鹽脅迫對葡萄砧木‘3309C’葉片光抑制的影響*
2020-05-16 07:21:14高玉錄仝亞軍張成超孫永江杜遠鵬
中國果樹 2020年1期
高玉錄,仝亞軍,張成超,孫永江,杜遠鵬,翟 衡
(山東農業大學園藝科學與工程學院,泰安 271018)
我國鹽漬土資源量多且分布廣泛,鹽堿地的改良利用是涉及農業可持續發展的重大問題,目前耐鹽‘海水稻’已經進入產業規模示范階段。葡萄是一種相對較耐鹽的果樹樹種,也是鹽堿地區農民喜歡種植的高效經濟作物之一,而葡萄砧木在抵御逆境、病蟲等方面發揮重要作用,利用砧木進行嫁接栽培已成為葡萄產業發展的必然趨勢。
植物光合作用對逆境脅迫非常敏感,可以用作反映逆境脅迫的有效探針。因此,研究鹽脅迫對葡萄砧木光合作用的影響,對揭示砧木的抗鹽機制具有重要的理論意義。植物吸收的光能超過光合作用所能利用的限度而導致光合效率下降的現象叫光抑制,它包括光系統II(PSII)光抑制和光系統I(PSI)光抑制。通常認為PSII 是逆境脅迫光抑制的主要位點[1],PSII 最大光化學效率(Fv/Fm)的下降是PSII 發生光抑制的重要指標[2]。相比PSII,一般情況下PSI 不易發生光抑制,但研究表明鹽脅迫情況下PSI 卻是容易傷害位點[3-4]。植物環式電子傳遞(CEF)由于具有產生跨膜質子梯度并只合成ATP且無還原力(NADPH)的形成的特點,在短期鹽脅迫過程中起到了重要作用[5],被認為是抵御逆境脅迫的重要防御機制[6-7],但缺少對長期鹽脅迫下葡萄CEF 活性變化觃律的報道。本研究以葡萄優良抗性砧木‘3309C’為試材,對盆栽苗進行澆鹽處理,模擬不同持續時間的鹽脅迫,研究了植株光合特性對鹽脅迫的響應,以期為葡萄抗鹽栽培提供理論基礎。
1 材料與方法
1.1 材料與處理
試驗于2017 年4—9 月在山東農業大學園藝實驗站葡萄園基地內進行,選取葡萄砧木品種‘3309C’1 年生苗為試驗材料,春季種植到觃格為外口直徑23.5 cm、內口直徑20.1 cm、高14.0 cm、底部直徑12.9 cm 的塑料盆中,基質為中性壤土、沙及有機肥按照的比例配制而成,待生長到6~7 片葉片時,選取長勢一致的植株,用濃度為100 mmolL 的NaCl 溶液澆灌植株,每株澆1 L,每隔2 d 澆灌1 次,對照澆等量清水,每3 株為1個處理,重復3 次。
分別在澆灌處理前、處理7 d(澆灌4 次)和處理14 d(澆灌8 次)選擇中部節位長勢一致的葉片進行葉綠素熒光參數測定。
1.2 葉綠素熒光參數的測定
使用連續激發式熒光儀(Handy PEA,Hansatech,英國)測定暗適應下的葉片快速葉綠素熒光誘導曲線。Fo為最小熒光(20 μs);Fk為K 點(300 μs)的熒光;FJ為J 點(2 ms)的熒光;Fm為最大熒光即P 點的熒光。通過JIP-test 分析可得到如下參數:


使用Dual-PAM100 葉綠素熒光儀(Walz,Germany)進行處理的活體測定。首先對經過一晚上暗適應的葉片進行Fm的測定,然后將葉片置于471 μmol·m-2·s-1的光下進行充分光啟動后測定光下熒光參數:
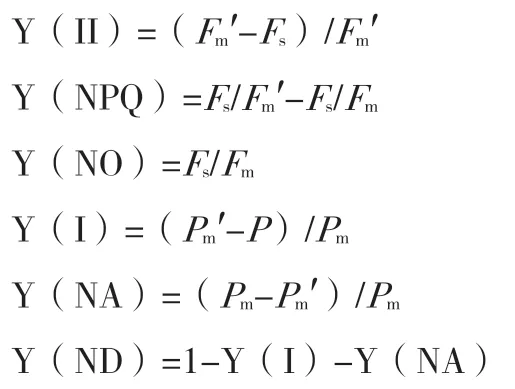
Y(II)表示PSII 的有效量子產率,Y(NPQ)和Y(NO)分別表示PSII 中調節和非調節能量耗散的量子產額,Y(NA)和Y(ND)分別表示由于PSI電子受體側和供體側限制引起的非光化學能量耗散的量子產額。Fm′表示光適應后的最大熒光,Fs是光適應后的穩態熒光,Pm為最大光氧化P700,Pm′表示光下PSI 最大熒光。

β/α-1 表示PSI 和PSII 間激發能分配的不平衡性[8]。
作用光關閉后葉綠素熒光瞬時上升測定:同樣使用Dual-PAM 100 葉綠素熒光儀進行作用光關閉后葉綠素熒光瞬時上升的測定。選取長勢一致的經過30 min 暗適應的葉片,測定開始后,先用小于0.1 μmol·m-2·s-1的調制測量光測定暗中最小熒光,然后再用大于7 000 μmol·m-2·s-1測定最大熒光,接著再用200 μmol·m-2·s-1活化光,此時熒光信號會隨時間逐漸衰減,3 min 后關閉活化光,熒光信號降低后再次瞬時上升,同時繼續記錄此時熒光信號的變化。
P700+再還原速率的測定:利用Dual-PAM 100葉綠素熒光儀進行葡萄葉片P700+再還原速率的測定,經過30 min 暗適應的植物被遠紅光(>705 nm,5.2 μmol·m-2·s-1)照射使P700+充分氧化,經過30 s 照射后,熒光信號充分平穩。然后再關閉遠紅光使P700+再還原,并記錄P700+再還原曲線的速率和時間,計算其半衰期(T1/2)。
1.3 數據分析
試驗數據用SPSS 13.0 軟件進行單因素方差分析,并進行LSD 多重比較。用Excel 進行數據分析以及作圖,數據以平均值±標準誤表示。
2 結果與分析
2.1 鹽脅迫對PSII 及PSI 活性的影響
PSII 最大光化學效率可以反映PSII 受脅迫程度。從圖1 可以看出,與對照相比,100 mmol/L 的NaCl 溶液處理到第7 d 時Fv/Fm開始出現降低,比對照下降了4.3%,第14 d 時出現顯著下降,相比對照下降了13.6%。最大光氧化P700 可以反映PSI 活性,由圖2 可知,脅迫處理第7 d 便導致了Pm值出現了顯著降低,相比對照下降了14.7%,第14 d 下降了41.3%。上述結果表明,鹽脅迫抑制了葡萄葉片PSII 和PSI 活性,而相比PSII,‘3309C’葉片PSI的鹽脅迫耐受能力較弱。
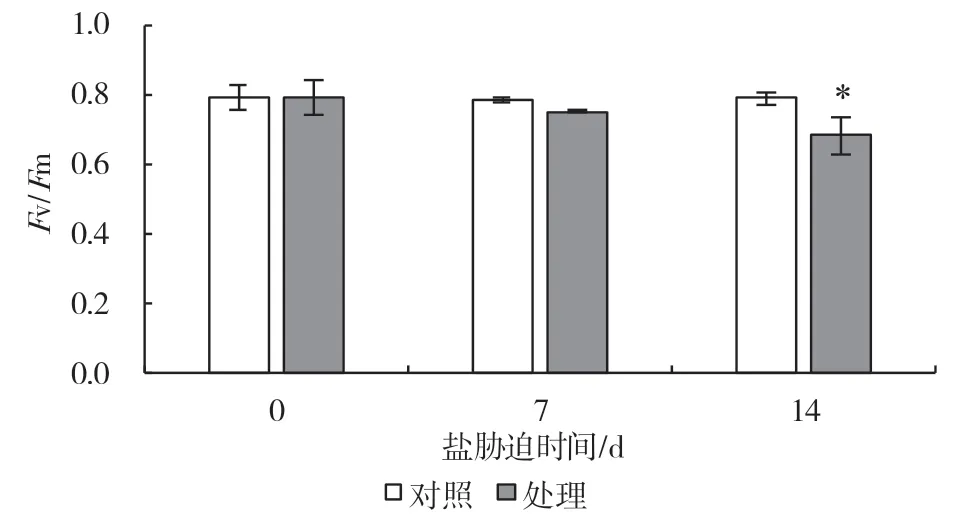
圖1 鹽脅迫對葡萄砧木葉片PSII 最大光化學效率的影響

圖2 鹽脅迫對葡萄砧木葉片最大光氧化P700 的影響
2.2 鹽脅迫對葉綠素熒光曲線及參數的影響
通過分析葉綠素熒光多項上升動力學曲線可以獲得與光合原初反應有關的豐富信息。從圖3 可以看出,隨著鹽脅迫處理時間的延長,葡萄砧木葉片O-J-I-P 曲線發生變化。長期鹽脅迫導致葉片熒光曲線發生明顯變化,Fm值發生明顯降低,說明鹽脅迫下葉片單位面積吸收的光能明顯降低。
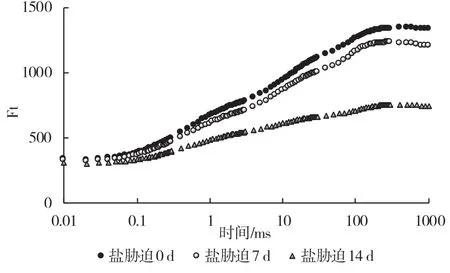
圖3 鹽脅迫下葡萄砧木葉片不同時間段穩態熒光變化
對植物快速葉綠素熒光動力學曲線參數進行進一步的分析,可以獲得大量關于PSII 原初光化學反應的信息。K 點相對可變熒光(Wk)可以表示PSII供體側放氧復合體的受傷害程度。由圖4 可以看出,隨著鹽處理時間的增加,Wk值也在逐漸升高,到第14 d 有顯著升高,上升幅度為14.8%,表明葡萄葉片PSII 供體側受到了鹽脅迫的傷害。而單位面積有活性反應中心數量(RC/CSm)及捕獲的電子傳遞至QA以后的概率(ΨEo)在處理至第7 d 時較對照都有所降低,下降的幅度分別為7.5%、14.3%,第14 d出現顯著下降,下降幅度分別為45.4%、30.3%(圖5、6)。這都表明鹽脅迫傷害了PSII 光合電子傳遞鏈。
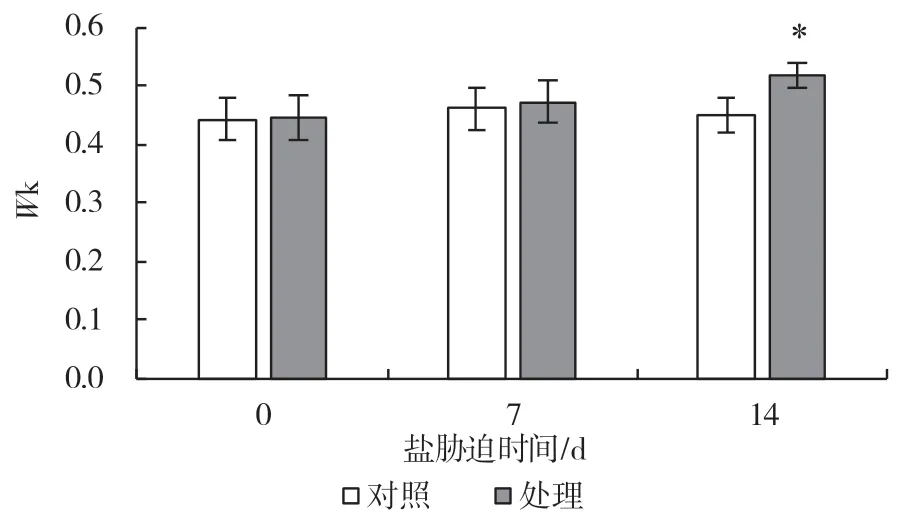
圖4 鹽脅迫對葡萄砧木葉片K 點相對可變熒光的影響

圖5 鹽脅迫對單位面積活性反應中心數量的影響

圖6 鹽脅迫對捕獲的電子傳遞至QA以后的概率的影響
2.3 鹽脅迫對光系統能量分配的影響
鹽脅迫改變了PSII 和PSI 的能量分配。由圖7可知,隨著鹽處理時間的延長,PSII 可調節的熱耗散Y(NPQ)逐漸升高,第14 d 與對照相比差異顯著,上升了12.3%;而不可調節的熱耗散Y(NO)雖有所上升但差異不顯著,表明PSII 活性受到了抑制。與此同時,PSI 氧化態P700 即Y(ND)逐漸降低,第14 d 與對照相比差異顯著,下降了12.3%,PSI 供體側受抑制程度明顯增加,而還原態P700+即Y(NA)逐漸升高,第7 d 即出現顯著升高,升高了23.5%,第14 d 相比對照升高了127.3%,說明PSI 受體側受抑制程度明顯增加,進一步表明鹽脅迫導致了PSI 光抑制的發生。
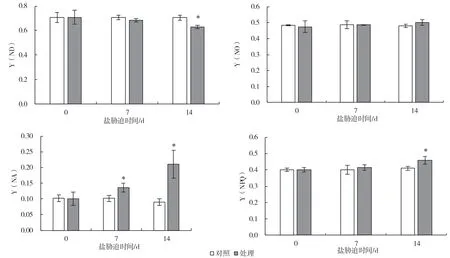
圖7 鹽脅迫對葡萄砧木葉片PSII 和PSI 光能利用效率的影響
利用PSII 熒光產量進一步分析2 個光系統之間的激發能分配,從圖8 可以看到,隨著鹽脅迫處理時間的延長(β/α-1)的值逐漸增大,與對照處理相比,第7、14 d 處理后分別增加了8.4%和75.8%,說明鹽脅迫導致了葡萄葉片PSII 和PSI 間激發能分配不平衡性的增大。
2.4 鹽脅迫對環式電子傳遞的影響
通過檢測作用光關閉后葉綠素熒光瞬時上升可以檢測NDH 途徑的環式電子傳遞能力的大小,從圖9 可以看出,隨著鹽處理時間的延長,作用光關閉后的熒光上升幅度及斜率均逐漸降低,處理第7 d 就出現明顯下降,表明長時間的鹽處理抑制了依賴NDH 途徑的CEF 活性。

圖8 鹽脅迫對葡萄砧木葉片PSII 和PSI 間激發能分配不平衡性的影響
P700+再還原動力曲線的半衰期能反映環式電子傳遞的速率[9],如圖10 所示,隨著鹽處理的進行,第7 dT1/2值出現了顯著升高,比對照升高了11.2%,第14 d 比對照升高了30.7%,結果進一步表明長時間鹽處理抑制了葡萄CEF 的活性。
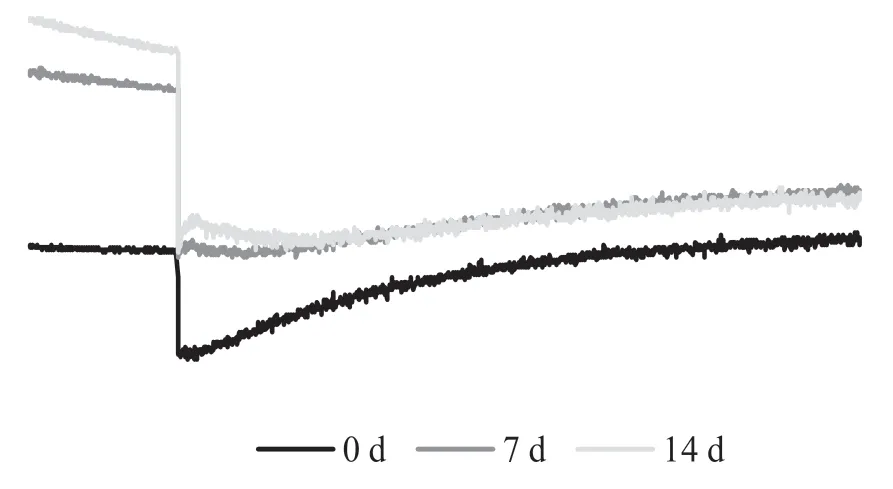
圖9 鹽脅迫作用光關閉后葉綠素熒光瞬時曲線
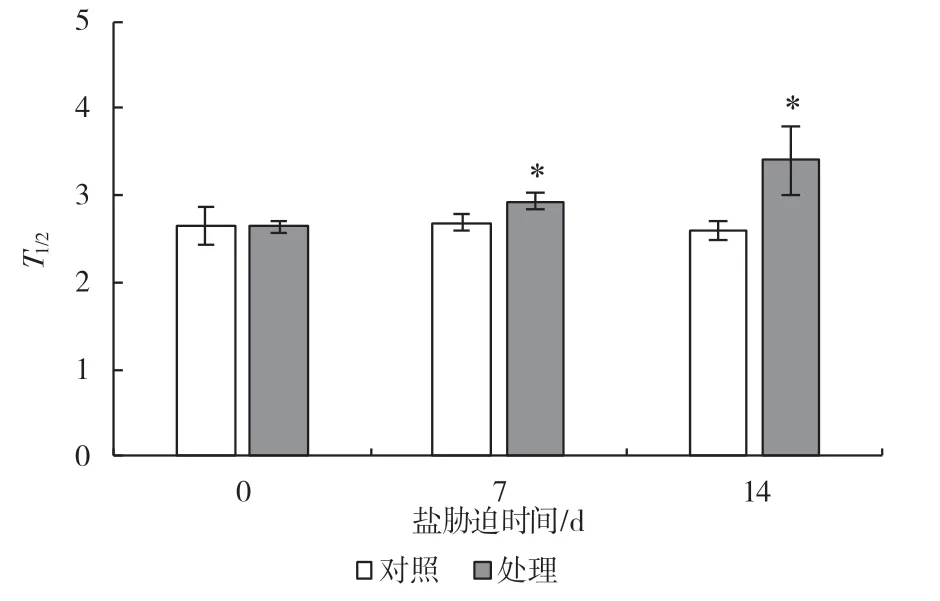
圖10 鹽脅迫對P700+再還原動力曲線半衰期的影響
3 討論與結論
葉綠體是植物進行光合作用的細胞器,對葉片的葉綠素熒光參數進行分析能夠反映脅迫條件下植物光合器官的傷害程度。光合機構由PSI 和PSII組成,當植物吸收的光能超過所能利用的光能時,過剩光能便會導致光合機構的光抑制,甚至造成光破壞[10]。研究表明氯化鈉鹽脅迫導致葉綠素發生降解[11],引起植物光系統活性發生抑制[4,10,12]。本研究中,隨著氯化鈉鹽處理時間的增加,葉片最大光化學效率及最大光氧化P700 均逐漸降低,且最大光化學效率發生降低時間早于最大光氧化P700,表明鹽脅迫抑制了葡萄葉片光系統活性,其中PSI 相比PSII 更容易受到鹽脅迫的抑制,這與顏坤等[13]在金銀花上的研究結果一致。
光合作用的高效率進行依賴于PSII 和PSI 的協調運轉,從而確保光合色素吸收的光能在2 個光系統間的均衡分配[10]。鹽脅迫下葉片2 個光系統間激發能分配失衡,葉片不能通過狀態轉換有效地將吸收的光能從PSII 傳遞給PSI,導致了葡萄葉片K 點相對可變熒光逐漸升高,而用于電子傳遞量子產額、單位面積有活性反應中心的數量均表現出下降趨勢,表明鹽脅迫抑制了葡萄葉片PSII 供體側和受體側導致PSII 電子傳遞受到抑制。隨著處理時間的延長,葉片可調節的能量耗散Y(NPQ)逐漸升高,一方面表明其接受的光強過剩,另一方面說明葉片啟動了與高能態淬滅有關的葉黃素循環過程并不能耗散掉所有的光能,隨著鹽脅迫時間的延長,過剩光能的比例越來越高,非調節性能量耗散Y(NO)逐漸升高,葉片發生嚴重光抑制[14]。
研究表明圍繞PSI 的環式電子傳遞在逆境脅迫下對穩定PSI 活性起到了重要的作用,進而有助于PSII 的保護和修復[15]。植物體內CEF 主要起到了2方面作用:通過NPQ 進行熱耗散[16-17]以及提供ATP進行CO2固定[18]。作用光關閉后熒光瞬時上升能夠反映依賴NDH途徑的CEF的活性[19],研究發現NDH途徑在光保護過程中起重要作用[20],能夠減輕逆境脅迫下葉綠體ROS 的積累及基質的過度還原[21]。隨著鹽脅迫處理時間的延長,葡萄葉片葉綠素熒光瞬時上升幅度和斜率均降低,表明長時間的鹽處理抑制了NDH 途徑。P700+再還原速率可以用來反映CEF 活性[22]。本研究中,鹽脅迫下伴隨光抑制的加劇,P700+再還原動力曲線的半衰期逐漸增大,這些結果說明長期鹽脅迫下,CEF 活性的抑制可能是導致能夠通過對葉綠體基質的過度還原,增加ROS 的累積,加劇光抑制程度。長期鹽脅迫導致葡萄砧木2 個光系統激發能分配嚴重偏離平衡狀態,抑制了光系統活性,更多的能量被用來進行熱耗散,而圍繞PSI 的環式電子傳遞活性的抑制則進一步加劇了葡萄葉片光系統光抑制程度。
