長鏈多不飽和脂肪酸結構脂合成方法及影響因素研究進展
2020-05-11 01:53:40王強賀稚非謝躍杰魏華恒李洪軍
食品與發酵工業 2020年8期
關鍵詞:結構
王強,賀稚非,謝躍杰,魏華恒,李洪軍*
1(西南大學 食品科學學院,重慶, 400715)2(重慶第二師范學院 生物與化學工程學院,重慶, 400067)3(重慶第二師范學院,脂質資源與兒童日化品協同創新中心,重慶,400067)
結構脂質(structured lipids,SLs),簡稱結構脂,是指經化學或酶法改變甘油三酯碳鏈骨架上脂肪酸組成或者位置分布,且具有特定分子結構、特殊功能作用的一類甘油三酯(triglyceride,TAG)。結構脂質在其結構上的差異不僅包含接入甘油三酯骨架上脂肪酸的不同種類,也包含由脂肪酸在甘油骨架上的隨機/選擇性定位效應(外側的sn-1和sn-3位,或中間的sn-2位)。由于甘油骨架特定位置上連接了具有特殊營養或生理功能的脂肪酸,所以結構脂質在保留天然油脂的部分或全部性質外,還能夠最大程度上發揮各種脂肪酸的功能[1]。有多項研究表明[2],結構脂作為一類新型油脂在體內消化吸收快,能量供應少,兼具有特定的保健營養和生物學功能,已逐步應用于食品、保健品、醫藥和化妝品等眾多行業。美國、日本先后開發出在減肥及脂肪替代等領域商用的結構脂Salatrim(美國納貝斯克)、Caprenin(美國寶潔)、Salatrim (美國Nabisco)、Olestra(美國寶潔)、diacylglycerol(日本花王)、NovaLipidTM以及Loders Croklaan CrokvitolTM系列產品。在所有結構脂中,長鏈多不飽和脂肪酸MLM型結構脂被認為是結構脂最理想的結構形式,具有吸收好、功能性強、營養特性顯著的優勢和特點。本文針對長鏈多不飽和脂肪酸結構酯的合成方法及產物合成影響因素的最新研究現狀進行綜述,以期為相關功能性結構脂的制備及結構修飾研究提供參考。
1 結構脂的分類及應用
結構脂并不是甘油三酯經簡單摻合形成的混合物,而是將不同脂肪酸以交叉組合的方式與甘油碳鏈骨架酯化后形成結構多樣的甘油三酯。結構脂有兩種公認的分類形式,一種是基于脂肪酸碳鏈長度分類。由于脂肪酸可分為短鏈脂肪酸(S,少于6個碳原子)、中鏈脂肪酸(M,6~12個碳原子)和長鏈脂肪酸(L,多于12個碳原子),因此,基于甘油三酯中sn-1,sn-2及sn-3位置的差異和脂肪酸碳鏈的長度,結構脂可劃分為sn-1-短鏈-2-中鏈-3-短鏈型(SMS)、sn-1-短鏈-2-長鏈-3-短鏈型(SLS)、sn-1-長鏈-2-中鏈-3-長鏈型(LML)、sn-1-中鏈-2-長鏈-3-中鏈型(MLM)、sn-1-中鏈-2-中鏈-3-中鏈型(MMM)、sn-1-中鏈-2-長鏈-3-長鏈型(MLL)等種類多樣的結構類型。此外,結構脂還可以基于結構的對稱性進行分類:非對稱性結構脂和對稱性結構脂。其中,對稱性結構脂還包括單酸型對稱結構脂(2-MAG)和二酸型對稱結構脂(1,3-DAG),非對稱結構脂包括二酸性非對稱結構脂(1,2-DAG)和三酸性非對稱結構脂(MLM或MMM等)[3]。有證據表明[4],結構脂的脂肪酸組成及其在甘油碳鏈骨架上的位置均與天然油脂原料有所不同,這些不同使得結構脂在物理、化學性質上與天然油脂有較為明顯的差異,特別是在油脂結晶構型、界面配向性、氧化穩定性和低熱值等方面有較大的差異。此外,甘油三酯的3個酰基位還可以連接低聚糖類、聚酯類化合物、淀粉類、生物堿等多種化合物,從而發揮更多理化及生理功能。SUN等[5]用蓖麻油作為咖啡酰基受體,通過酶促酯交換成功制備了一種新型蓖麻油基咖啡酰結構脂,該結構脂質具有較強的生物抗氧化和紫外吸收能力,目前已用于多個食品和化妝品領域用作抗氧化劑和紫外線吸收劑。在營養和醫療領域,結構脂還廣泛用于生產模擬母乳的嬰兒配方奶粉、塑性脂肪、可可脂替代脂以及低熱量脂肪的生產與應用[6]。
2 長鏈多不飽和脂肪酸結構脂的合成方法
長鏈多不飽和脂肪酸結構脂可以通過物理調配、化學催化、酶催化等方法改性制得。在這些方法中,物理調配法雖然較為安全,但產物合成率較低,往往無法有效定量得到理想的結構脂。化學催化法雖然能夠通過特定的反應合成路線獲得目標結構脂,但催化反應是隨機的酯交換反應,無特異選擇性,且反應產物難控制,不易生產出具有特定結構的重構脂質。此外,化學法反應條件劇烈(>100 ℃)、副反應多,產物得率低且分離困難,生產過程中的化學試劑也容易污染環境[7]。與化學催化法相比,酶催化法是一種較為安全、綠色和可控的有效制備方法,具有反應時間短、條件溫和(<70 ℃)、選擇性強、高效可控、操作簡單、環境友好等優點,可以通過調控反應條件(如時間、溫度、底物摩爾比、酶量等)來增加產品的純度和產量,是結構脂制備的首選方法。SUBAJINY等[8]認為酶法合成結構脂不含反式脂肪酸的優勢也是其可以廣泛用于食品工業油脂合成的重要原因。目前長鏈多不飽和脂肪酸結構脂的合成方法主要有3種:酯交換法、酸解法和醇解法。
2.1 酶法制備長鏈多不飽和脂肪酸結構脂方法
對于天然油脂而言,脂肪酸在甘油酰基位上的分布是隨機的。目前學術界主要有3種分布學說,分別是:全隨機分布學說、sn-1-隨機-sn-2-隨機-sn-3-隨機分布學說、sn-1,3-隨機-sn-2-隨機分布學說。對于天然植物油的甘三酯來說,sn-1,3-隨機-sn-2-隨機分布學說具有更廣泛的普適性。與天然油脂的結構相比,脂肪酶可定向作用于甘油三酯的特殊酰基位置,因此酶法合成結構脂的產物主要由結構脂的制備方法決定。KIM等[9]用兩種固定化脂肪酶將二十碳戊烯酸(EPA,eicosatrmacnioc acid)與葵花籽油進行酯化,不僅降低了天然玉米油中三油酸甘油三酯的含量,改性后的甘油三酯還含有更高含量的EPA。PINA-RODRIGUEZ等[10]使用Novozym 435脂肪酶在莧菜油sn-2位置接入了棕櫚酸,然后用脂肪酶RM IM在sn-1,3位置接入二十二碳六烯酸(DHA,docosahexaenoic acid),最終將DHA作為目標脂肪酸加入至莧菜油甘油三酯中,改變了原有天然莧菜油的脂肪酸組成。
2.2 基于脂肪酶的長鏈多不飽和脂肪酸結構脂酯交換法
酯交換法是酰基脂肪酸在TAG內部和TAG之間隨機化或定向交換位置的過程。酶促酯交換反應分兩步完成,脂肪酶Ser—OH基團親核進攻酰基碳,形成酰基-酶共價復合物后導致脂肪酸和甘油骨架碳鏈之間的鍵發生斷裂,待釋放后的脂肪酸與游離脂肪酸混合后,再隨機或定向接入至甘油骨架“空白”的位置形成新的甘油三酯。TEICHERT等[11]通過Novozym 435脂肪酶催化三棕櫚酸甘油酯(PPP)與硬脂酸大豆油酯交換反應合成sn-2位棕櫚酸結構甘油三酯,在反應時間18 h,反應溫度65 ℃,底物摩爾比1∶2條件下得到了含量為60.63%的sn-2位棕櫚酸酯。NAGACHINTA等[12]則通過Lpozyme TL IM脂肪酶催化三棕櫚甘油酯與二十二碳六烯酸、花生四烯酸(AA)乙酯乙酸進行酯交換合成含有sn-2位富含棕櫚酸與不飽和脂肪酸的結構脂質,其sn-2位棕櫚酸含量可高達40%以上。在脂肪酶作用的酯交換反應中,當原料油脂與目標產物均為甘油三酯時,目標產物分離較為困難且無法通過分離副產物的方式使反應平衡向目標產物方向移動。因此,針對不同的原料脂肪酸差異化的選擇定向脂肪酶是降低副產物、提高最終產物合成率的關鍵因素之一(圖1)。
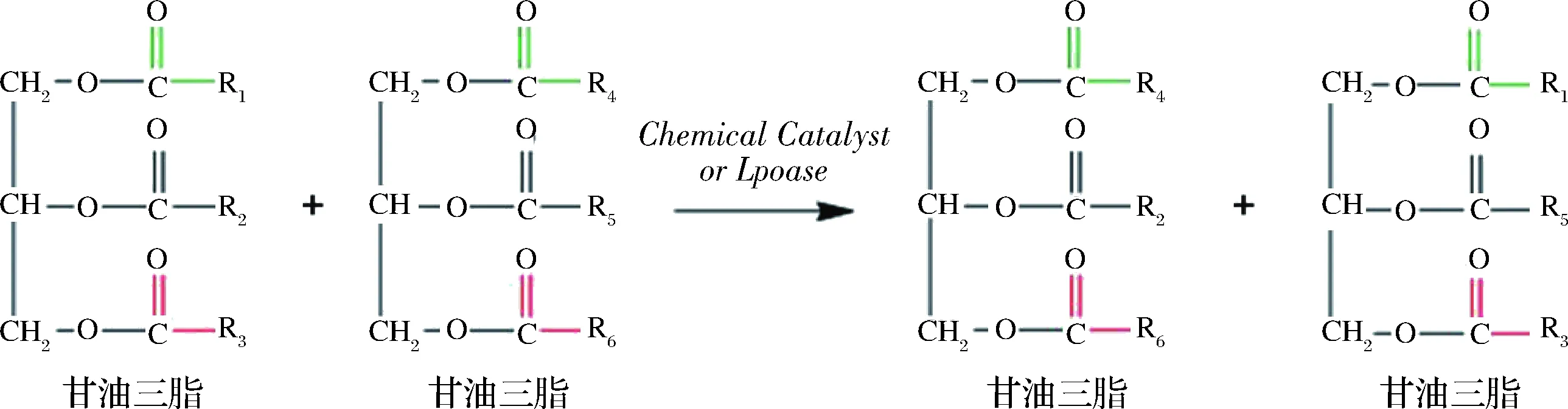
圖1 酯交換法制備結構脂反應示意圖Fig.1 Schematic diagram of structural lipid reaction prepared by transesterificatio
2.3 基于脂肪酶的長鏈多不飽和脂肪酸結構脂酸解法
與酯交換法類似,結構脂的脂肪酶酸解反應也需要兩步完成:水解與酯化。首先TAG在酶作用下水解生成游離脂肪酸和DAG(或MAG),在脂肪酶的作用下新的游離脂肪酸再與DAG(或MAG)酯化合成TAG。對于酸解反應而言,要制備sn-2長鏈多不飽和脂肪酸TAG,游離脂肪酸最好來源于單一長鏈多不飽和脂肪酸,在此條件下得到的目標TAG含量才會比較高。TEICHERT等[13]通過脂肪酶催化sn-2位含棕櫚酸的單硬脂酸大豆油結構脂與DHA和α-亞麻酸(ALA)酸解反應制成含PUFA的結構脂,其sn-2位棕櫚酸含量均能夠達到54%以上。酸解反應常被用于在非水介質中由sn-1,3-特異性脂肪酶催化合成sn-2長鏈多不飽和脂肪酸結構脂,其優勢在于能夠更簡單地預測反應產物的組成[14]。但是,在無溶劑系統中使用熱穩定性較好的脂肪酶可以有效降低最終產品中脂肪酶純化和回收,因此,基于脂肪酶的酸解反應通常需要增加中長鏈多不飽和脂肪酸的含量來提高結構脂的合成率。
在酸解法制備多不飽和脂肪酸TAG過程中,辛酸和癸酸常被用作sn-1,3位置的酰基供體積。YANG等[15]利用脂肪酶對金槍魚油和辛酸進行酸解,得到的MLM型結構脂類反應產物中sn-1,3位的辛酸占比為22.50%,sn-2位點DHA+EPA的含量則與生金槍魚油相似。該研究表明制備sn-2長鏈多不飽和脂肪酸TAG的供體最好以富含長鏈多不飽和脂肪酸的動植物油為供體進行酯基化反應為宜。除了酰基供體以外,在水解過程中影響酸解反應的主要因素還包括特異性脂肪酶和底物的摩爾比,適量増加脂肪酸和選擇適當的脂肪酶均可提高結構脂的產量。WEI等[16]在有溶劑條件下使用sn-1,sn-3特異性脂肪酶催化油酸與PPP生成sn-2位棕櫚酸甘油酯,并研究了酶添加量和底物摩爾比對酸解反應的影響,結果顯示,結構脂質在無溶劑和有溶劑情況下得到的產率分別為40.23%和32.34%,其中sn-2位棕櫚酸酯的含量分別高達86.62%、92.92%。由于酸解法中的甘油三酯可根據需要直接采用各種油脂,由于原料來源廣泛,所以生產成本相對較低,產物中過量的游離脂肪酸可采用蒸餾的方法從混合物中簡便的分離出去。因此,工業化生產選擇性結構脂質生產大多采用酸解法。

圖2 酶促酸解法制備結構脂反應示意圖Fig.2 Schematic diagram of enzymatic hydrolysis reaction
2.4 基于脂肪酶的長鏈多不飽和脂肪酸結構脂醇解法
酶催化醇解合成多不飽和脂肪酸TAG通常采用兩步法:首先是富含PUFA的油脂或三多不飽和脂肪酸甘油酯(如DHA甘油三酯)與乙醇在sn-1,3特異性脂肪酶作用下催化醇解反應生成sn-2單甘酯(2-MAG);其次是游離脂肪酸與2-MAG在脂肪酶的催化下進行酯化得到長鏈多不飽和脂肪酸TAG。SCHMID等[17]通過sn-1,3位特異脂肪酶催化醇解法制備sn-2位棕櫚酸甘油酯,第一步將PPP醇解得到MPG,使用低溫溶劑結晶法分離MPG,其純度超過95%且得率達85%;第二步在相同酶催化作用下使油酸與MPG進行酯化反應制得sn-2位棕櫚酸甘油酯,其含量高達96%。PFEFFER等[18]也采用相同的方法合成sn-2位棕櫚酸甘油酯,第一步得到的2-MPG純度為77%且產率為73%,第二步制得sn-2位棕櫚酸含量達95%。盡管采用醇解法制備的TAG副產物較少且易分離,目標產物sn-2不飽和脂肪酸TAG的純度和產量也較高,但是醇解反應分兩步進行,反應中間環節還需要分離單甘酯,因此酶催化醇解方法的成本較高,并不適用于工業化生產。
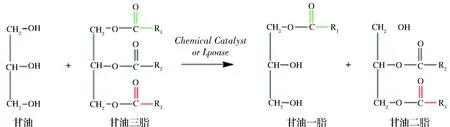
圖3 酶促醇解法制備結構脂反應示意圖Fig.3 Schematic diagram of enzymatic hydrolysis reaction
盡管如此,與一步催化反應相比,醇解法制備TAG的過程可大大減少酰基轉移的發生,在增強反應特異性的同時,還提高了工藝生產率,適合于制備結構復雜、生產成本高的結構脂。一般情況下,在結構脂質合成的過程中甘油酯的酰基容易發生轉移,這個過程會產生不必要的甘油酯致使目標產物純度下降。即使采用sn-1,3區域選擇性脂肪酶,也會發生酰基遷移現象生成TAG的副產物。因此,酰基遷移必須主要通過控制反應參數(如反應溫度、催化劑負載、含水量和溶劑類型)來加以控制。操麗麗等[19]以菜籽油和無水CH3CH2OH為原料,用Lipozyme TL IM固定化脂肪酶催化制備高純度的2-MAG來獲取中長碳鏈型(MLM)結構脂。在最優化工藝條件下,2-MAG的含量從38.82%上升到90.76%。結果證實醇解法減少了sn-2酰基遷移并提高了工藝生產率。ABED等[20]在無溶劑體系中,用Lipozyme RM IM脂肪酶催化醇解微生物油與辛酸,得到的結構脂質在sn-2位含有49.45%的花生四烯酸,在sn-1,3位含有29.7%辛酸。
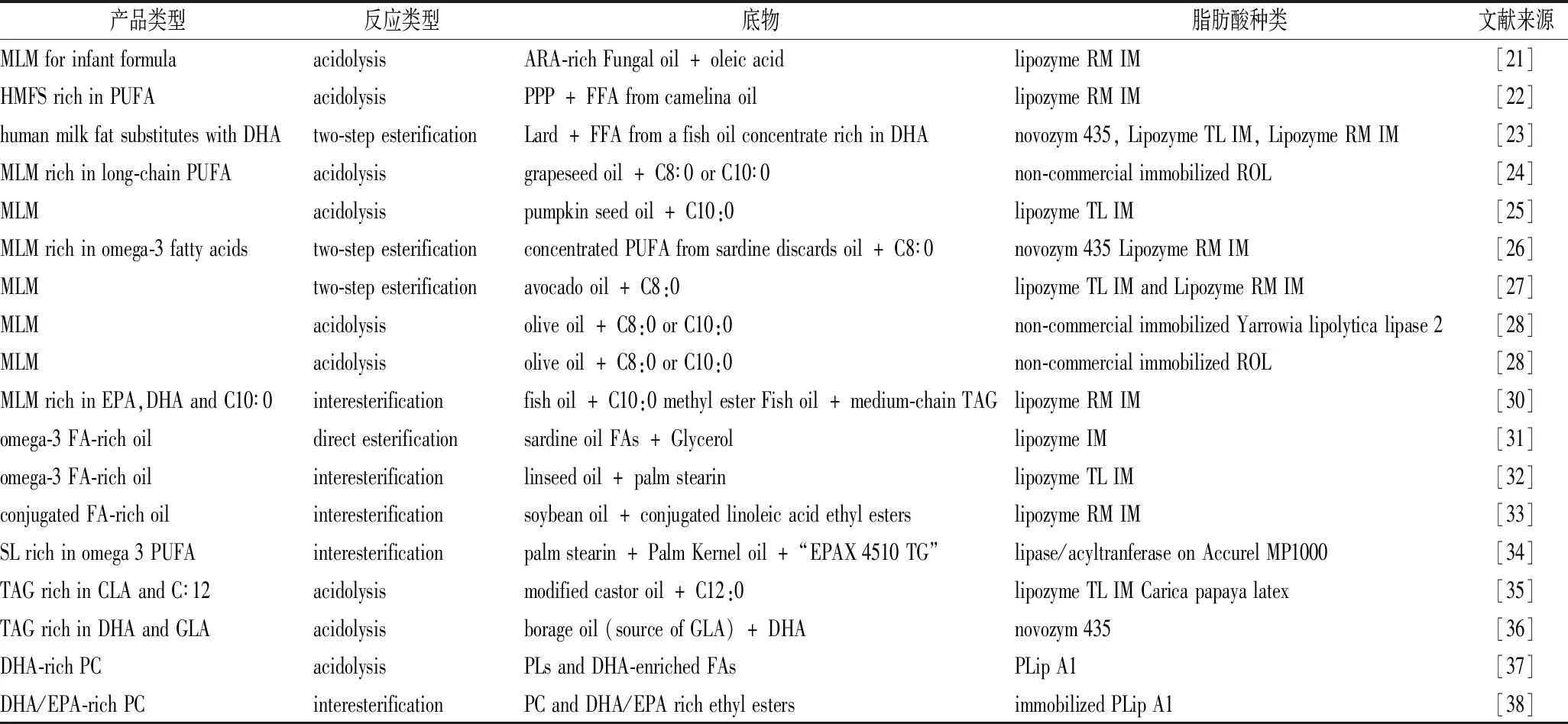
表1 長鏈多不飽和脂肪酸結構脂酶法合成研究底物及合成方法Table 1 Studies on the Lipase Synthesis of PUFAs for Structured lipids
2.5 基于脂肪酶的長鏈多不飽和脂肪酸結構脂直接酯化法
直接酯化法是以長碳鏈脂肪酸、中碳鏈脂肪酸(酰基供體)和甘油為原料,在合適的溫度下,控制好底物相應的摩爾比,在脂肪酶的催化作用下使反應平衡向有利于酯化的方向進行。SNEHAL等研究[39]在異辛烷中用直接酯化法探究了反應時間、脂肪酶的用量、底物摩爾比對合成膽固醇癸二酸單烯酯的影響,結果該實驗的平均酯化率與響應面擬合方程酯化率的預估值較吻合,表明直接酯化法工藝反應時間短,合成條件簡單,取得了理想的結果。姜洋等[40]在傳統的直接酯化法上,改變不同環境介質體系,將辛酸、癸酸和甘油等物料混合,用 Novozyme435 脂肪酶制備中碳鏈TAG,研究所得的中碳鏈TAG的產率為 95.1%,酯化率為 98.62%。結果表明,該制備工藝不僅有效提高了脂肪酸的利用率及中碳鏈TAG的產率,而且所得酯化產物色澤比較淺。由此可見,直接酯化法可一步完成,且反應時間較短,酶反應器利用率高,生成的副產物少,產物純度高,易分離純化出產品。但是,直接酯化法在反應過程中的酯化反應均為一次性完成,因此反應過程中需要及時脫水,水量的不斷增加會加厚酶分子表面的水膜,會阻礙底物與酶活性部位的結合,致使產率降低。因此,在sn-2位長鏈結構脂直接酯化法反應中要隨時除去水分,以防止逆向水解反應降低結構脂質的產率。
3 酶法合成長鏈多不飽和脂肪酸結構脂的影響因素
脂肪酶(lipase),又稱甘油三酯水解酶,是由以甘露糖為主的親水糖基部分和以蛋白質為主體的疏水部分組成,其活性中心靠近分子的蛋白質部分。脂肪酶的來源很多,商業用途脂肪酶主要來源于微生物、植物和動物源性。其中,微生物脂肪酶大多屬于依賴于由 Ser、His 和 Asp 殘基形成的催化三聯體的α/β脂解酶,熱穩定性好,催化過程中無需輔酶,應用范圍較廣。盡管脂肪酶用于酶解合成結構脂,并作為生物催化劑用于TAG的水解和酯化,但是在酶法合成結構脂反應中,并不是所有的脂肪酶都具有酰基位置選擇性,大多數脂肪酶水解TAG的酰基位置也具有隨機性,只有一部分特異性脂肪酶(如sn-1,3脂肪酶)在接近室溫和常壓條件下才會在甘油三酯的某個酰基位點發揮定點催化作用。
盡管來自微生物源的商業固定化脂肪酶已廣泛用于結構脂的制備,但因為其價格昂貴,限制了某些工藝和產品難以工業化生產[41]。相對于化學反應來說,酶法合成結構脂的優點是操作溫度較低,熱降解最小,且可得到理想的反應產物。盡管如此,受反應條件的制約,還是有諸多因素會影響酶法合成結構脂,這些因素包括:酶的種類、酰基供體種類、反應溫度、反應時間、水分活度、酶量及底物比例等[42]。在眾多影響因素中,酶的種類對結構脂合成類別尤為重要,是結構脂合成的主要限制性條件之一。因此,脂肪酶的活性、穩定性、重復利用次數、成本和可得到性等因素也決定著酶催化結構脂工業化生產的可行性。
3.1 長鏈多不飽和脂肪酸結構脂脂肪酶種類及特性
脂肪酶對結構脂位置及酰基種類的特異選擇性與脂肪酶來源、脂肪酶結合位點、界面處物理化學狀態以及底物結構特征有較大關系。根據反應特異性類型,長鏈結構脂脂肪酶大致可以分為三類:第一類脂肪酶催化時具有位置或區域選擇性,這類脂肪酶會定向專一水解TAG中sn-1和/或sn-3的酰基連接位置,使反應環境中較多的脂肪酸優先連接在TAG的sn-1和/或sn-3位置上。由于空間位阻及催化活性效應,這類脂肪酶對sn-2位酰基連接位點不產生催化連接作用,因此適用于選擇sn-1和/或sn-3的酰基合成的TAG。位置專一性由脂肪酶種類和底物濃度所決定。LIN等[43]研究證實,使用sn-1,3特異性脂肪酶催化甘油和sn-2位不飽和脂肪酸含量高的甘三酯的酯交換,可以降低甘油三酯整體的飽和程度,提高不飽和脂肪酸的在甘油三酯的相對含量。第二類脂肪酶由于自身結合位點的局限性,能夠有效識別TAG的sn-1和sn-3酰基結合位置,其水解這兩類酯鍵的速度存在較大差異。立體專一性由脂肪酶來源,底物濃度和TAG連接的酰基所決定。HE等[44]分別使用Novozym 435, Lipozyme 435, Lipozyme TL-IM和Lipozyme RM IM四種sn-1,3特異性脂肪酶催化微藻油與PUFA酸解合成出富含PUFA的母乳模擬結構脂,結果表明,盡管4種特異性脂肪酶催化效果存在差異,但其均能夠較好實現對sn-1,3酰基位的有效識別。第三類脂肪酶能夠有效識別脂肪酸種類。對于不飽和脂肪酸、長碳鏈脂肪酸、中碳鏈脂肪酸以及短碳鏈脂肪酸具有差別化的催化能力。胰脂肪酶對短鏈脂肪酸具有專一性作用,Lipozyme TL IM則對中長鏈脂肪酸具有特異選擇性。

表2 常用的商業化結構脂固定脂肪酶Table 2 Common commercial lipid-immobilized lipase
3.2 長鏈多不飽和脂肪酸結構脂脂肪酶反應溫度
脂肪酶是一種活性蛋白質,其催化作用受溫度的影響較大,提高溫度可以增加酶促反應的速度。大多數sn-2位長鏈結構脂脂肪酶的最佳溫度為30~60 ℃,在此溫度范圍,脂肪酶活性最強,酶促反應速度最大。當溫度每升高10 ℃時反應速度加快一倍左右,溫度繼續升高后,高溫則會破壞脂肪酶分子的二硫鍵,導致肽鍵水解、天門冬氨酸和谷氨酰胺殘基的脫氨反應,進而降低脂肪酶在反應體系中的穩定性、親和力和競爭反應優勢,最終導致sn-2位長鏈結構脂脂肪酶失活。一般而言,動物和植物來源脂肪酶通常比胞外微生物來源脂肪酶的熱穩定差。在酯交換反應中所用的最佳溫度選擇主要是考慮對原料性質及反應體系的影響。無溶劑體系中,溫度必須保持足夠高以保持底物的液體狀態,在某些情況下溫度必須超過一定溫度才能使底物液化,此時熱穩定性更強的脂肪酶才能發揮催化作用,因此,有/無溶劑體系是結構脂合成反應選擇脂肪酶的重要考量之一。
3.3 長鏈多不飽和脂肪酸結構脂酶用量及底物組成
一般情況下,催化反應中反應速率與脂肪酶濃度成正比關系,即酶濃度越高,催化反應速率越快。但這種量效線性關系僅在一定范圍內是成立的,反應速率不會由于酶濃度的持續升高而無休止的增加。KADIVAR等[48]研究發現,隨著Lipozyme RM IM脂肪酶加入量的增加,油酸在甘油三酯中sn-2位的插入率有顯著提高,當脂肪酶添加量超過底物質量的10%時,產物的得率不再發生明顯變化。究其原因是由于脂肪酶量的增加會加快反應速度,與此同時油脂副水解反應也會相應加速。隨著脂肪酶催化作用的增強,水解作用會逐步減少酶對底物的酶解作用。此外,由于具有特異性脂肪酶的來源途徑比較單一,制備成本較高,因此脂肪酶的作用效率與產物的生產成本密切相關,這兩者均決定了結構脂在合成過程中對脂肪酶的需求量水平。
盡管底物濃度對酶的活性不產生影響,但底物濃度和底物分子構型卻能通過增加底物和酶接觸面積從而影響脂肪酶催化酯交換的速率。當底物濃度不斷增加,酶促反應速率也會逐漸加快,達到某一值后酶促反應不再隨著底物濃度的增加而增加。有研究證實[46],低底物摩爾比較高底物摩爾比需要更長的催化反應時間,后者的反應平衡會更容易向產物生成的方向進行移動,此時酰基與甘油碳鏈的結合比率也會有所提高。此外,反應過程中間產物(例如水)也會影響脂肪酶對結構脂的催化效果。在脂肪酶催化酯化反應中,反應的本質是酯化和水解反應的結合。隨著酯化過程的深入,反應中的水會逐步增加,當水含量過高時,則會誘導脂肪酶對TAG的水解反應,為了增加酯化反應減少水解,獲得高產率的產物,在反應過程中不斷地去除水分是很重要的。然而,在系統中保持一定的水分是很重要的,因為脂肪酶在非共價反應中的動力學行為需要水來維持。另一方面,有水存在的環境會阻礙脂肪酶的整體構象移動從而限制脂肪酶催化作用的發揮。因此,水解和酯化之間的底物之間的平衡關系對脂肪酶發揮催化作用十分關鍵[49]。
4 結論
相比一步酶法和三步酶法而言,兩步酶法是一種形成的副產物相對較少的理想方法,這種方法能夠很好地利用sn-1,3特異性脂肪酶酶解天然油脂,生成2-MAG,再通過脂肪酶催化純化的2-MAG和中鏈脂肪酸酯化得到純MLM型結構脂。在天然甘油三酯的所有組成類型中,當長鏈多不飽和脂肪酸位于sn-2位置時其吸收效果要優于位于sn-1,3位置或隨機分布狀態。因此,今后有必要開展將DHA、ALA等n-3長鏈多不飽和脂肪酸通過兩步酶法接入甘三酯的sn-2位合成sn-2長鏈多不飽和脂肪酸結構脂的相關研究。此外,由于脂肪酶醇解法獲得的TAG副產物較少且易分離,目標產物不飽和脂肪酸TAG的純度和產量也較高,因此是未來長鏈多不飽和脂肪酸結構脂合成路線中極具潛力的綠色高效方法。
猜你喜歡
小獼猴智力畫刊(2023年4期)2023-04-23 08:49:58
哲學評論(2021年2期)2021-08-22 01:53:34
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
中學生數理化·高一版(2018年1期)2018-02-10 05:20:03
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
七彩語文·寫字與書法(2016年7期)2016-07-28 21:40:22
七彩語文·寫字與書法(2016年6期)2016-07-15 19:36:34
人間(2015年21期)2015-03-11 15:23:21
現代企業(2015年9期)2015-02-28 18:56:50
