電點,將漆酶發酵液經過10 000 rpm、4 ℃、離心20 min后,分別收集上清液和沉淀,使用pH7.5、20 mmol/L的Tris·HCl緩沖液將沉淀沖洗4~5次,然后加入Tris·HCl緩沖液進行超聲波破碎(冷卻進行)后,10 000 rpm、4 ℃、離心10 min,取上清液進行透析過夜、濃縮至所需體積即可。上清液則可直接進行透析過夜、濃縮至所需體積。等電點聚焦使用了Bio-Rad的IPG預制膠條,pH為3.0~10,pH梯度為線性梯度,通過Bio-Rad PDQuest 2-D Analysis software軟件分析獲得酶蛋白等電點pI。第二向SDS-PAGE電泳對木蹄層孔菌(F.fomentarius)SO3發酵產酶高峰時的菌體及發酵液進行上樣檢測對比分析。并將表達量高的漆酶蛋白點切下進行質譜測序分析(委托北京軍事醫學科學院完成)。
2 結果與分析
2.1 產漆酶發酵條件的研究
2.1.1 不同碳源對SO3產漆酶的影響
基礎產酶培養基以麥麩加葡萄糖(CK)的碳源組合酶活相對較高,為10 966 U/L,麥麩產酶活性為1 326 U/L,麥芽糖產酶活為313 U/L;添加蔗糖、甘油、葡萄糖、羧甲基纖維素鈉、淀粉替代基礎培養基中的麥麩加葡萄糖均未檢測到酶活(圖1),由此確定基礎培養基MF中麥麩加葡萄糖的碳源組合為最佳。
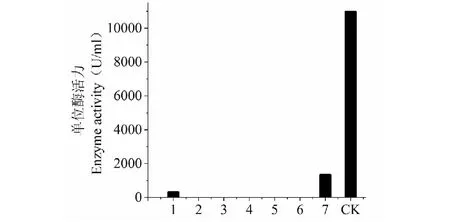
圖1 不同C源對SO3產漆酶的影響Fig.1 Effect of different carbon sources on the production of SO3 laccase注:1.麥芽糖;2.蔗糖;3.甘油;4.葡萄糖;5.羧甲基纖維素鈉;6.淀粉;7.麥麩;8.對照。Note:1.Maltose;2.Sucrose;3.Glycerin;4.Glucose;5.Carboxymethylcellulose sodium;6.Starch;7.Wheatbran;8.CK.
2.1.2 不同氮源對SO3產漆酶的影響
添加酵母粉替代原培養基中的酒石酸銨時,酶活最高,為7 566 U/L;其次為磷酸二氫銨,酶活為4 994 U/L;再次為蛋白胨,酶活為3 312 U/L;添加硝酸銨替代原培養基中的酒石酸銨時,酶活為1 224 U/L;添加硫酸銨和干酪素替代原培養基中的酒石酸銨時,酶活分別為1 155和1 080 U/L;而以酒石酸銨(CK)作為氮源時,酶活為1 113 U/L,由此確定,SO3產漆酶的最佳氮源為酵母粉,其次為(NH4)H2PO4(圖2)。
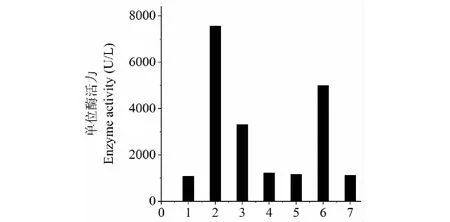
圖2 不同N源對SO3產漆酶的影響Fig.2 Effect of different nitrogen sources on the production of SO3 laccase注:1.干酪素(0.5%);2.酵母粉;3.蛋白胨;4.硝酸銨;5.硫酸銨;6.磷酸二氫銨(0.5%);7.酒石酸銨。Note:1.Casein(0.5%);2.Yeast extract powder;3.Pepton;4.Ammonium nitrate;5.Ammonium sulfate;6.Ammonium dihydrogen phosphate;7.Ammonium tartrate(CK).
2.1.3 不同金屬離子濃度對產漆酶的影響
以不添加金屬離子組(0 mmol/L)為對照(圖3), Mg2+、Mn2+、Zn2+、Ca2+的添加對SO3產漆酶有一定的促進作用,其中0.1 mmol/L的Mg2+、0.1 mmol/L和2 mmol/L Zn2+、0.2 mmol/L和0.5 mmol/L Ca2+的效果最明顯。添加0.1 mmol/L的Mg2+在第7 d所測得的酶活由對照組的761 U/L增加至7 983 U/L, 0.1 mmol/L Zn2+在第7 d所測得的酶活由4 809 U/L增加到 9 870 U/L,Zn2+濃度在2 mmol/L時酶活由4 809 U/L 增加到9 774 U/L;Fe2+和Co2+的添加,漆酶產量較低,對產酶影響不明顯;不同濃度Cu2+的添加都使酶活明顯下降,說明Cu2+對產酶具有抑制作用。
2.1.4 不同誘導劑對SO3產漆酶的影響
由圖4可知,添加沒食子酸和單寧酸可使產酶高峰期由對照的第7天提前到第5天;添加ABTS可使酶活由6 736 U/L提高至8 470 U/L;而藜蘆醇抑制產酶,愈創木酚對產酶無明顯影響。
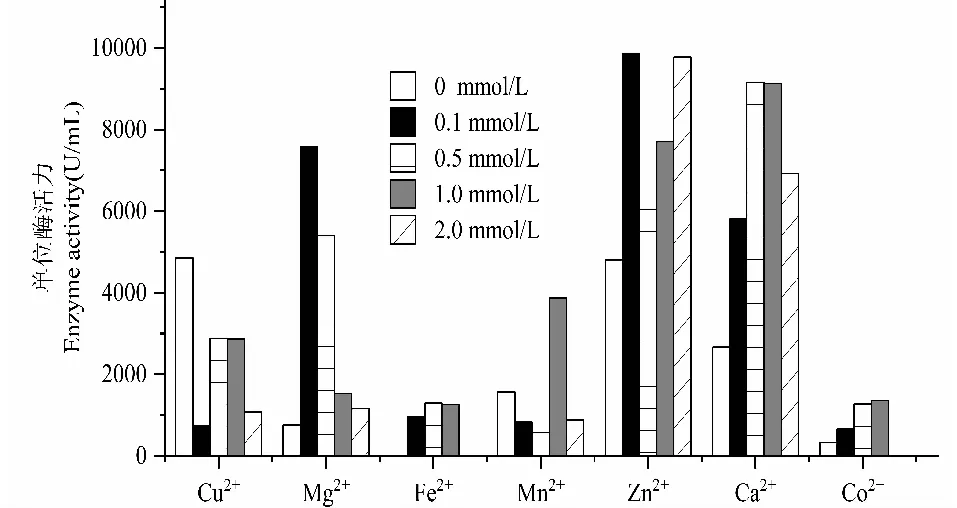
圖3 不同金屬離子添加對SO3產漆酶的影響Fig.3 Effect of different metallic ion
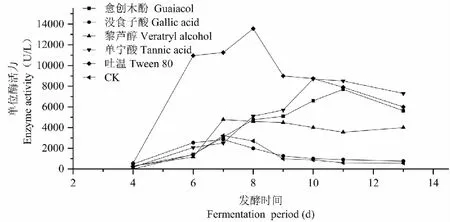
圖4 不同誘導劑對SO3產漆酶的影響Fig.4 Effect of different inducer
2.1.5 培養基的正交實驗結果
從表2和表3的正交實驗結果和方差分析可知,因素A(酵母粉)的第二個水平最佳,因素B(麥麩)的第三個水平最佳,因素C(葡萄糖)和D(磷酸二氫銨)最佳水平分別是第一和第二。因此最優培養基組分為A2B3C1D2,即酵母粉0.5%、麥麩2.5%、葡萄糖0.5%、磷酸二氫銨0.5%。極差(R)反映了不同因素對酶活影響的程度,其主次順序為A(酵母粉)>D(磷酸二氫銨)>B(麥麩)>C(葡萄糖)。通過此培養基進一步優化后,木蹄層孔菌(Fomesfomentarius)SO3漆酶活性達到10 863 U/L。
2.2 漆酶部分酶學特性研究
2.2.1 SO3菌漆酶的最適反應溫度
在預試驗基礎上,設定在50 ℃條件下測定的SO3漆酶活性為100%,20到80 ℃溫度區間內測定的反應液酶活分別為60.7%、79.6%、88.4%、100%、91%、98%、74%(圖5)。
2.2.2 SO3菌漆酶的熱穩定性
SO3漆酶反應液繼續保溫10 min后,對比相同溫度下起始酶活, 計算剩余酶活百分比,結果表明,40 ℃時穩定性最好(圖6)。
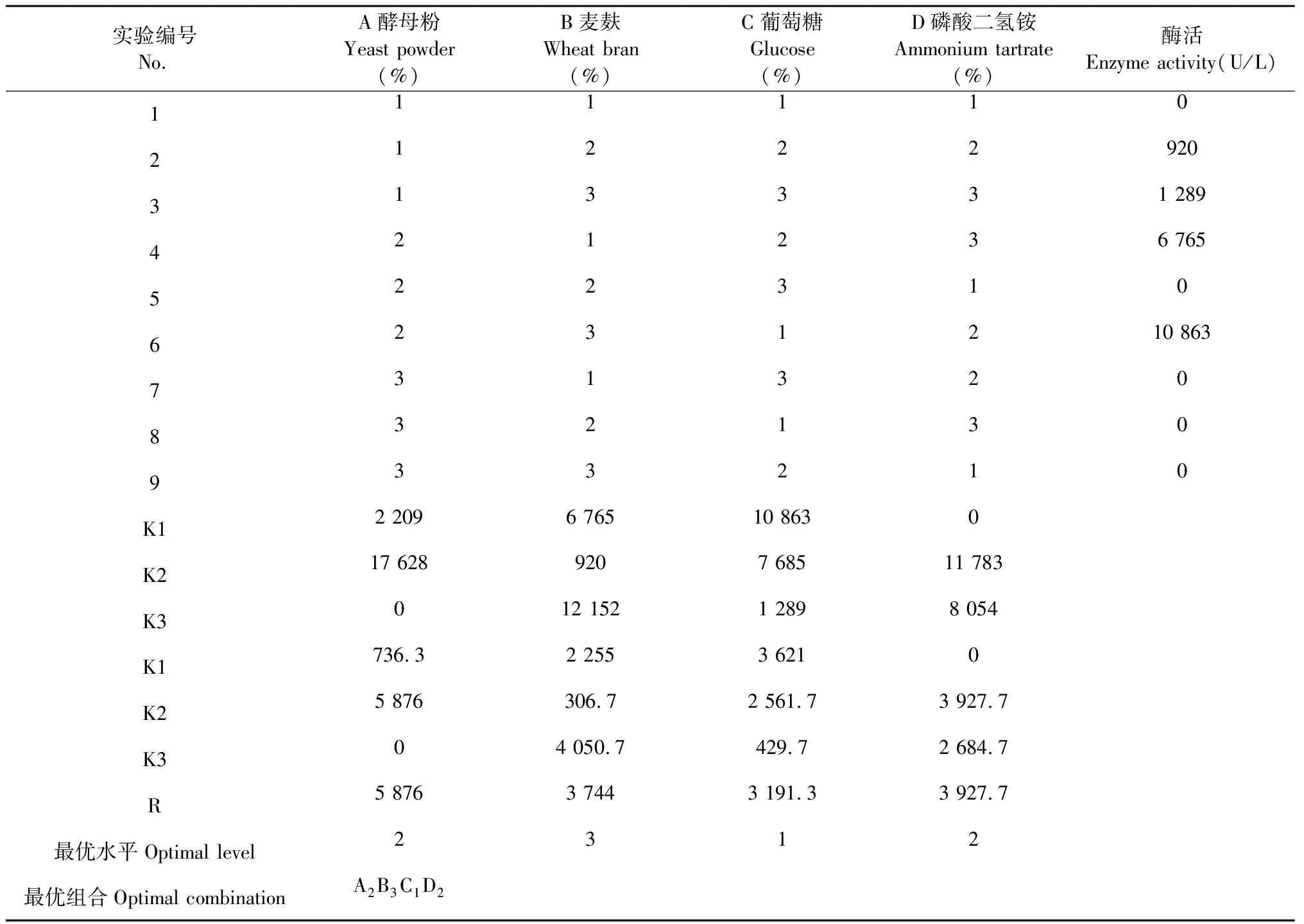
表2 木蹄層孔菌(Fomes fomentarius)SO3 L9(34)產酶條件正交實驗結果
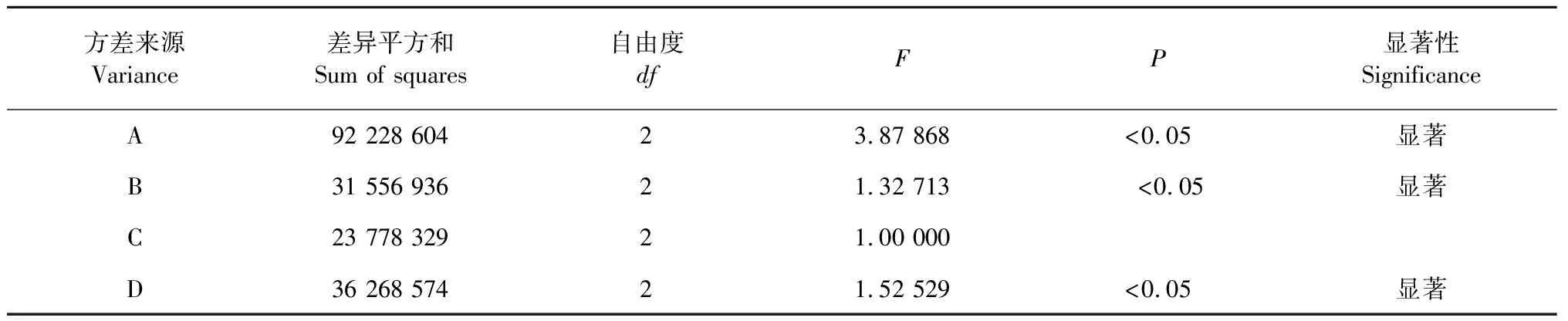
表3 方差分析表

圖5 SO3漆酶最適反應溫度Fig.5 The optimum reaction temperature of SO3
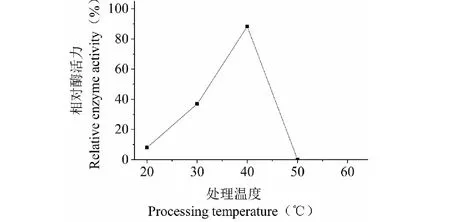
圖6 SO3漆酶熱穩定性Fig.6 Thermal stability of SO3
2.2.3 SO3菌漆酶的最適反應pH
實驗結果表明,pH為2.0到3.0時所測定的酶活相對較高,pH為2.0時酶活為1 295 U/L,pH4.0時酶活為897 U/L,pH為6.0以上時所測酶活為0(圖7)。
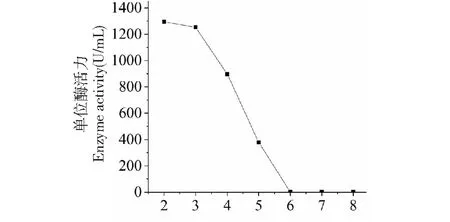
圖7 SO3漆酶最適反應pHFig.7 Effect of optimal pH on SO3 laccase
2.2.4 SO3菌漆酶pH穩定性
實驗結果表明,漆酶在40 ℃保溫24 h,pH4.0~5.0時酶活較穩定,pH3.0時相對酶活為42.1%,pH4.0時為73.6%,pH5.0時,酶活達到最大為97.8%,隨著pH值的升高酶活逐漸降低,超過pH6.0時殘留酶活為0(圖8)。
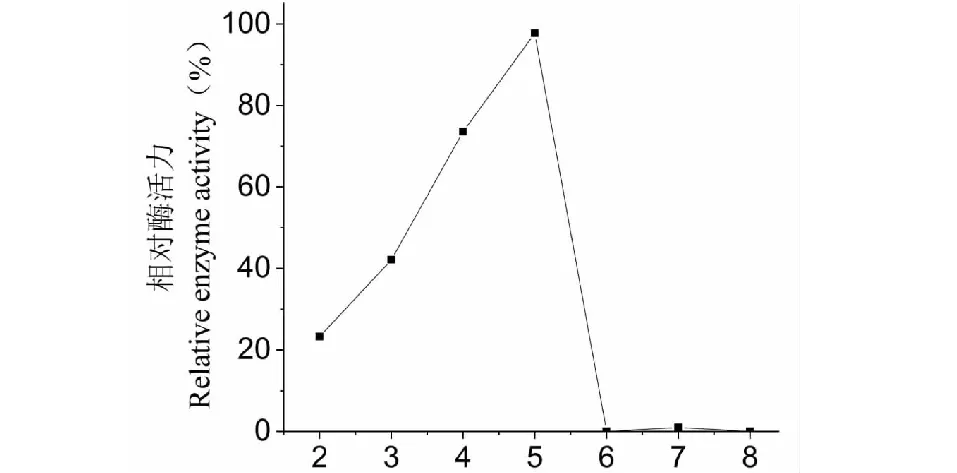
圖8 SO3漆酶的pH穩定性Fig.8 Effect of pH on SO3 laccase stability
2.2.5 不同金屬離子對漆酶穩定性的影響
結果表明,Mn2+和Zn2+對酶穩定性有一定的促進作用,其中在含有Zn2+的pH4.0的NaAc-HAc緩沖液反應體系中,所測酶活為955 U/L,為對照的102%,Mg2+、Cu2+、Ba2+酶穩定影響不大;Co2+、Cd2+、Cr2+和Pb2+對酶的穩定性具有明顯的破壞作用,其中加入Co2+時酶活為28%,Cd2+為47%,Pb2+為54%;而Fe2+對漆酶活性完全抑制(圖9)。
2.3 SO3漆酶蛋白性質研究
2.3.1 不同純化步驟的SDS-PAGE電泳
SDS-PAGE電泳結果如圖10所示,發酵培養基中含有多種蛋白及多肽,發酵液上清在61 KD左右有較明顯的蛋白帶,經過離子交換層析后基本呈單一條帶,經過凝膠過濾層析后,SDS-PAGE電泳顯示只有一條蛋白帶。通過一系列的蛋白分離純化實驗,獲得的漆酶已經達到了電泳純級。應用Bio-RAD quantity one分析軟件[17],根據標準蛋白相對分子量,得出木蹄層孔菌漆酶相對分子量約為61.5 KD。
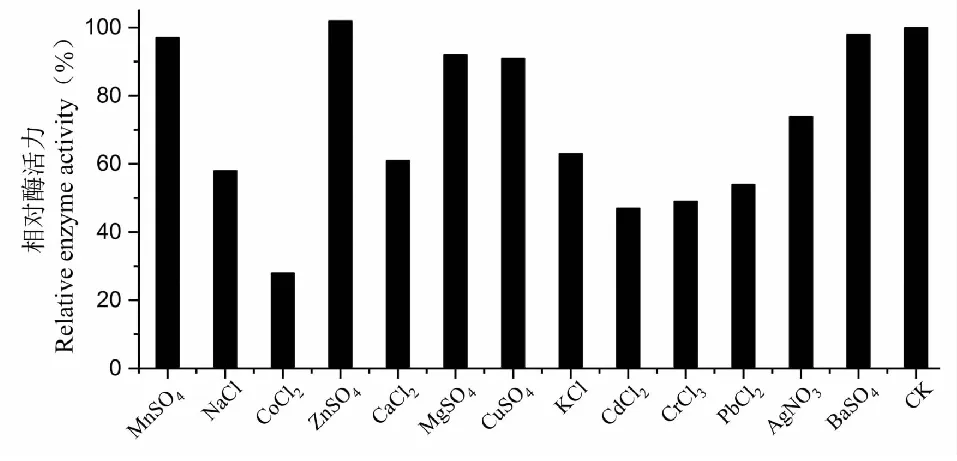
圖9 不同金屬離子對SO3漆酶穩定性的影響Fig.9 Effect of different metal ion on SO3
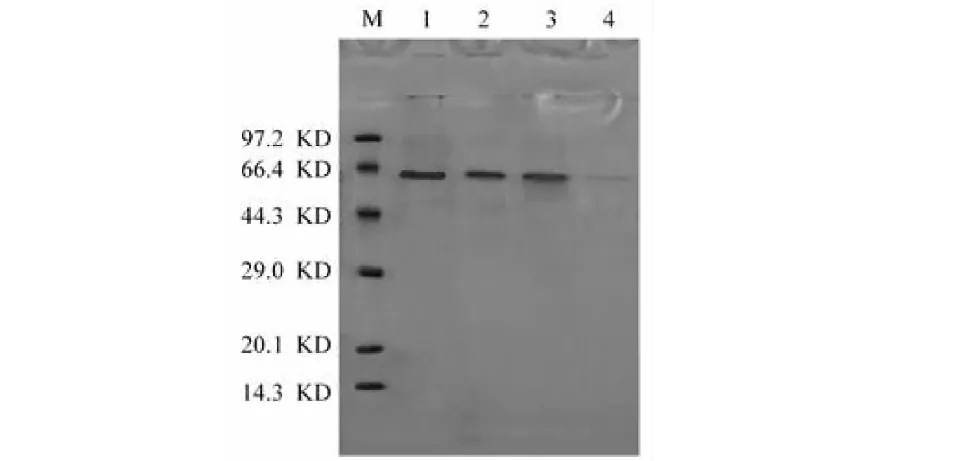
圖10 菌株不同純化步驟的SDS-PAGE圖Fig.10 SDS-PAGE of purification steps of F.fomentarius 注:M.蛋白相對分子量標樣;1.硫酸銨鹽析(60%);2.離子交換層析;3.硫酸銨鹽析(80%);4.凝膠過濾層析。Note:M.protein markers;1.Ammonium sulfate salting-out(60%);2.Ion-exchang chromatography;3.Ammonium sulfate salting-out(80%);4.Gel filtration chromatography.
2.3.2 木蹄層孔菌(F.fomentarius)SO3漆酶蛋白質雙向電泳及等電點
分別將以木蹄層孔菌(F.fomentarius)SO3發酵產酶高峰時的菌體及發酵液蛋白樣品用pH為3.0-10范圍的IPG非線性膠條進行等點聚焦,然后用12%的SDS-PAGE凝膠進行第二向分離。結果表明,菌體蛋白檢測出較多蛋白質點,而發酵液蛋白點則主要在同一等電點區域,通過Bio-Rad PDQuest 2-D Analysis software軟件分析,酶蛋白pI為4.1,將菌體及發酵液表達量高的同一蛋白質點切下進行質譜分析。
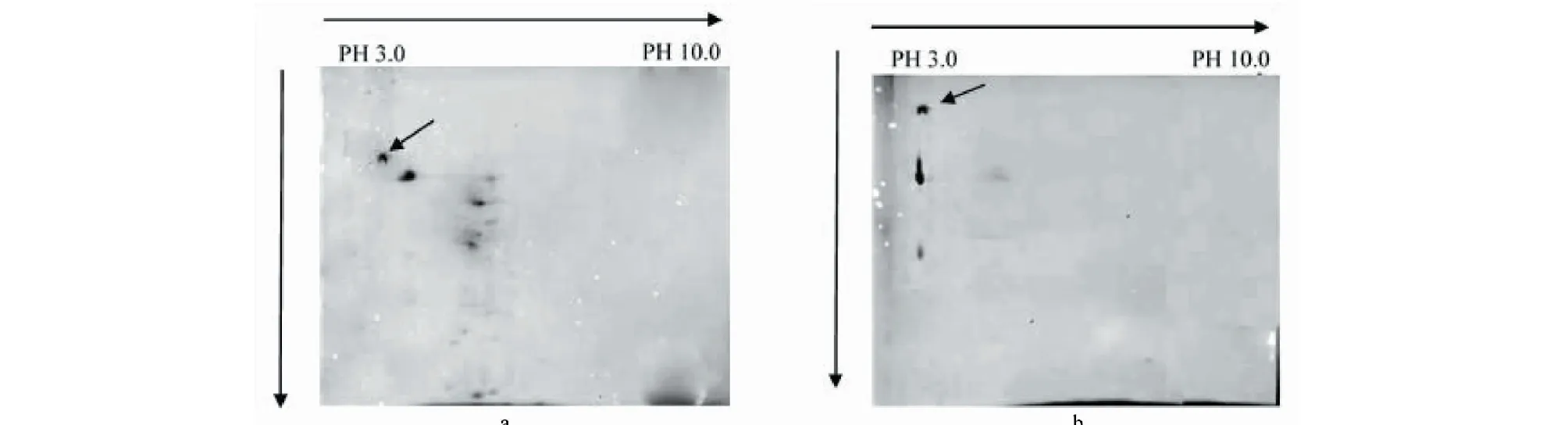
圖11 木蹄層孔菌(F.fomentarius)SO3菌體及發酵液的蛋白質雙向電泳Fig.11 Protein two-dimensional electrophoresis注:a.菌體;b.發酵液。Note:a.Ultrasonic disintegration sample cell;b.Upernatant of fermentation liquor.
2.3.3 木蹄層孔菌(F.fomentarius)SO3漆酶質譜分析
對主要表達蛋白進行MALDI TOF/TOF質譜分析(圖11),利用GPS Explor軟件進行分析,MASCOT檢索NCBI Nr蛋白質數據庫,成功鑒定1個蛋白點(蛋白質得分>50,可信度>90)(表4),鑒定的蛋白為LaccaseE(Trametessp.420),其肽脂紋圖譜中有8條序列均與LaccaseE蛋白匹配。
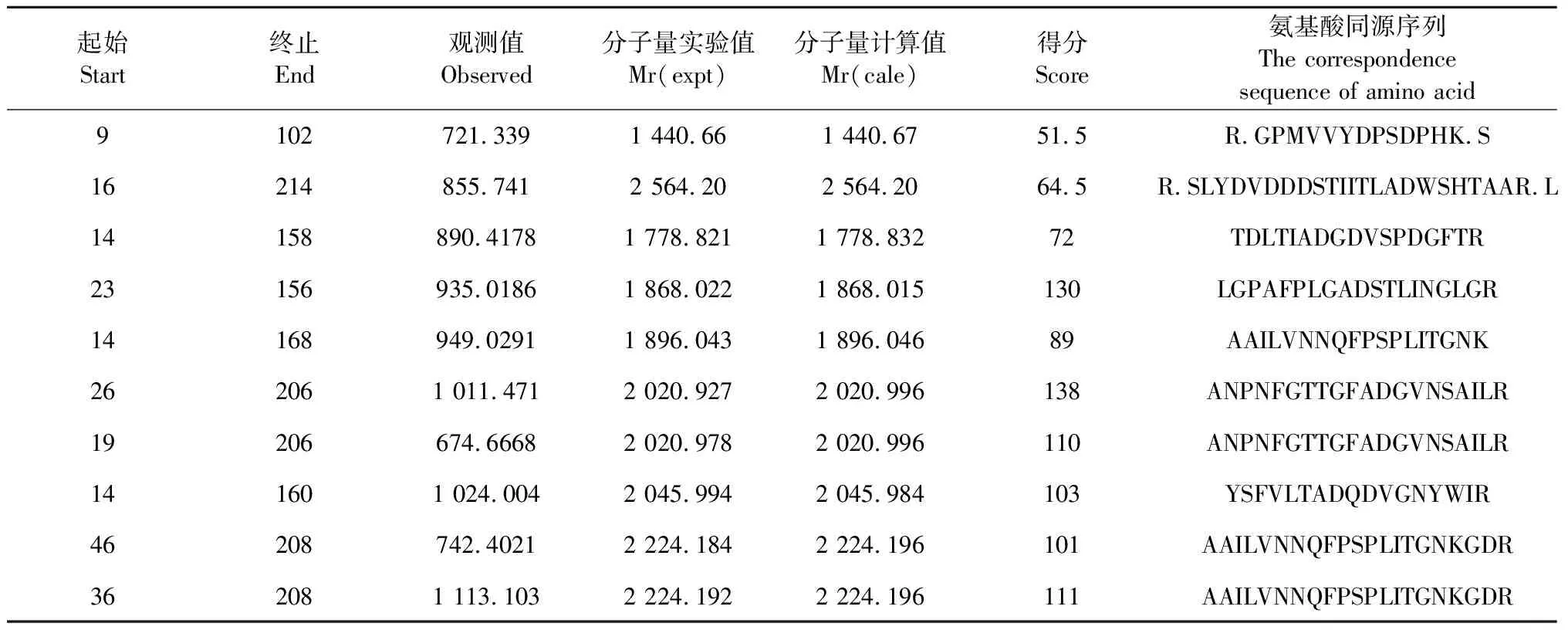
表4 木蹄層孔菌(F.fomentarius)SO3漆酶質譜分析中基本信息和匹配的肽段序列
3 討論與結論
漆酶是一種分布廣泛的多酚氧化酶,在食品、能源和環保等領域具有重要的應用價值。木蹄層孔菌可以合成分泌包括漆酶在內的多種木質纖維素降解酶,這些酶的活性會隨著發酵工藝的不同而不同,所以對產酶發酵工藝進行優化,對于提高漆酶產量和酶活具有重要意義。漆酶的合成和分泌受到營養水平、培養條件、生長階段以及培養基中誘導劑的嚴格調控,木質素或木質素相關的芳香類化合物、N源和C源也能調節漆酶的合成[18]。
微生物酶類分為組成酶和誘導酶。組成酶是指微生物無論在任何培養基中,總是適量地存在的一些酶類;誘導酶是依賴于酶作用底物或底物結構類似物的存在而合成的酶類[19]。漆酶胞外組成酶產量較低,當添加與木質素或木質素衍生物相關的芳烴類和酚類誘導物時,能顯著提高漆酶酶活。Hao[20]研究發現,ABTS、甲苯胺和對苯二酚對新疆野生巴爾喀什蘑菇產漆酶有明顯的促進作用,而咖啡酸、沒食子酸、鄰苯二酚、2,6-二甲氧基釀和愈創木酸對產酶誘導不明顯。Li等[21]利用Cu2+和麥麩作為共同誘導劑使白腐菌產漆酶活性達到51004 U/L。木蹄層孔菌(F.fomentarius)SO3漆酶對不同種類、不同濃度的金屬離子以及誘導劑的響應不盡相同,表明其具有較復雜的生理功能及調控機制,結果可為進一步闡明漆酶作用機制提供基礎。
培養基是發酵工藝優化中的重要因素,本試驗在基礎產酶培養基的基礎上,對碳氮源進行了進一步的篩選,并利用正交試驗對篩選出的碳氮源進行組合,最終確定0.5%酵母粉、2.5%麥麩、0.5%葡萄糖、0.5%磷酸二氫銨的最優組合,此時漆酶活性可達10 863 U/L。Chen等[22]對產漆酶黃孢原毛平革菌的培養基進行篩選優化,最終確定乳糖和酵母粉為最佳碳、氮源,使該菌的產酶能力提高了4.7倍。 Liu等[23]采用20 g/L 蔗糖、2 g/L酵母膏、3.2 g/L K2HPO4、0.2 g/L MgSO4·7H2O、3 mg/L SDS、6 mmol/L Cu2+、pH7.0的優化培養基,使有柄樹舌靈芝菌產漆酶活性達到496.18 U/mL,是優化前的12.2倍。
雙向電泳的主要分泌蛋白質譜分析結果表明,木蹄層孔菌(F.fomentarius)SO3漆酶胞外分泌蛋白類型主要為LaccaseE(Trametessp.420),獲得了該蛋白質的10個漆酶氨基酸序列片段,為后續差異基因表達序列的獲得提供了依據。