1-磷酸鞘氨醇抑制TGF-β誘導的人臍靜脈內皮細胞間充質轉化*
2020-06-09 02:41:50湯石林彭良善趙正亮譚一清王橋生
中國病理生理雜志 2020年5期
關鍵詞:功能
湯石林, 彭良善, 趙正亮, 譚一清, 顏 波, 王橋生
(南華大學附屬第一醫院重癥醫學科,湖南衡陽421001)
心肌纖維化(myocardial fibrosis,MF)是各種心肌損害發展至終末階段的共同病理特征,可誘發心律失常,心肌炎和心力衰竭,是導致心源性猝死的潛在危險因素。因此,改善或逆轉MF是治療心血管疾病的重要策略[1]。MF是肌成纖維細胞過度增殖,膠原過度沉積及異常分布所引起的病理過程。以往研究認為,肌成纖維細胞主要來源于局部組織中的成纖維細胞、上皮細胞等。新近研究表明,血管內皮細胞也是肌成纖維細胞的重要來源之一[2-3]。相關研究表明,內皮細胞可在多種因素刺激作用下逐漸失去其形態和功能,并獲得間充質或肌成纖維細胞表型及其生物學特性,即發生內皮-間充質轉化(endothelial-to-mesenchymal transition,End-MT)[4]。因此,深入探究內皮細胞的轉化過程可為MF的治療提供參考依據。
1-磷酸鞘氨醇(sphingosine 1-phosphate,S1P)屬于鞘磷脂代謝產物,能夠通過S1P受體1~5(S1Preceptor 1~5,S1PR1~5)激活下游信號通路,調節細胞生存、分化、增殖、遷移等生物學功能,或以胞內第二信使的方式發揮其調節作用。相關研究證實,S1P可通過S1PR1、S1PR3等參與調節內皮細胞增殖、遷移、分化等過程[5-6]。此外,S1P能夠顯著增加間充質干細胞的抗纖維化能力[7],說明S1P可能在抑制心肌纖維化過程中發揮重要的調節作用。但是,S1P抑制心肌纖維化的作用是否依賴S1PR以及End-MT是否在其中發揮功能,有待進一步探討。
Smads家族蛋白作為轉化生長因子β(transforming growth factor-β,TGF-β)受體下游重要的信號轉導因子,在TGF-β誘導的End-MT經典模型中發揮重要的調節作用[8]。研究表明,在調節CD4+T細胞分化過程中,S1PR1能夠抑制Smad3信號通路,進而拮抗TGF-β功能[9]。然而,S1P介導的S1PR1信號途徑是否可通過調節Smad3信號通路影響TGF-β誘導的End-MT尚不明確。本研究擬通過人臍靜脈內皮細胞(human umbilical vein endothelial cells,HUVECs)觀察S1P及其受體在TGF-β誘導的End-MT中的作用,并對其機制進行初步探討,以期為抑制心肌纖維化及其誘發的心功能不全提供參考資料。
材料和方法
1 材料與試劑
HUVECs購自ATCC。DMEM高糖培養液、胎牛血清、青霉素和鏈霉素購自Gibco;小鼠抗人CD31單克隆抗體、小鼠抗人VE-cadherin單克隆抗體和重組人S1P蛋白購自Abcam;小鼠抗人α-平滑肌肌動蛋白(α-smooth muscle actin,α-SMA)單克隆抗體和兔抗人成纖維細胞特異性蛋白1(fibroblast-specific protein 1,FSP1)多克隆抗體購自 Sigma;兔抗人Smad3單克隆抗體和兔抗人p-Smad3抗體購自Thermo Fisher;小鼠抗人GAPDH單克隆抗體和辣根過氧化物酶標記的羊抗小鼠Ⅱ抗購自武漢博士德公司;其它試劑均為進口或國產分析純。
2 方法
2.1 細胞培養 HUVECs用含10%胎牛血清、1×105U/L青霉素和100 mg/L鏈霉素的DMEM培養液,在37℃、5%CO2培養箱中靜置培養。倒置顯微鏡觀察細胞狀態,當細胞融合度超過80%時,采用0.25%胰蛋白酶消化細胞、傳代培養。根據實驗要求,分為TGF-β(10 μg/L[10])組、TGF-β+S1P(200 nmol/L[11])組和 TGF-β+S1P+VPC23019(10 μmol/L[12];S1PR1抑制劑)組。
2.2 Western blot實驗 收集細胞裂解液,并用BCA蛋白定量試劑盒進行蛋白質定量;SDS-PAGE上樣緩沖液(5×)矯正蛋白濃度,煮沸 5~8 min,冷卻,-20℃保存備用。采用SDS-PAGE(80 V,30 min;120 V,90 min)分離蛋白;轉膜(200 mA,2 h);TBST封閉液(含5%脫脂奶粉)封閉2 h;按比例稀釋CD31、VE-cadherin、α -SMA、FSP1、GAPDH 和 p-Smad3抗體,4℃孵育過夜;TBST洗3次,每次10 min;辣根過氧化物酶標記的Ⅱ抗室溫孵育2 h,TBST洗3次,每次10 min;經ECL顯影劑處理后,采用自動化學發光系統拍照分析。
2.3 免疫熒光染色 24孔板培養細胞,加入處理因素后,PBS浸洗3次,每次3 min;4%的多聚甲醛固定細胞30 min,PBS浸洗3次,每次3 min;0.5%Triton X-100室溫通透30 min;PBS浸洗3次,每次3 min;正常山羊血清工作液室溫封閉30 min;棄封閉液,I抗工作液4℃孵育過夜;PBS浸洗3次,每次3 min;熒光Ⅱ抗37℃避光孵育1 h,PBS浸洗3次,每次3 min;倒置熒光相差顯微鏡拍照觀察。
3 統計學處理
以GraphPad Prism 6.0軟件進行統計分析。所有實驗數據均采用均數±標準差(mean±SD)表示,組間比較采用方差分析或t檢驗,以P<0.05認為差異有統計學意義。
結 果
1 S1P抑制TGF-β誘導的End-MT
Western blot檢測End-MT相關標志物的表達,結果顯示,與TGF-β組相比,S1P+TGF-β組內皮細胞標志物CD31和VE-cadherin表達水平顯著增加(P<0.05),間充質細胞標志物α-SMA和FSP1表達水平則顯著降低(P<0.05),見圖1。
2 S1P通過S1PR1降低TGF-β誘導的End-MT
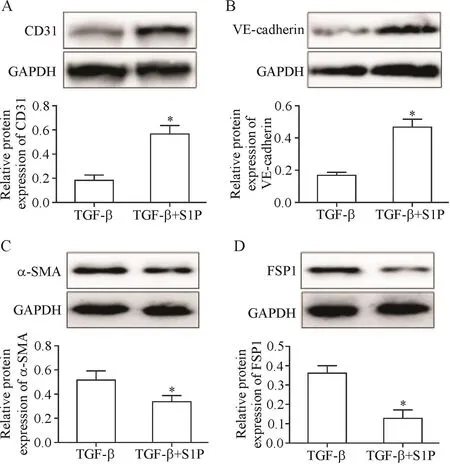
Figure 1.S1Pinhibited TGF-β-induced End-MT.The expression of CD31(A),VE-cadherin(B),α-SMA(C)and FSP1(D)was determined by Western bolt.Mean±SD.n=3.*P<0.05 vs TGF-β group.圖1 S1P抑制TGF-β誘導的End-MT
為了進一步證實S1PR1在S1P抑制TGF-β誘導的End-MT中的作用,我們采用S1PR1抑制劑VPC23019處理HUVECs。結果顯示,與S1P+TGF-β組相比,S1P+TGF-β+VPC23019組內皮細胞標志物CD31和VE-cadherin的表達顯著降低(P<0.05),而間充質細胞標志物α-SMA和FSP1的表達顯著增加(P<0.05),見圖2。
3 S1PR1抑制劑VPC23019下調S1P作用下HUVECs中Smad3活性
與TGF-β處理組相比,S1P顯著降低了HUVECs中Smad3的磷酸化水平,并抑制其核轉位(P<0.05);而添加S1PR1抑制劑VPC23019處理后,S1P降低Smad3磷酸化及核轉位的作用被逆轉(P<0.05),見圖3。
討 論
End-MT能夠進一步增加間充質或肌成纖維細胞,尋找調控End-MT的策略對有效控制和減輕心肌纖維化及其誘發的各類心血管疾病具有重要意義[13-14]。S1P作為細胞鞘脂類物質的代謝活性物質,其相關信號通路在調節心肌纖維化進程、內皮細胞生存及功能過程中發揮重要作用。鑒于S1P在心肌及內皮細胞中的功能,我們推測S1P可能通過介導內皮細胞表型轉化參與心肌功能的調節。本實驗數據表明,S1P具有抑制TGF-β誘導的HUVECs的End-MT功能,說明S1P抑制心肌纖維化的功能可能部分依賴于其對End-MT進程的抑制作用。
S1P作為胞內第二信使,可直接特異性結合組蛋白脫乙酰酶 1/2(histone deacetylases 1/2,HDAC1/HDAC2),抑制其活性,進而增強組蛋白H3乙酰化程度,促進相關基因的轉錄[15]。HDAC1被證實在上皮-間 充 質 轉 化(epithelial-to-mesenchymal transition,EMT)過程中發揮重要調節作用[16],而 End-MT 是EMT的一種特殊類型,提示S1P介導胞內HDAC1的變化可能參與了其對End-MT的調節。S1PR1作為S1P在內皮細胞中的受體之一,是介導S1P發揮功能的關鍵蛋白[17]。相關研究表明,S1P受體激動劑FTY720能夠通過抗炎和抗凋亡途徑降低異位心臟移植后心肌纖維化水平[18]。本研究也表明,S1PR1抑制劑VPC23019能夠顯著逆轉S1P對TGF-β誘導的End-MT的作用,說明S1P抑制TGF-β誘導的End-MT依賴S1PR1。
另外,我們利用Western blot及免疫熒光染色證實,S1P可能通過S1PR1途徑降低Smad3的磷酸化水平和核轉位,進而影響TGF-β誘導的End-MT進程。然而,在本研究中,我們僅觀察了S1P-S1PR1軸對Smad3信號的影響,而S1P對TGF-β介導的End-MT的具體調節機制是否依賴于Smad途徑,以及是否存在非Smad依賴信號途徑尚不清楚。
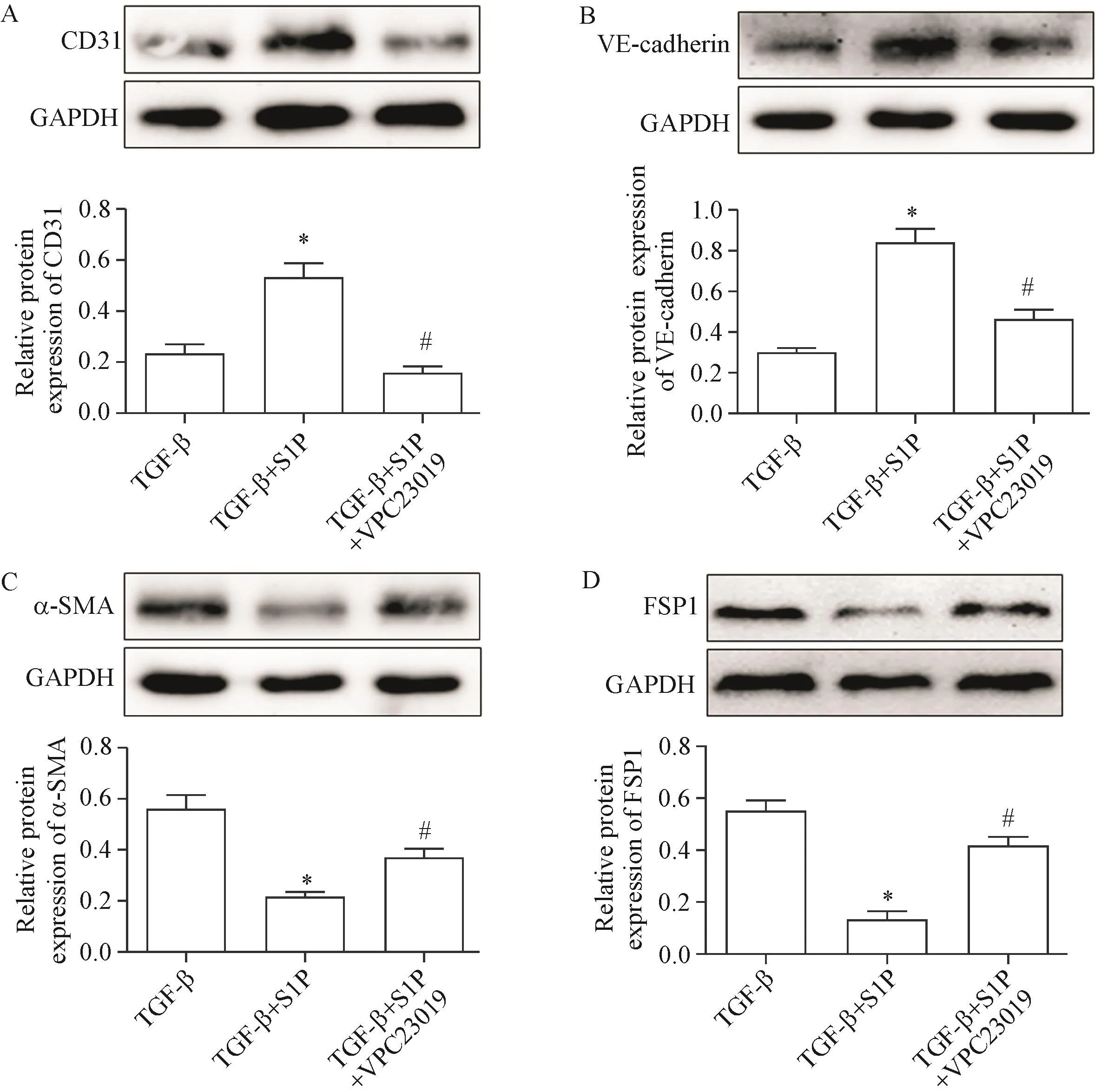
Figure 2.S1Pinhibited TGF-β-induced End-MTthrough S1PR1.The expression of CD31(A),VE-cadherin(B),α-SMA(C)and FSP1(D)was detected by Western bolt.Mean±SD.n=3.*P<0.05 vs TGF-β group;#P<0.05 vs TGF-β+S1Pgroup.圖2 S1P經S1PR1途徑抑制TGF-β誘導的End-MT
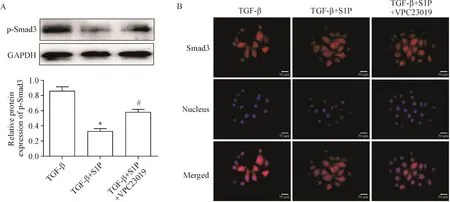
Figure 3.The effect of S1Pand S1PR1 on Smad3 signaling under the action of TGF-β.A:the phosphorylation level of Smad3 was detected by Western blot;B:the nuclear translocation of Smad3 was detected by immunofluorescence staining.Mean±SD.n=3.*P<0.05 vs TGF-β group;#P<0.05 vs TGF-β+S1Pgroup.圖3 S1P及S1PR1對TGF-β作用下Smad3信號的影響
綜上所述,本研究在體外證實S1P-S1PR1可抑制TGF-β誘導的HUVECs的End-MT,其機制可能與Smad3磷酸化及核轉位有關。
猜你喜歡
鐘表(2023年5期)2023-10-27 04:20:44
中華詩詞(2022年6期)2022-12-31 06:41:24
當代陜西(2021年21期)2022-01-19 02:00:26
中學生數理化(高中版.高考數學)(2020年1期)2020-02-20 13:23:44
經濟技術協作信息(2018年11期)2019-01-14 03:07:20
中國科技論壇(2017年7期)2017-07-25 08:49:53
制造技術與機床(2017年3期)2017-06-23 08:11:33
媽媽寶寶(2017年2期)2017-02-21 01:21:24
國際漢語學報(2016年1期)2017-01-20 08:21:20
中國中醫藥現代遠程教育(2014年22期)2014-03-01 04:32:55
