岑軟3號油茶落果規(guī)律及其葉片營養(yǎng)元素含量的動態(tài)變化
2020-05-14 07:46:34曾雯珺謝少義黃宏珊江澤鵬王東雪
廣西林業(yè)科學 2020年1期
關鍵詞:生長
曾雯珺,謝少義,黃宏珊,江澤鵬,王東雪
(1.廣西壯族自治區(qū)林業(yè)科學研究院 廣西油茶良種與栽培工程技術中心廣西特色經(jīng)濟林培育與利用重點實驗室,廣西南寧 530002;2.岑溪軟枝油茶種子園,廣西岑溪 543200;3.廣西平果縣同老鄉(xiāng)林業(yè)站,廣西平果 531400)
廣西是我國油茶(Camellia oleifera)三大主產區(qū)之一。近年來自治區(qū)政府從糧油安全的戰(zhàn)略角度出發(fā),在政策和財政上給予油茶產業(yè)許多支持。油茶“雙千”計劃的實施不但推動了鄉(xiāng)村產業(yè)振興,而且使廣西的油茶產業(yè)進入了高質、高效和快速發(fā)展的全新階段。但由于經(jīng)營管理水平參差不齊,油茶林中常有只見滿樹花不見滿枝果的情況,油茶落果是長期以來影響經(jīng)濟效益和制約油茶產業(yè)健康持續(xù)發(fā)展的瓶頸問題。
油茶落果與栽培技術、氣候和樹齡等密切相關,也與病蟲害和機械損傷有關[1]。研究表明,油茶落果大致可分為3個階段,早期落果、中期落果和采前落果[2]。3個階段的落果原因不盡相同,有觀點認為90%以上的早期落果為授粉和受精不完全造成的生理落果,有的認為早期果實的脫落與絕對低溫值有關,還有的認為油茶的生理特性和病蟲害發(fā)生情況共同影響早期落果;中期落果的主要原因集中在水分和養(yǎng)分的供應失調方面,也有的認為是病蟲害、絕對高溫值與光照條件等;采前落果的主要原因集中在病蟲害和機械損傷方面,個別觀點認為與養(yǎng)分和水分供應有關[1-5]。
影響油茶落果的諸多因素中,光照、氣溫和降水等氣候因素是不可控的,林地的養(yǎng)分水平則可人為調控。通過測定油茶葉片營養(yǎng)元素含量的變化,了解果實生長周期對養(yǎng)分的需求,在果實發(fā)育的關鍵時期調控樹體營養(yǎng),可減少油茶落果[6]。本研究以廣西主栽的油茶良種岑軟3號為試材,調查研究其落果規(guī)律與葉片營養(yǎng)元素含量的動態(tài)變化,旨在為油茶果實生長發(fā)育期內進行科學營養(yǎng)調控與豐產穩(wěn)產管理提供理論依據(jù)和實踐指導。
1 材料與方法
1.1 試驗地概況
試驗地設在廣西壯族自治區(qū)林業(yè)科學研究院油茶種質資源庫(108°21′E,22°56′N),地處南寧市北郊,海拔95 m,為南亞熱帶季風氣候,年均氣溫21.8℃,1月平均氣溫12.8℃,7月平均氣溫27.8℃,最低氣溫-1.5℃,最高氣溫39.4℃。年均降水量1 350 mm,平均相對濕度80%。土壤為砂頁巖發(fā)育而成的磚紅壤,pH值5~6。坡度較緩,土層深厚,土壤肥力高,陽光充足。
1.2 試驗材料
材料來自2005年種植的岑軟3號油茶試驗林。選取10株生長基本一致、正常結果且無病蟲害的植株為樣株,在樣株樹冠外圍的東、西、南、北4個方向各選定1枝30 cm以上且長勢一致的結果枝為樣枝,掛牌進行跟蹤調查。試驗林采取常規(guī)施肥和墾復等經(jīng)營管理措施,生長良好。
1.3 試驗方法
1.3.1 落果規(guī)律調查
2018年1月中旬起對供試植株的落果規(guī)律進行調查,至10月下旬油茶果實成熟,每周觀察1次,調查樣枝的果實數(shù)量并統(tǒng)計落果率。
落果率=(N0-Nt)/N0×100%,式中N0為初始落果數(shù),Nt為現(xiàn)存落果數(shù)。
利用Excel及SPSS 17.0軟件進行數(shù)據(jù)統(tǒng)計與分析。
1.3.2 葉片營養(yǎng)元素測定
在油茶果實的每個生長發(fā)育關鍵期,即幼果形成期(1—2月)、果實生長初期(3—4月)、籽粒形成期(5—7月)、果實快速生長期(7—8月)和果實成熟期(9—10月)[7-8],從標準枝上采集50片當年生老熟葉片混合,測定氮(N)、磷(P)、鉀(K)及硼(B)、鈣(Ca)、鎂(Mg)、銅(Cu)、鋅(Zn)、鐵(Fe)、錳(Mn)等元素含量。所有營養(yǎng)元素均參照LY/T 1210~1275—1999[9]進行分析測定,全N采用凱氏法測定;全P采用硝酸-高氯酸消煮,鉬銻抗比色法測定;全B采用干灰化-甲亞胺比色法測定;全K、Ca、Mg、Cu、Zn、Fe和Mn采用硝酸-高氯酸消煮,原子吸收分光光度法測定。
2 結果與分析
2.1 油茶果實的動態(tài)變化
岑軟3號油茶試驗林的總落果率(落果總數(shù)占初始果實數(shù)的比例)為70.13%,即成果率為29.87%。早期落果、中期落果和采前落果3個階段的總落果數(shù)占初始果實總數(shù)的67.53%,占整個果實生長周期總落果數(shù)的96.29%(圖1)。早期落果發(fā)生在1月中旬—3月上旬,落果率為12.99%;中期落果在4月上旬—5月下旬,落果量最大,落果率為40.26%,落果集中在4月中旬—5月中旬;采前落果在8月上旬—10月上旬,落果率為14.29%。
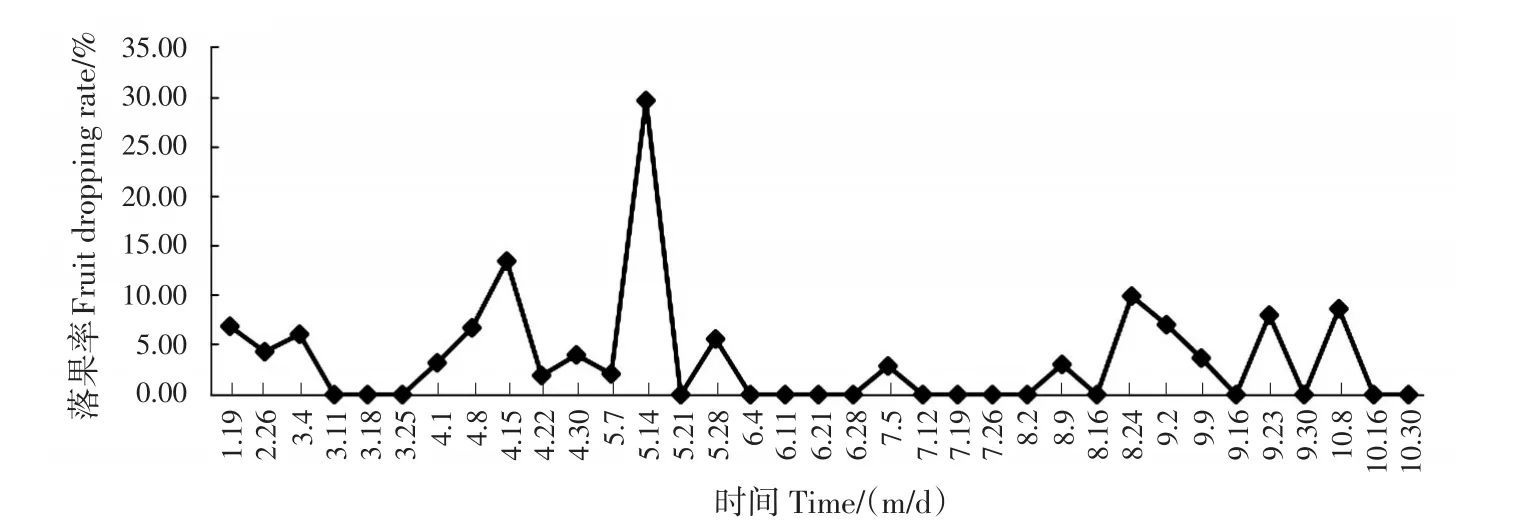
圖1 果實發(fā)育期間落果率Fig.1 Fruit dropping rate during fruit development
莊瑞林[8]將油茶果實成熟前的生長發(fā)育分為3個階段,幼果形成期、果實生長期和油脂轉化期。將落果與生長發(fā)育階段進行對比,發(fā)現(xiàn)二者基本對應。
2.2 油茶葉片營養(yǎng)元素的動態(tài)變化
2.2.1 油茶葉片中大量元素的動態(tài)變化
N是果樹合成蛋白質、核酸及各種生理活性物質的重要成分,充足的N是細胞分裂的必要條件[10-12],N含量過低或過高均會造成果實產量減少[13]。3月和7月分別為油茶春梢和夏梢的生長期,N富集在葉片中,含量相對較高;在油茶幼果形成初期(1月)、籽粒形成初期(5月)和油脂轉化期(8—9月),對N的消耗加大導致葉片中的N含量下降,在養(yǎng)分補給不充分的情況下會造成油茶落果(圖2a)。在適量的情況下,油茶果實生長與N含量成正比。油茶葉片中的N含量呈M型變化趨勢,與落果率變化曲線呈負相關關系,在一定范圍內N含量直接影響油茶落果。
P能刺激激素產生和調節(jié)激素平衡,其含量是決定花芽分化、促進果實與種子成熟及提升種子品質的重要條件[14]。油茶葉片中的P含量呈N型變化規(guī)律,其中3月和10月為P的積累期(圖2b)。幼果發(fā)育時期(3—5月)需要較多的P參與合成同化產物[15],葉片吸收的P發(fā)生轉移,含量下降;5—7月,P不僅要為花芽分化提供條件,還要滿足果實快速生長和油脂轉化的需求,含量持續(xù)下降,直至果實成熟,葉片中的P才開始累積。
K是多肽合成酶和硝酸還原酶等的活化劑,促進糖的轉運,充足的K能增加果實干重,促進其早熟[16]。果實發(fā)育期間,油茶葉片中的K含量呈單峰變化規(guī)律,1—3月K含量幾乎無變化,1月的平均含量為5 087 mg/kg,3月為5 007 mg/kg;3 — 5月K在葉片中富集,5月的平均含量(7 334 mg/kg)為1和3月的近1.5倍;5月籽粒形成后至7月為果實快速膨大的時期[8],葉片中的K向果實轉移,并繼續(xù)參與果實的油脂轉化,至10月果實成熟時,葉片中的K含量降至5 098 mg/kg,與1和3月基本持平。油茶對K的消耗與需求主要集中在幼果形成期與果實籽粒形成后至油脂轉化期2個階段(圖2c)。
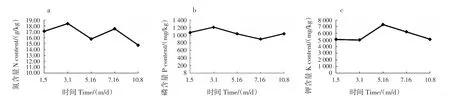
圖2 果實發(fā)育期間葉片中N、P和K含量的動態(tài)變化Fig.2 Dynamic changes of N,P and K contents in leaves during fruit development
2.2.2 油茶葉片中中量元素的動態(tài)變化
中量元素多為植物體內促進光合作用、呼吸作用及物質轉化等的酶或輔酶的組成部分,在植物體內非常活躍。作物缺乏任何一種中量元素,生長發(fā)育都會受到抑制,導致減產和品質下降,甚至絕收。
Ca是一些重要酶類的活化劑,影響籽粒形成,增加果實硬度,使果實耐貯,提高維生素C含量[17-18]。油茶葉片中的Ca含量呈先緩慢增加后急劇減少再逐漸增加的變化趨勢,其中5月的平均含量(9 204 mg/kg)較1月(19 095 mg/kg)和3月(20 861 mg/kg)減少一半以上;7月,Ca的轉移消耗減少,葉片中的Ca含量有所增加;10月,果實逐漸成熟,葉片中的Ca含量開始累積,基本達到1月的水平(圖3a)。
Mg可以促進植物的光合作用,促進蛋白質合成,提高作物產量,改善作物品質[19]。油茶葉片中的Mg含量在果實發(fā)育期間總體波動較小,為1 706~2 131 mg/kg,其中籽粒形成期(5—7月)Mg含量達到低谷,5月平均含量為1 714 mg/kg,7月為1 706 mg/kg;果實成熟期(10月)的含量達到峰值(2 131 mg/kg),高于1月(圖3b)。說明從油茶幼果形成至果實膨大對Mg均有一定程度的需求,果實快速膨大后至果實成熟Mg的參與減少,累積在葉片中。
5月為油茶果實籽粒形成初期,這時對Ca的需求尤其大,葉片中的Ca含量降至全年最低水平[7];雖然葉片中的Mg含量變化不顯著,但5—7月也向果實進行了一定程度的轉移。鑒于油茶果中籽粒的生長發(fā)育對Ca和Mg的需求,此階段應重視中量元素的施用,對于促進油茶的生長發(fā)育和結實、提升果實品質有重要意義[20]。
2.2.3 油茶葉片中微量元素的動態(tài)變化
微量元素在作物體內含量雖少,但它們是植物體內酶或輔酶的組成部分或活化物質,對開花結果及物質轉化均有很大影響,有時甚至起著決定性作用[21-22]。Cu與植物體內的氧化還原反應和呼吸作用有關,對蛋白質代謝及葉綠素的形成有重大影響,能增強光合作用,促進花粉萌發(fā)和花粉管伸長,提高結實率。Zn和Mn是許多酶的活化劑,其中Zn是微量元素中對蛋白質合成影響最大的元素,間接影響生長素的合成,同時通過對植物碳和氮的代謝產生廣泛影響,提高籽粒重量,改變籽實與莖桿的比率;Mn控制著植物體內的許多氧化還原反應,促進養(yǎng)分運往新生組織和器官,促進維生素C的形成。Fe是植物中一些重要的氧化還原酶的組分。B能促進根系生長,對碳水化合物的合成與轉運有重要作用,對受精過程的正常進行有特殊作用,有利于授粉和座果、細胞分化和伸長、糖分運輸和細胞壁中膠質合成,從而提高果實產量和貯藏品質[22-24]。
在果實發(fā)育期間,葉片中的Cu含量累積峰值(6.64 mg/kg)出現(xiàn)在3月油茶開始抽梢后,為1月Cu含量(3.13 mg/kg)的2.12倍,這是由于植株將根系中貯存和從土壤中吸收的Cu全部積累到地上部分,滿足新生器官的生長[25](圖4a)。1月油茶尚在休眠期,也是Cu含量的低谷期。3月幼果形成后至果實成熟,葉片中的Cu含量不斷下降,結合曹永慶等[25]的研究可知,這一時期油茶植株吸收的Cu幾乎全部運輸至地上部分,并不斷從葉片向果實中轉移。
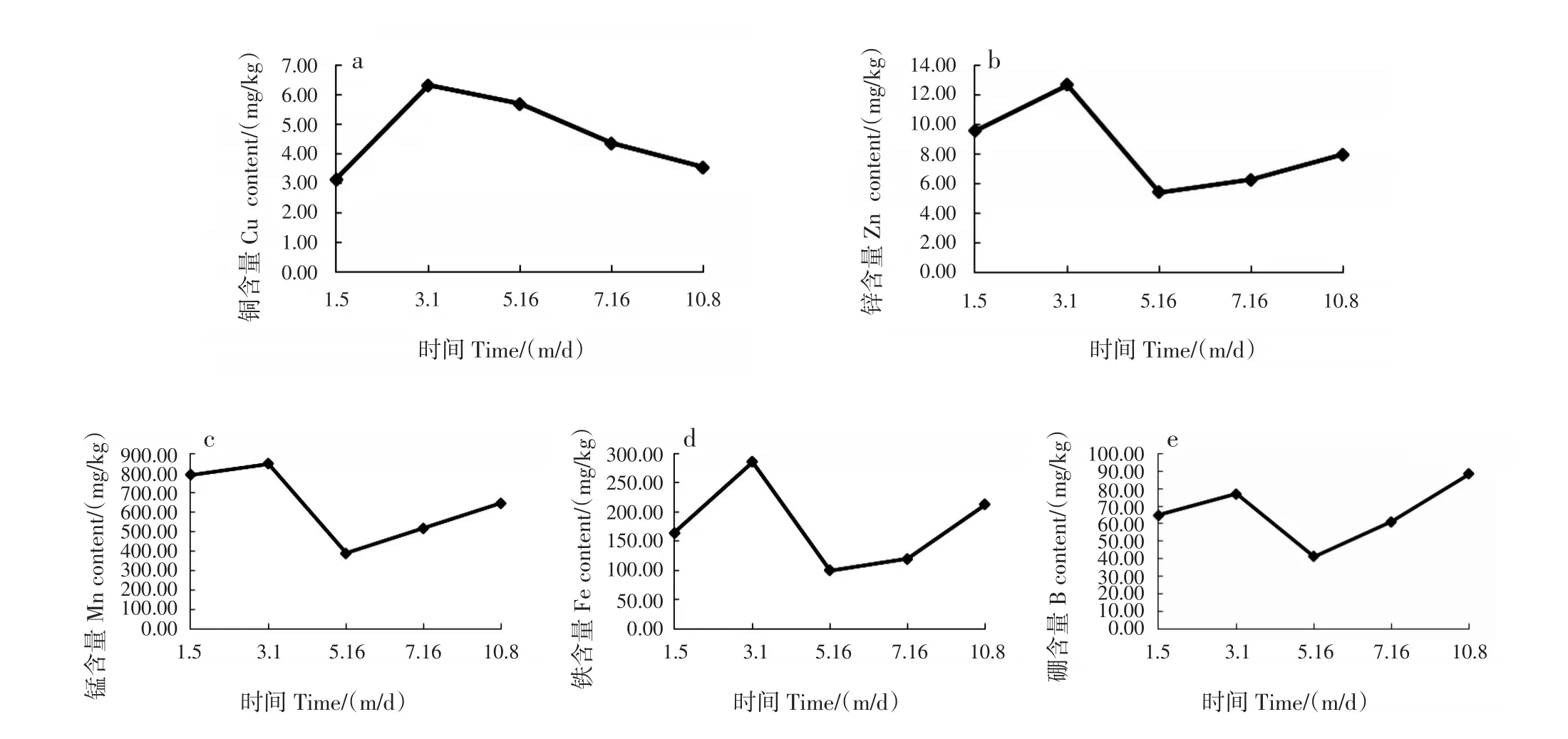
圖4 果實發(fā)育期間葉片中Cu、Zn、Mn、Fe和B含量的動態(tài)變化Fig.4 Dynamic changes of Cu,Zn,Mn,Fe and B contents in leaves during fruit development
葉片中的Zn、Mn、Fe和B含量呈N型變化趨勢,與Ca一樣拐點出現(xiàn)在3月和5月,在整個果實發(fā)育期間,Zn、Mn和Fe含量的峰值均出現(xiàn)在3月新梢大量生長時,B的峰值出現(xiàn)在果實成熟期(10月)(圖4b-e)。3月,新梢大量生長,將這些元素從根系累積到葉片中[22],之后果實生長速度逐漸加快,轉運消耗顯著增加,葉片中這些營養(yǎng)元素的含量至5月籽粒形成后到達低谷,分別為5.42、387.63、100.37和41.25 mg/kg,分別較3月減少了43%、46%、35%和53%;之后隨著轉移消耗的減少,葉片從根系中吸收的這4種微量元素含量逐漸增加,但Zn和Mn的含量仍較1月休眠期低,B在葉片中的累積較明顯,至10月果實成熟時,葉片中的累積量為5月低谷時的2.14倍,且高于1月。
2.3 油茶落果與葉片中營養(yǎng)元素的相關性
除K、Cu及大量元素總和外,油茶落果率與N、P、Ca、Mg和Zn等其余8種元素及中量元素總和、微量元素總和、養(yǎng)分總量均呈負相關關系,其含量越高,落果率越低;Ca、養(yǎng)分總量、中量元素總和及K含量與油茶落果率的相關系數(shù)絕對值相對較高,分別為0.825、0.819、0.818和0.784,說明養(yǎng)分總量的不足,尤其是Ca等中量元素含量的不足是影響油茶落果率的主要因素,同時過量的K也會導致油茶落果增加(表1)。油茶落果是由氣候和經(jīng)營水平等諸多因素協(xié)同作用的結果,這可能是本試驗營養(yǎng)元素含量與落果率之間相關性不顯著的原因。
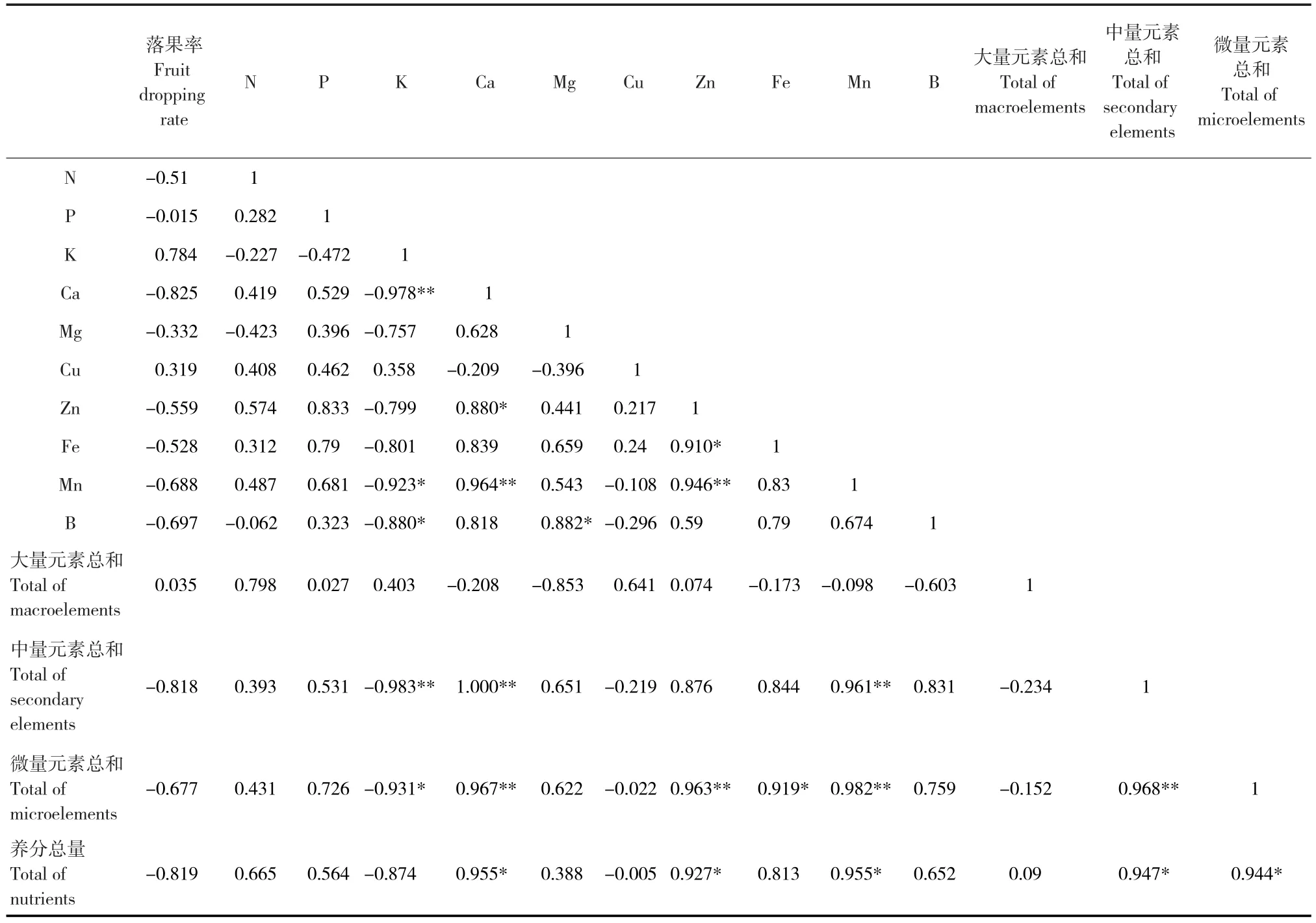
表1 油茶落果與油茶葉片中營養(yǎng)元素間的相關性Tab.1 Correlation between fruit dropping and nutrient elements in leaves of C.oleifera
3 結論與討論
觀測結果表明,岑軟3號油茶有3個落果階段,這與其他研究人員對油茶落果的觀測結果一致[1-3],但3個落果階段發(fā)生的時間和落果率不同。本研究中3個落果階段分別發(fā)生在1月上旬—3月上旬、4月上旬—5月中旬和8月上旬—采前,落果率分別為12.99%、40.26%和14.29%。周律相[2]在3、6和9月調查廣西油茶的落果率,分別為58%、24%和33%;卯吉華等[3]的研究發(fā)現(xiàn),油茶在云南高原山區(qū)的落果發(fā)生在4月初—5月初、5月中旬—7月下旬和7月下旬—9月下旬,落果率分別為32.5%、59.6%和7.9%。本研究中3個落果階段發(fā)生的時間相對較早,這與調查統(tǒng)計方法、樣地氣候條件及油茶林齡等因素相關;前者的研究結果顯示落果率最高的時期為早期落果階段,本研究落果率最高的階段與云南高山區(qū)的結果一致,均為中期落果的落果率最高,這是由于前者所調查的樣地經(jīng)營管理水平不高,普遍存在缺肥少肥的情況,而后2個樣地采取了施肥和墾復等經(jīng)營措施;此外,本研究中的試驗林為13a生的壯年林,處于生理落果率最低的林齡階段[1,4]。
油茶落果大多是由水分和養(yǎng)分供應失衡造成。在幼果形成初期,降水量大造成柱頭溶液濃度降低和花粉提前脫落,導致受精不完全,本研究的試驗地從1月開始逐漸形成幼果,此時為1年中的旱季,因此造成落果的主要原因不是水分條件;果實形成初期,植物體需要消耗N和P為果實生長提供活性物質,因此葉片中的N和P含量相對較低,此時由養(yǎng)分不足引起的花器官發(fā)育不完全和花期短,均會影響油茶的授粉與受精,進而導致油茶生理落果[3]。
3月進入果實生長期后,油茶果實的生長逐漸加快,對水分和養(yǎng)分的需求逐漸增大,此階段的落果與降水量大致呈負相關關系,3—4月落果相對較多,降水量大的6—7月落果減少,這與陳育松等[1]的研究結果一致。3月油茶開始進入中期落果階段,此時除K外,其他養(yǎng)分含量充足,降水量也有所增加,但由于3月春梢生長和4—5月花芽分化,一定的養(yǎng)分和水分競爭造成落果增加;降水量在6—7月達到1年中的峰值(223.1和337.8 mm)[26],而此時還未到果實的迅速膨大期,對養(yǎng)分需求不高且水分充足,落果情況開始改善。6月為果實感染炭疽病的高峰期[27],部分落果由病蟲害的原因造成。
采前落果的影響因素多而復雜,陳育松等[1]和舒金平等[4]認為油茶采前落果的主要原因為茶籽象蟲(Curculio chinensis)和油茶炭疽病(Colletotrichum gloeosporioides)等病蟲害及機械損傷,且與種植年限密切相關;朱英芝等[28]研究發(fā)現(xiàn),廣西油茶炭疽病病原菌最適生長溫度為28℃,最適產孢溫度為32℃,因此8—9月為發(fā)病的第2個高峰期;油茶象甲成蟲在6月下旬—7月上旬產卵于油茶種內或種仁間隙,被象甲蟲寄生的果實在果長、果徑、果重及籽重方面明顯小于未受害果,7月中旬—9月中旬幼蟲脫果而出,果實在成熟前落下[29]。
本試驗中K和Cu與落果率呈正相關關系,說明過量的K和Cu會導致油茶落果,對這2種營養(yǎng)元素的補充需適量。資料顯示,過量的K會造成植物體內Ca等營養(yǎng)元素與之發(fā)生拮抗作用,導致產量降低[30-32]。本試驗中K與Ca和B呈極顯著和顯著負相關,過量的K抑制葉片對Ca和B的吸收,驗證了3種元素間的拮抗關系。本試驗中,Ca與Zn和Mn呈顯著和極顯著正相關,這與張?zhí)熘镜萚33]和于婷等[34]的研究結果一致;Ca還與中量元素總和、微量元素總和及養(yǎng)分總量呈極顯著和顯著正相關,說明適量的Ca對多種營養(yǎng)元素的吸收均有促進作用。許建楷等[35]和于澤源等[36]認為Ca和K能使樹體養(yǎng)分失衡,是對裂果影響最突出的營養(yǎng)元素,在今后的研究中,可進一步探索K和Ca的調控對油茶落果的影響。本試驗中,Zn和Mn與多種元素、元素總和和養(yǎng)分總量呈顯著和極顯著相關,在對油茶進行營養(yǎng)調控時不可忽視它們的重要作用。
岑軟3號油茶的成果率為29.87%。3個落果階段分別發(fā)生在1月上旬—3月上旬、4月上旬—5月下旬和8月上旬—10月上旬,落果率分別為12.99%、40.26%和14.29%。
油茶在果實不同的生長發(fā)育關鍵期對養(yǎng)分元素的需求不同。1月油茶處于休眠期,植株從根系中獲得的養(yǎng)分有限,幼果的形成消耗了一定量的N、P、K、Ca、Cu、Zn、Mg、Fe和B,此時葉片中這些元素的含量較3月均處于相對較低的水平。3月后果實進入生長期,樹液開始流動,新梢大量生長,除了K和Mg,其余養(yǎng)分元素均在葉片中累積,可能是因為果實的生長發(fā)育需要K和Mg。3—5月,果實生長速度加快,葉片中的N、P、Ca、Mg、Cu、Zn、Mn、Fe和B均向果實中轉移,為其生長發(fā)育提供養(yǎng)分,只有K累積在葉片中。5—7月油茶果實中形成籽粒并開始逐漸膨大,果實生長對P、K和Cu的依賴較高,葉片中這3種元素的含量減少,而N、Ca、Mg、Zn、Mn、Fe和B在葉片中累積,可能是因為7月為夏梢生長期,對這些元素的需求較大。7—10月,果實進入快速膨大期,開始油脂轉化并逐漸成熟,這一階段葉片中的N、K和Cu含量明顯降低,說明這3種元素更多地參與了果實的發(fā)育和成熟;對P、Ca、Zn和Mn的消耗轉移相對較少,其在葉片中的含量出現(xiàn)小幅回升;Mg、Fe和B在這一時期快速累積在葉片中,可以認為這3種元素不是該階段的關鍵因素。
綜合以上情況,在對岑軟3號油茶林進行營養(yǎng)調控時,建議冬季休眠前的施肥應對本研究測定的所有營養(yǎng)元素進行不同程度的補充;夏季施肥應適當補充N、P、K、Cu、Ca、Zn和Mg。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現(xiàn)代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
