法尼醇對白念珠菌生物膜死亡作用的研究初探
2020-06-06 01:53:50姜劉鎏孫凱瑞
口腔醫學 2020年5期
周 鵬,章 萍,姜劉鎏,孫凱瑞,魏 昕
白念珠菌是最常見的條件致病性真菌[1],宿主免疫系統下降時可引起感染。白念珠菌在口腔、皮膚等部位主要以生物膜狀態存在,與浮游狀態白念珠菌比較,生物膜狀態白念珠菌對抗真菌藥物的耐受度明顯增高[2]。
法尼醇是一種從真菌中檢測出來的群體感應分子[3]。它可抑制白念珠菌細胞由酵母相向菌絲相的轉換,從而抑制生物膜的形成及成熟[4]。先前研究表明法尼醇可以誘導白念珠菌浮游狀態及生物膜狀態細胞死亡[5-6],可促進耐藥株白念珠菌浮游狀態細胞壞死[6]。誘導白念珠菌細胞死亡可能對控制白念珠菌特別是耐藥白念菌株的機會性感染有重要意義。法尼醇誘導白念珠菌生物膜的具體死亡機制尚不明確,未見法尼醇對白念珠菌耐藥株生物膜壞死或凋亡作用的比較研究。
CDR1是編碼氟康唑耐藥相關的ABC轉運蛋白基因[7]。文獻發現,法尼醇導致浮游狀態白念珠菌CDR1表達顯著增加;在法尼醇作用下,浮游狀態CDR1缺陷株白念珠菌細胞死亡率較低,過表達CDR1白念珠菌則死亡率升高[8],MCA1編碼Caspase酶,Caspase酶與真核細胞凋亡密切相關[9]。基于上述研究發現,本研究對法尼醇誘導白念珠菌耐藥株和標準株生物膜死亡的作用是壞死還是凋亡進行初步探討。研究假設:法尼醇可誘導白念珠菌生物膜凋亡或壞死,其作用與生物膜時相及法尼醇作用濃度相關;CDR1和MCA1基因表達與法尼醇對白念珠菌生物膜細胞凋亡或壞死的調控作用相關。
1 材料與方法
1.1 菌株
白念珠菌標準株(SC5314)購自美國標準生物品收藏中心。白念株菌耐藥株由SC5314通過氟康唑濃度梯度法誘導獲得(已在過往研究中使用[10]),采用KONT真菌顯色MIC藥敏系統穩定性檢測實驗條鑒定,對抗真菌藥物氟康唑(MIC ≥64 μg/mL)、5氟胞嘧啶(MIC ≥32 μg/mL)、兩性霉素B(MIC ≥2 μg/mL)、伊曲康唑(MIC ≥1 μg/mL)耐藥。
1.2 試劑與儀器
沙堡瓊脂培養基(SDA,6.3 g/100mL),YPD培養液(2%葡萄糖2 g/100 mL,1%胰蛋白胨1 g/100 mL,1%酵母提取物1 g/100 mL),RPMI1640培養基(GIBCO公司,美國),PBS(pH=7.2,0.2 g/L KCL,0.2 g/L,8.0 g/L NaHCO3,2.16 g/L Na2HPO4·7H2O),E,E-法尼醇(100 μmol/L,Sigma公司,美國),PI試劑(YEASEN公司,中國),annexin V-FITC/PI(寶賽公司,中國),蝸牛酶(YEASEN公司,中國),DEPC水(Sigma公司,美國),iTaq-SYBR Green 預混酶(Roche公司,瑞士),玻璃底共聚焦培養皿(NEST公司,中國),激光共聚焦(Carl Zeiss公司,德國),超微量分光光度計(Nanoplus公司,德國),ABI 7300 FAST熒光定量PCR儀(ABI公司,美國),ABI Prism 7300SDS分析軟件,流式細胞分析儀(BD公司,美國),YCP系列二氧化碳培養箱(易亮醫療器械有限公司,中國),低溫高速離心機(EPPENDORF公司,德國)。
1.3 分組
白念珠菌標準株和耐藥株生物膜法尼醇未處理對照組;白念珠菌標準株和耐藥株生物膜法尼醇處理組(50、100、200、400、800 μmol/L)。
1.4 構建白念珠菌標準株及耐藥株生物膜
將10 mL白念珠菌標準株和耐藥株菌懸液(1×106個/mL)分別接種至10 cm玻璃底培養皿中。37 ℃,5% CO2孵育2 h。法尼醇未處理組每隔2 h換置新鮮的10 mL RPMI1640培養液。法尼醇處理組每隔2 h換置新鮮的10 mL相應梯度濃度法尼醇的RPMI1640培養液。分別培養6、12、24 h,獲得不同濃度法尼醇處理下,不同時相的白念珠菌標準株和耐藥株生物膜。
1.5 PI染色觀察法尼醇對白念珠菌生物膜的壞死作用
取2 mL白念珠菌標準株及耐藥株標準菌懸液分別接種至2 cm玻璃底共聚焦皿中,37 ℃,5% CO2恒溫培養箱培養,獲得各濃度法尼醇處理組和未處理對照組白念珠菌耐藥株和標準株的6、12、24 h生物膜。4%多聚甲醛固定后,每皿加入50 μL PI(50 μg/mL),避光5 min,激光共聚焦顯微鏡下(× 400)觀察法尼醇作用后耐藥株和標準株壞死的形態學變化,激發光為535 nm。
1.6 Annexin-V-FITC/PI流式分析法尼醇對白念珠菌生物膜凋亡和壞死的作用
1.6.1 制備原生質體 按上述方法培養不同濃度法尼醇處理組和未處理對照組的白念珠菌耐藥株和標準株6、12、24 h生物膜,收集后分別置于1.5 mL EP管中,12 000×g離心5 min。棄上清,加入1 mL脫壁促進劑(50 mmol/L二硫蘇糖醇(DTT)+50 mmol/L 乙二胺四乙酸(EDTA)+50 mmol/L Tris(pH 8.0)),30 ℃,30 min后PBS洗滌2次,再加入1 mL 1%蝸牛酶,30 ℃,30 min,PBS洗滌2次,得到白念珠菌脫壁后原生質體的懸浮液。
1.6.2 流式細胞技術分析 收集白念珠菌生物膜原生質體,70%乙醇,4 ℃固定1 h。對標本進行標記后,使用膜聯蛋白Annexin-V-FITC/PI凋亡檢測試劑盒進行處理,加入FITC標記的Annexin V,室溫避光30 min,加入PI,避光5 min,加入Buffer,之后在流式細胞儀上分析。分別計算正常、早期凋亡、晚期凋亡、壞死細胞的百分比。
1.7 q-PCR法分析法尼醇對白念珠菌生物膜CDR1及MCA1基因表達的改變
采用改良熱酸酚法[11]提取總RNA,根據PrimeScript RT reagent Kit逆轉成cDNA,超微量分光光度計測定RNA濃度,-20 ℃保存。
q-PCR使用iTaq-SYBR Green 預混酶進行反應,20 μL反應體系:10 μmol/L的上下游引物各0.6 μL,20倍稀釋的cDNA模板0.2 μL,8.6 μL滅菌蒸餾水,再避光加入10 μL預混酶。將配制好的cDNA樣本加入96孔培養板,ABI 7300 FAST熒光定量PCR儀進行q-PCR反應。循環條件:95 ℃,10 min,隨后95 ℃變性15 s,55 ℃ 退火1 min,40個循環;70 ℃,20 s;最后4 ℃冷卻。ACTIN為陰性對照基因。qRT-PCR 完成后,在 ABI 7300 System SDS Software 1.3.1.21軟件上分析,查看每個基因的擴增情況,記錄相應的Ct值。所有樣本均重復3次。引物序列見表1。
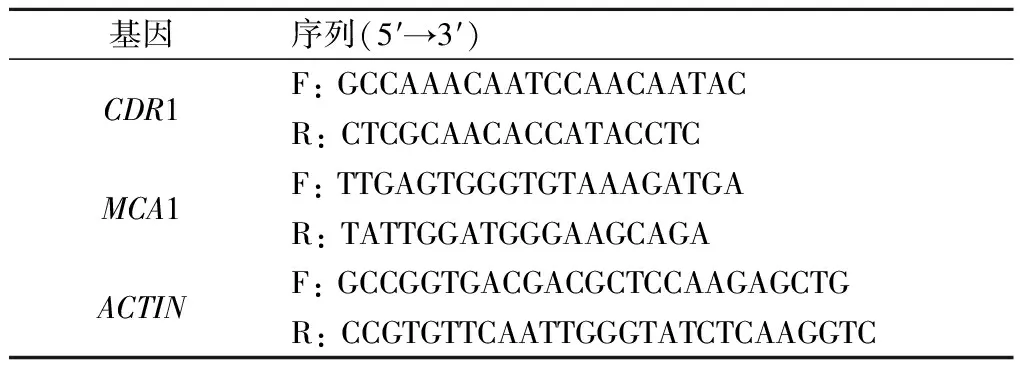
表1 引物序列Tab.1 Sequences of primers
1.8 統計方法
本研究使用SPSS 17.0軟件進行統計學分析。對流式分析數據采用t檢驗分析,對q-PCR基因檢測數據采用單因素方差分析。P<0.05為差異有統計學意義。
2 結 果
2.1 激光共聚焦下觀察法尼醇促進白念珠菌生物膜細胞的壞死
隨著法尼醇處理濃度的升高,白念珠菌耐藥株和標準株生物膜中細胞密度降低,菌絲長度變短,壞死細胞數目增加。標準株及耐藥株各時相鏡下表現具有一致性。結果參見圖1。

白念珠菌標準株和耐藥株生物膜細胞經法尼醇處理后,細胞膜完整性破壞。PI染色后,壞死細胞在熒光顯微鏡下觀察為紅色
2.2 流式分析觀察法尼醇對白念珠菌生物膜凋亡和壞死的作用
在不同生物膜時相下,法尼醇對白念珠菌耐藥株和標準株生物膜細胞凋亡與壞死的作用存在差異。結果見圖2。
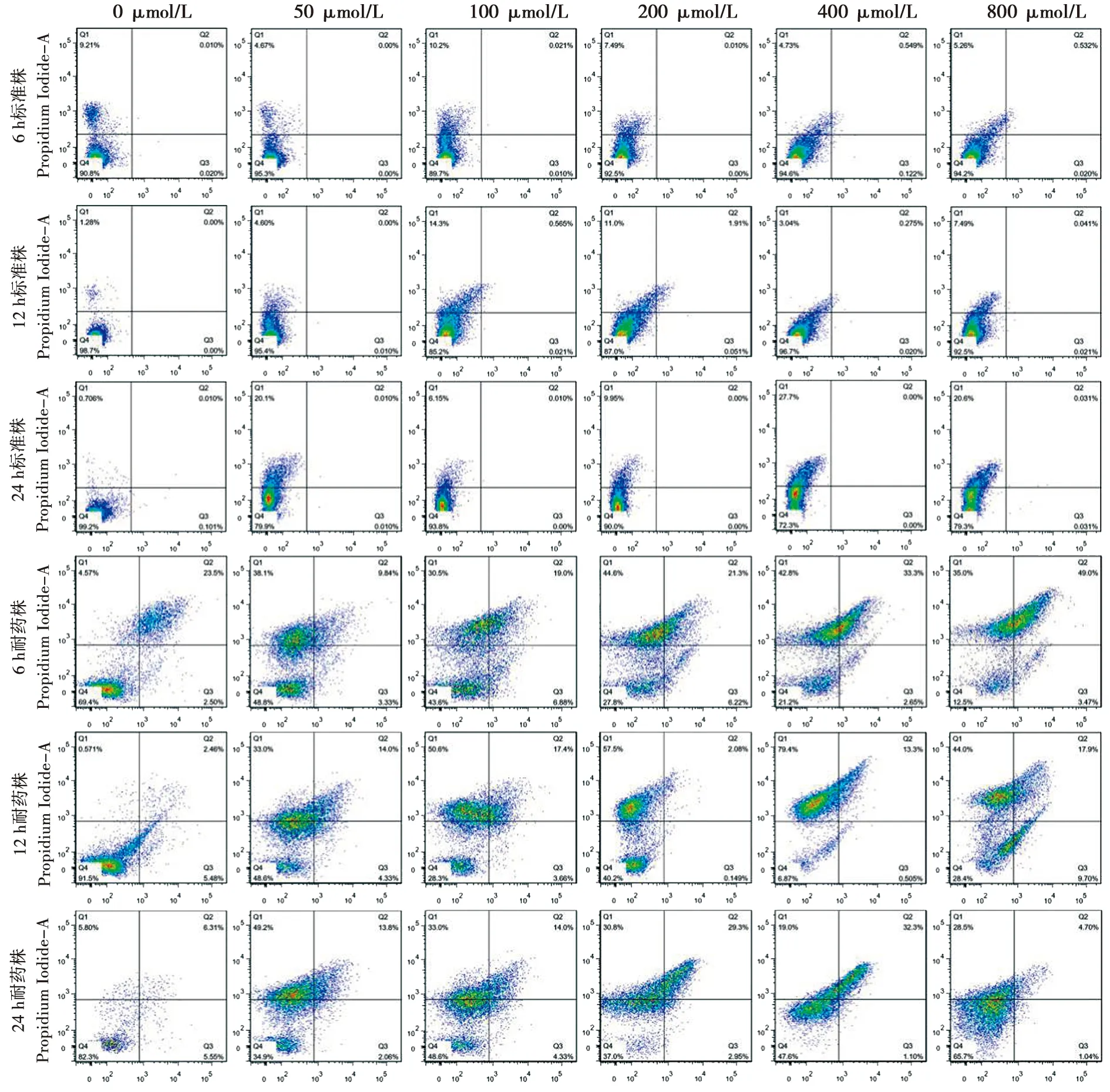
Q1、Q2、Q3和Q4分別表示:壞死細胞、早期凋亡細胞、晚期凋亡細胞和正常細胞
圖2流式細胞法檢測標準株及耐藥株生物被膜細胞的凋亡和壞死
Fig.2Test of cell apoptosis and necrosis ofC.albicansbiofilms of resistant and standard strains stained by flow cytometry
2.2.1 法尼醇對白念珠菌耐藥株和標準株生物膜凋亡作用的比較 法尼醇對白念珠菌耐藥株和標準株生物膜凋亡作用的比較:50~800 μmol/L法尼醇處理組及未處理對照組中,白念珠菌耐藥株凋亡率較標準株高(P<0.05),且在不同生物膜時相下具有一致性。結果見表2。
2.2.2 法尼醇對白念珠菌耐藥株和標準株生物膜壞死作用的比較 法尼醇未處理對照組中,6 h和12 h生物膜時相下,白念珠菌耐藥株壞死率較標準株低(P<0.05);24 h生物膜時相下,白念珠菌耐藥株壞死率較標準株高(P=0.05)。在50~400 μmol/L法尼醇處理下,各時相下白念珠菌耐藥株的壞死率均高于標準株(P<0.05)。在800 μmol/L法尼醇處理下,6 h生物膜時相,耐藥株與標準株的壞死率無明顯差異(P>0.05);12 h生物膜時相,耐藥株壞死率較標準株低(P<0.001);24 h生物膜時相,耐藥株壞死率較標準株高(P<0.001)。結果見表3。
表2 法尼醇對白念株菌標準株和耐藥株生物膜凋亡作用的比較Tab.2 Comparison of the apoptosis effect of farnesol on C.albicans biofilms of resistant and standard strains

表3 法尼醇對白念珠菌標準株和耐藥株生物膜壞死作用的比較 Tab.3 Comparison of the necrotic effect of farnesol on C.albicans biofilms of resistant and standard strains %
2.2.3 法尼醇對白念珠菌耐藥株生物膜凋亡與壞死的作用 法尼醇對白念珠菌耐藥株生物膜凋亡的作用:耐藥株法尼醇處理組與其相應生物膜時相的未處理對照組的細胞凋亡率無顯著性差異(P>0.05)。耐藥株法尼醇處理組與未處理對照組的細胞凋亡率在各生物膜時相具有一致性,且各時相下耐藥株生物膜凋亡率無顯著性差異(P>0.05)。結果見圖3。
法尼醇對白念珠菌耐藥株生物膜壞死的作用:各時相耐藥株生物膜經法尼醇處理后的細胞壞死率顯著高于其相應生物膜時相的未處理對照組(P<0.001);6 h生物膜時相,隨著法尼醇處理濃度的增加,耐藥株生物膜細胞壞死率先上升后下降、再上升后下降,且在200 μmol/L濃度的法尼醇處理下,耐藥株生物膜細胞壞死率最高。12 h生物膜時相,隨著法尼醇處理濃度的增加,耐藥株生物膜細胞壞死率先上升后下降、再上升后下降,且在400 μmol/L濃度的法尼醇處理下,耐藥株生物膜細胞壞死率最高。24 h生物膜時相,隨著法尼醇處理濃度的增加,耐藥株生物膜細胞壞死率先上升后下降、再上升,且在50 μmol/L濃度的法尼醇處理下,耐藥株生物膜細胞壞死率最高。
在100 μmol/L、400~800 μmol/L濃度的法尼醇處理下,12 h耐藥株生物膜細胞壞死率高于6 h生物膜(P<0.05);但在200~800 μmol/L濃度法尼醇處理下,24 h耐藥株生物膜細胞壞死率低于6 h生物膜(P<0.05)。結果見圖3。

各時相下法尼醇處理組白念珠菌耐藥株生物膜細胞壞死率高于相應時相的未處理對照組(P<0.001),法尼醇對白念株菌耐藥株生物膜的凋亡作用不明顯(P>0.05)
圖3法尼醇對白念珠菌耐藥株生物膜細胞凋亡和壞死的作用
Fig.3The effect of farnesol on necrosis and apoptosis ofC.albicansbiofilms of resistant strains
2.2.4 法尼醇對白念珠菌標準株生物膜凋亡與壞死的作用 法尼醇對白念珠菌標準株生物膜凋亡的作用:標準株的法尼醇處理組與其相應生物膜時相的未處理對照組的細胞凋亡率無顯著性差異(P>0.05)。標準株法尼醇處理組與未處理對照組的細胞凋亡率在各生物膜時相具有一致性,且各時相下標準株生物膜凋亡率無顯著性差異(P>0.05)。
法尼醇對白念珠菌標準株生物膜壞死的作用:各個時相標準株生物膜經法尼醇處理后的細胞壞死率高于其相應生物膜時相的未處理對照組(P<0.029)。6 h和24 h生物膜時相,隨著法尼醇處理濃度的增加,標準株生物膜細胞壞死率先上升后下降再上升,且在800 μmol/L濃度的法尼醇處理下,標準株生物膜細胞壞死率最高。12 h生物膜時相,標準株生物膜細胞壞死率與法尼醇處理濃度呈正相關,且在800 μmol/L濃度的法尼醇處理下,標準株生物膜細胞壞死率最高。
在400~800 μmol/L濃度的法尼醇處理下,12 h標準株生物膜細胞壞死率高于6 h生物膜(P<0.05);但在100~800 μmol/L濃度法尼醇處理下,24 h標準株生物膜細胞壞死率低于6 h生物膜(P<0.009)。見圖4。

各時相下法尼醇處理組白念珠菌標準株生物膜細胞壞死率高于相應時相的未處理對照組(P<0.05),法尼醇對白念株菌標準株生物膜的凋亡作用不明顯(P>0.05)
圖4法尼醇對白念珠菌標準株生物膜細胞凋亡和壞死的作用
Fig.4The effect of farnesol on necrosis and apoptosis ofC.albicansbiofilms of standard strains
2.3 q-PCR分析法尼醇對白念珠菌生物膜CDR1和MCA1基因表達的改變
法尼醇對白念株菌標準株和耐藥株生物膜CDR1基因表達的改變:標準株6 h生物膜法尼醇處理組CDR1表達高于未處理對照組(P<0.05),與法尼醇濃度呈正相關。在50~800 μmol/L濃度法尼醇處理下的耐藥株6 h生物膜,其CDR1表達高于未處理對照組(P<0.05),且在200 μmol/L濃度的法尼醇處理下,CDR1表達最高。在50~800 μmol/L濃度法尼醇處理下的耐藥株12 h生物膜,其CDR1表達高于未處理對照組(P<0.05),且在400 μmol/L濃度的法尼醇處理下,CDR1表達最高。在200~400 μmol/L法尼醇處理下的耐藥株24 h生物膜,其CDR1表達高于未處理對照組(P<0.05),且在400 μmol/L濃度的法尼醇處理下,CDR1表達最高。標準株12 h和24 h生物膜法尼醇處理組與其相應生物膜時相的未處理對照組CDR1表達無明顯差異(P>0.05)。見圖5。
法尼醇對白念株菌標準株和耐藥株生物膜MCA1基因表達的改變:在100 μmol/L、400~800 μmol/L濃度法尼醇處理下的標準株6 h生物膜,其MCA1表達高于未處理對照組(P<0.05),且在800 μmol/L濃度的法尼醇處理下,MCA1表達最高。在400 μmol/L濃度法尼醇處理下的標準株12 h生物膜,其MCA1表達高于未處理對照組(P=0.023)。在50~800 μmol/L濃度法尼醇處理下的耐藥株6 h生物膜,其MCA1表達高于未處理對照組(P<0.05),且在200 μmol/L濃度的法尼醇處理下,MCA1表達最高。標準株24 h生物膜、耐藥株12 h和24 h生物膜法尼醇處理組與其相應生物膜時相的未處理對照組MCA1表達無明顯差異(P>0.05)。見圖5。
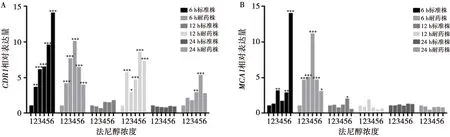
1、2、3、4、5、6分別是:0 μmol/L、50 μmol/L、100 μmol/L、200 μmol/L、400 μmol/L、800 μmol/L
圖5法尼醇對白念珠菌生物被膜CDR1基因(A)和MCA1基因(B)表達的影響
Fig.5Comparison of the effect of farnesol on the expression ofCDR1gene (A) andMCA1gene (B) ofC.albicansbiofilms
3 討論
細胞死亡根據形態學特征和機制的不同,分為細胞凋亡和細胞壞死。細胞凋亡是由基因控制的有秩序、有控制、有預定程序的死亡過程,它涉及部分基因的激活、表達和調控。細胞壞死是一種由化學、物理或生物等因素造成的傷害所引起的細胞死亡情況。近年來,研究發現壞死亦可被調節,通常在凋亡被抑制的情況下發生,且同細胞凋亡一樣受細胞內信號因子的調節[12]。法尼醇作為一種密度感應分子,對微生物的繁殖生長有調控作用[13-14]。多個研究發現法尼醇可以導致白念珠菌細胞死亡[5,8],但缺乏法尼醇對白念珠菌生物膜死亡作用機制的相關研究。本實驗對不同濃度法尼醇誘導不同時相耐藥株和標準株生物膜死亡的作用在凋亡和壞死方面進行初步探討。
Hwang等[15]證實法尼醇可以在浮游狀態白念珠菌細胞膜上累積,破壞細胞膜的完整性。本實驗通過對白念珠菌生物膜PI染色觀察其作用。PI是一種核酸染料,無法透過正常的細胞膜。正常細胞及凋亡細胞因細胞膜完整性未被破壞,細胞核無法被著色,而壞死細胞的細胞膜破裂,染料透過細胞膜對細胞內核酸進行染色,熒光顯微鏡下觀察壞死細胞為紅色。Maurya等[16]使用PI試劑對白念珠菌細胞進行形態學研究時,壞死細胞熒光顯微鏡下呈現紅色熒光。本實驗觀察到生物膜中PI染色后紅色熒光標記的白念珠菌細胞,提示法尼醇作用下白念珠菌細胞膜完整性受到破壞。進而通過形態學研究及流式分析,發現法尼醇作用于白念珠菌生物膜后,凋亡細胞少,壞死細胞表現顯著,且與法尼醇未處理對照組相比,細胞凋亡無明顯變化,細胞壞死明顯增加。提示法尼醇促進白念珠菌生物膜細胞死亡的作用主要為促進壞死的作用。有研究也發現高濃度法尼醇可導致白念珠菌生物膜壞死[5],與本研究一致。
同時,本研究發現同一生物膜時相、在同一濃度法尼醇處理下,耐藥株生物膜細胞的壞死率高于標準株,提示法尼醇對白念珠菌耐藥株生物膜具有明顯的促進壞死的作用,提示法尼醇可用于對抗耐藥株的耐藥性。康健等也報道法尼醇促進浮游狀態白念珠菌耐藥株壞死[6],與本研究對生物膜白念珠菌研究結果一致。
研究發現法尼醇對白念珠菌生物膜的抑制作用與法尼醇濃度有關[17]。本組實驗形態學研究發現隨著法尼醇處理濃度的升高,白念珠菌生物膜中細胞密度降低,菌絲長度減少,壞死細胞數目增加。流式分析顯示耐藥株和標準株細胞壞死率與法尼醇濃度相關。此外,耐藥株和標準株生物膜在不同時相下雖然存在不同的最佳抑菌濃度,但各濃度法尼醇處理組的壞死率均高于相應時相的未處理對照組,提示法尼醇對生物膜的抑制作用可能與其誘導壞死作用相關。
法尼醇對白念珠菌生物膜的抑制作用與生物膜的時相有關[18]。本研究選取三個時間位點進行研究,分別為6 h(生物膜早期)、12 h(生物膜中期)和24 h(生物膜成熟期)。不同生物膜時相,白念珠菌的菌絲相和酵母相所占比例不同。隨著生物膜的成熟,白念珠菌菌絲相增多,而酵母相減少。在6 h早期生物膜,白念珠以酵母相為主,但代謝較低。在12 h中期生物膜,白念珠菌從酵母相向菌絲相轉化,代謝增高。在24 h生物膜,白念珠菌以維持菌絲相為主,代謝隨之降低。Nagy等的菌絲生長動力學實驗分析顯示,法尼醇既抑制菌絲生長率,又延長菌絲長出的時間[19]。法尼醇能通過抑制Cup9的降解控制白念珠菌菌絲形成的啟動[20]。對不同時相生物膜死亡情況進行比較,發現相同濃度法尼醇作用于耐藥株及標準株生物膜,法尼醇對12 h生物膜的促進壞死作用最強,這可能與12 h生物膜代謝增高以及酵母相細胞所占總細胞的比率大,導致其對法尼醇的抑制作用最敏感相關。法尼醇促進24 h生物膜的壞死作用最弱,推測與生物膜進入成熟階段后代謝降低、酵母細胞較少、菌絲多且長、細胞外基質較多、基質包裹酵母細胞和菌絲形成復雜的三維結構等因素相關。這些研究結果說明生物膜時相影響法尼醇促進耐藥株及標準株生物膜細胞壞死的作用。
CDR1基因編碼的ABC轉運蛋白可將小分子物質進行跨膜轉運,導致細胞體內藥物濃度降低,造成細胞耐藥[7]。研究發現法尼醇對浮游狀態白念珠菌CDR1高表達菌株具有較高的殺滅作用,推測CDR1通過外排法尼醇-谷胱甘肽復合物,消耗白念珠菌內源性谷胱甘肽,使其抗氧化能力降低最終導致死亡[5]。本研究中在法尼醇作用下,早期白念珠菌標準株生物膜CDR1基因表達增加,耐藥株生物膜3個時相的CDR1基因表達均上升。提示法尼醇處理下,白念珠菌生物膜細胞的CDR1基因表達增高,說明法尼醇對耐藥株生物膜及標準株早期生物膜細胞壞死作用與其對高表達CDR1基因細胞壞死的促進作用一致。
MCA1編碼的Caspase酶,參與細胞的生長、分化與凋亡調節,其過度表達會導致細胞體內過氧化物集聚,殺傷細胞[9]。本研究中,法尼醇促進早期白念珠菌標準株和耐藥株生物膜MCA1基因的表達,MCA1基因的表達在中期和晚期生物膜中無顯著性改變。研究提示,法尼醇促進白念珠菌耐藥株及標準株晚期生物膜死亡的作用可能與MCA1基因無明顯相關性。推測法尼醇對白念珠菌中期及晚期生物膜的作用是一種Caspase酶非依賴性的過程,可能不會促進生物膜的凋亡,其具體作用機制有待進一步探討。
以往對法尼醇抑制白念生物膜的研究主要為研究法尼醇影響細胞形態轉變、抑制菌絲的生長及成熟。對于耐藥白念珠菌的研究集中在通過藥物抑制耐藥基因及蛋白的表達,減少治療藥物的外排從而達到治療感染的作用。未見不同濃度法尼醇對不同時相白念珠菌耐藥株及標準株生物膜死亡的作用及其比較研究。本實驗對不同濃度法尼醇誘導耐藥株和標準株生物膜死亡的作用在凋亡和壞死方面進行初步探討,初步證實法尼醇對白念生物膜有促進壞死的作用。本研究為如何利用法尼醇促進白念珠菌生物膜壞死的作用,以治療白念珠菌生物膜感染,特別是耐藥生物膜的感染提供理論思路。
4 結 論
法尼醇對白念珠菌生物膜的作用以促進壞死為主,且對耐藥株作用較對標準株顯著。法尼醇的這種作用與生物膜時相相關,CDR1和MCA1與耐藥株生物膜壞死相關。
猜你喜歡
城市道橋與防洪(2022年4期)2022-07-01 06:04:12
保健醫苑(2022年5期)2022-06-10 07:46:38
現代臨床醫學(2022年3期)2022-06-06 07:59:40
昆明醫科大學學報(2022年1期)2022-02-28 07:43:40
科學大眾(2020年12期)2020-08-13 03:22:22
當代陜西(2019年8期)2019-05-09 02:22:48
動漫星空(興趣百科)(2019年3期)2019-03-07 07:23:10
家庭影院技術(2018年4期)2018-05-09 07:07:52
專用汽車(2016年4期)2016-03-01 04:13:43
質量與標準化(2015年9期)2015-12-31 11:41:40
