水稻OsDTH13 基因編輯突變體的創(chuàng)制與分析
2020-06-10 02:13:44石雨鷺靳亞軍欒維江
關(guān)鍵詞:水稻
石雨鷺,鄭 瑞,靳亞軍,欒維江
(1.天津師范大學(xué)生命科學(xué)學(xué)院,天津300387;2.天津師范大學(xué)天津市動(dòng)植物抗性重點(diǎn)實(shí)驗(yàn)室,天津300387)
水稻成花素是抽穗開花的促進(jìn)因子,是一種在葉片中形成并轉(zhuǎn)運(yùn)到頂端分生組織起作用的小分子蛋白.該蛋白因具有磷脂酰乙醇胺(phosphatidylethanolamine-binding protein,PEBP)的結(jié)合結(jié)構(gòu)域,屬于PEBP蛋白家族.PEBP 基因家族在進(jìn)化上十分保守,廣泛存在于細(xì)菌、動(dòng)物和植物中[1].就植物中的PEBP 編碼基因而言,根據(jù)編碼氨基酸序列的同源性,又分為FT-like、TFL-like 和MFT-like 3個(gè)亞家族[2].FT-like 和TFL-like亞家族基因可能是由MFT-like 亞家族基因進(jìn)化而來[1].
植物成花素基因最早在擬南芥中得到鑒定,擬南芥基因組包含8個(gè)PEBP 編碼基因[3],其中已知的FT是成花素,可以促進(jìn)擬南芥的開花;TFL1 是擬南芥的反成花素,抑制其開花[4].油菜基因組包含26個(gè)PEBP 基因[5].大豆基因組包含23個(gè)PEBP 基因[6],其中GmFT2a和GmFT5a 可以調(diào)控開花時(shí)間,促進(jìn)大豆的開花[7].水稻基因組包含19個(gè)PEBP 基因,其中,13個(gè)為FTLike 成員,已明確功能的Hd3a 和RFT1 分別是水稻在短日照(SD)和長日照(LD)條件下的成花素基因[8],可以在SD 和LD 條件下促進(jìn)水稻的成花,其余成員的功能未知;4個(gè)屬于TFL-like 成員,可能具有反成花素作用;2個(gè)屬于MFT-like 成員,即OsMFT1 和OsMFT2,它們?cè)诜N子發(fā)育和萌發(fā)過程中呈現(xiàn)出較高的表達(dá)水平[9],過量表達(dá)OsMFT1 可推遲開花,并且改變穗型,2個(gè)成員的詳細(xì)功能還不明確.
CRISPR/Cas9 系統(tǒng)是近幾年發(fā)展的一種準(zhǔn)確、高效、應(yīng)用廣泛的基因編輯技術(shù),與鋅指核酸酶技術(shù)(ZFNs)、轉(zhuǎn)錄激活樣效應(yīng)因子核酸酶技術(shù)(TALENs)相比優(yōu)勢(shì)突出[10].CRISPR/Cas9 系統(tǒng)作用機(jī)理簡單,僅需通過1個(gè)sgRNA(single guide RNA) 和核酸酶(Cas9)就可以實(shí)現(xiàn)基因的定點(diǎn)編輯[11-12].在gRNA 的引導(dǎo)下識(shí)別將要切割的靶序列,由Cas9 蛋白在靶點(diǎn)附近切割DNA 雙鏈產(chǎn)生染色體雙鏈斷裂,隨后通過非同源末端連接進(jìn)行修復(fù),由于修復(fù)的不準(zhǔn)確導(dǎo)致堿基插入或缺失,從而造成基因突變[13].目前,已有研究表明,CRISPR/Cas9 系統(tǒng)在擬南芥、小麥、水稻、玉米、煙草以及苔蘚植物中得到了應(yīng)用[14].
本研究利用CRISPR/Cas9 技術(shù)對(duì)水稻的OsDTH13基因進(jìn)行定點(diǎn)編輯,并對(duì)獲得的編輯植株進(jìn)行測(cè)序分析,明確其編輯突變類型,得到了編輯突變體,為后續(xù)OsDTH13 基因的功能研究提供實(shí)驗(yàn)基礎(chǔ).
1 材料與方法
1.1 實(shí)驗(yàn)材料
粳稻品種中花11(Oryza sativa L.ssp. Japonica cv.Zhonghua11)及其轉(zhuǎn)基因植株均種植于天津師范大學(xué)實(shí)驗(yàn)田,田間保持常規(guī)管理.
1.2 靶位點(diǎn)選擇及引物設(shè)計(jì)
利用E-CRISPR(http://www.e-crisp.org)在線設(shè)計(jì)平臺(tái)對(duì)OsDTH13 基因的編碼區(qū)序列進(jìn)行分析,選用2個(gè)GN19NGG 特征的序列作為編輯靶位點(diǎn)(分別為TG1 和TG2),以提高編輯效率.這2個(gè)靶點(diǎn)在目的基因的第一個(gè)外顯子上,GC 含量分別為60%和50%,發(fā)夾結(jié)構(gòu)較少. 將設(shè)計(jì)好的靶點(diǎn)在NCBI 數(shù)據(jù)庫中進(jìn)行BLAST 比對(duì),證實(shí)其具有良好的特異性,脫靶概率低.
1.3 CRISPR/Cas9 載體的構(gòu)建
分別取2個(gè)靶點(diǎn)TG1 和TG2 的上下游引物各10 μL,進(jìn)行退火反應(yīng).PCR 反應(yīng)程序?yàn)椋?5℃,3 min;以0.1 ℃/s 的速率降溫至22 ℃,獲得靶點(diǎn)特異片段.含有sgRNA 的OsU6SK 載體用限制性內(nèi)切酶BbsⅠ進(jìn)行過夜酶切,回收酶切產(chǎn)物,將其分別與2個(gè)靶位點(diǎn)特異片段于16 ℃過夜連接.連接產(chǎn)物熱激轉(zhuǎn)化后,挑取單克隆進(jìn)行菌液PCR 鑒定.鑒定正確的陽性質(zhì)粒與含有Cas9 的35S-Cas9-SK 載體及雙元pCAMBIA1300載體,分別用相應(yīng)的限制性內(nèi)切酶進(jìn)行酶切,回收后進(jìn)行連接,獲得可以轉(zhuǎn)化水稻的雙元重組載體.
1.4 轉(zhuǎn)基因植株的分子鑒定
將構(gòu)建成功的CRISPR/Cas9 重組載體轉(zhuǎn)入農(nóng)桿菌,利用農(nóng)桿菌介導(dǎo)的遺傳轉(zhuǎn)化法獲得轉(zhuǎn)基因植株.將轉(zhuǎn)基因植株移栽至實(shí)驗(yàn)田,待其生長到5 葉期,收集葉片,用CTAB 法提取水稻基因組DNA.以載體中的特異性潮霉素基因作為選擇標(biāo)記進(jìn)行PCR 分子鑒定.所用PCR 程序?yàn)椋?4 ℃預(yù)變性4 min;94 ℃變性30 s,56 ℃退火30 s,72 ℃延伸30 s,30個(gè)循環(huán);72 ℃終延伸7 min,4 ℃保存.
1.5 轉(zhuǎn)基因植株的突變類型分析
由于2個(gè)靶點(diǎn)TG1 和TG2 的位置相近,因此可以用同一對(duì)引物進(jìn)行檢測(cè). 依據(jù)OsDTH13 的基因組序列,在靶點(diǎn)序列上下游約250 bp 處設(shè)計(jì)靶點(diǎn)編輯檢測(cè)引物進(jìn)行PCR 擴(kuò)增,用瓊脂糖凝膠電泳進(jìn)行檢測(cè).將PCR 產(chǎn)物送至生工生物工程股份有限公司測(cè)序,測(cè)序結(jié)果與野生型序列比對(duì)分析.如測(cè)序結(jié)果為套峰,則利用在線平臺(tái)DSDecodeM(http://dsdecode.scgene.com/)對(duì)突變位點(diǎn)進(jìn)行解碼,并用凝膠電泳進(jìn)行驗(yàn)證,確定突變類型.
2 結(jié)果與分析
2.1 OsDTH13 序列分析及靶位點(diǎn)的選擇
OsDTH13 基因的DNA 序列長1576 bp,含有4個(gè)外顯子和3個(gè)內(nèi)含子,編碼序列總長558 bp,編碼186個(gè)氨基酸,其結(jié)構(gòu)如圖1 所示.
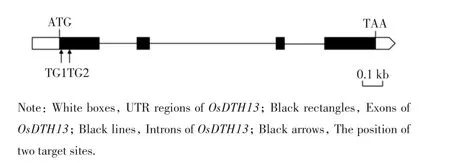
圖1 OsDTH13 基因結(jié)構(gòu)及靶點(diǎn)位置Fig.1 Structure of OsDTH13 gene and the position of target sites
依據(jù)OsDTH13 的基因結(jié)構(gòu)特點(diǎn)及靶點(diǎn)設(shè)計(jì)原則,在第一個(gè)外顯子內(nèi)篩選出了2個(gè)打靶效率高、特異性好的靶位點(diǎn)TG1 和TG2,分別位于ATG 后27~46 bp及48~67 bp.由于FT-like 家族成員的同源性較高,本研究通過BLAST 搜尋比對(duì),發(fā)現(xiàn)這2個(gè)靶點(diǎn)的脫靶率低,不會(huì)造成該家族與其序列相似成員的編輯.
2.2 CRISPR/Cas9 載體構(gòu)建
分別將2個(gè)靶點(diǎn)的正反向引物進(jìn)行退火反應(yīng),獲得了寡聚二聚體,連接至含有sgRNA 的OsU6SK 質(zhì)粒骨架上,獲得帶有靶點(diǎn)序列的sgRNA 融合載體(OsU6-SK-TG1 和OsU6-SK-TG2). 重組載體熱激轉(zhuǎn)化后,挑取單克隆進(jìn)行菌液PCR 擴(kuò)增,得到片段大小約為400 bp的目的條帶,如圖2(a)所示,與預(yù)期條帶大小相符.進(jìn)一步提取陽性克隆的質(zhì)粒進(jìn)行測(cè)序分析,結(jié)果表明2個(gè)目的靶點(diǎn)分別與OsU6SK 載體連接成功.然后將融合載體OsU6-SK-TG1、OsU6-SK-TG2、含有Cas9 的35S-Cas9-SK 空載體及pCAMBIA1300 雙元載體分別酶切,分別回收帶有靶點(diǎn)的sgRNA 片段、Cas9 片段及pCAMBIA1300 載體骨架進(jìn)行連接,轉(zhuǎn)化后獲得重組載體.將重組載體進(jìn)行酶切鑒定,結(jié)果顯示切出了預(yù)期大小的目的片段,如圖2(b)所示,表明帶有靶點(diǎn)的sgRNA 和Cas9 成功連入了pCAMBIA1300 雙元載體中,可以用于后續(xù)的轉(zhuǎn)基因.
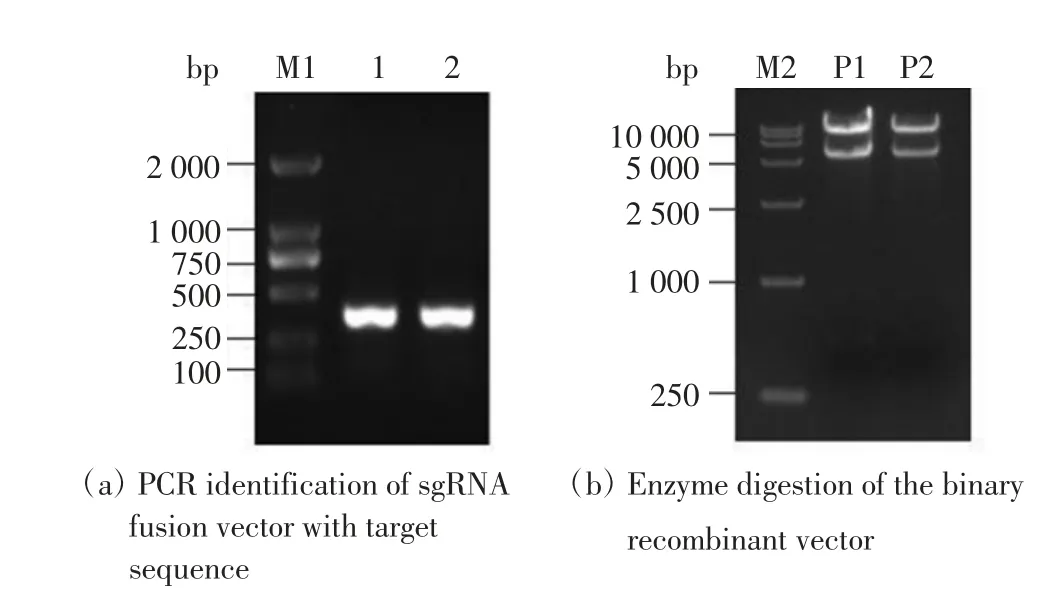
圖2 OsDTH13 基因的CRISPR/Cas9 載體構(gòu)建Fig.2 Construction of CRISPR/Cas9 vector for OsDTH13
2.3 轉(zhuǎn)基因植株的獲得和分子檢測(cè)
重組載體構(gòu)建成功后,通過農(nóng)桿菌介導(dǎo)的遺傳轉(zhuǎn)化法,經(jīng)愈傷組織的誘導(dǎo)、繼代、農(nóng)桿菌的侵染、抗性愈傷的篩選、分化、生根過程,最終獲得了20 株水稻轉(zhuǎn)基因植株.待轉(zhuǎn)基因植株生長健壯后,單株提取基因組DNA,用載體中特異的潮霉素基因進(jìn)行PCR 鑒定,結(jié)果如圖3 所示,有13 株為轉(zhuǎn)基因陽性植株.
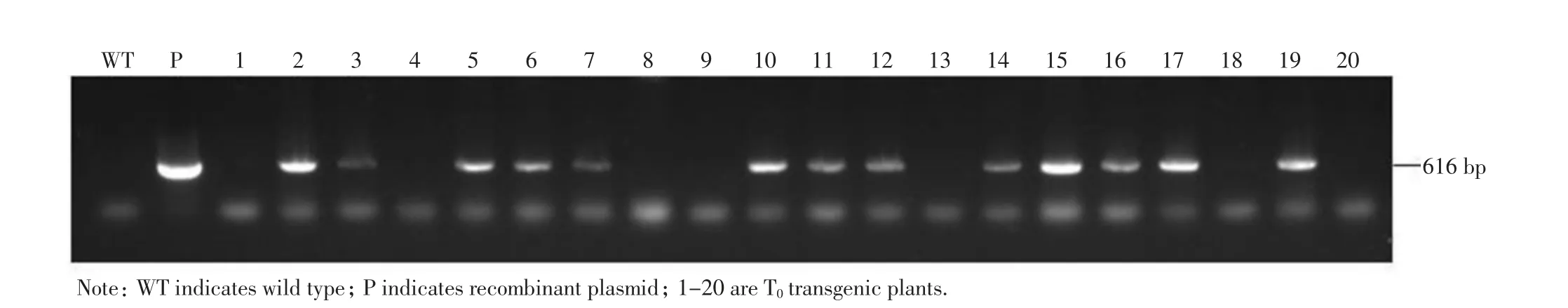
圖3 獲得的CRISPR/Cas9 轉(zhuǎn)基因植株的分子檢測(cè)Fig.3 Molecular detection of the obtained CRISPR/Cas9 transgenic plants
2.4 突變體的編輯類型分析
為了明確水稻轉(zhuǎn)基因植株中的目的基因是否被編輯,對(duì)獲得的全部T0代轉(zhuǎn)基因植株進(jìn)行了突變位點(diǎn)分析,采用CTAB 法提取基因組DNA,然后用包含靶點(diǎn)的特異性檢測(cè)引物進(jìn)行PCR 擴(kuò)增.擴(kuò)增產(chǎn)物經(jīng)測(cè)序分析,結(jié)果表明,13 株轉(zhuǎn)基因植株中有5 株發(fā)生了基因編輯(#1~#5).#1 編輯突變體是一個(gè)缺失的純合突變體,在第一靶點(diǎn)PAM 序列-8 處缺失24個(gè)堿基,用2%瓊脂糖凝膠電泳檢測(cè)PCR 擴(kuò)增產(chǎn)物,發(fā)現(xiàn)其產(chǎn)物比野生型擴(kuò)增產(chǎn)物小,如圖4(a)所示.#2 是一個(gè)雜合缺失突變體,其中一條同源染色體在第一靶點(diǎn)PAM序列-4 處缺失了1個(gè)堿基,另一條同源染色體在第一靶點(diǎn)PAM 序列-4 處缺失了9個(gè)堿基,因此測(cè)序結(jié)果出現(xiàn)套峰,如圖4(b)所示.#3 編輯突變體是一個(gè)純合突變體,產(chǎn)生了堿基缺失和替換2 種突變形式,PCR檢測(cè)結(jié)果表明,突變體擴(kuò)增出的條帶小于野生型,測(cè)序后發(fā)現(xiàn)與野生型相比,該突變體的2 條同源染色體在第一靶點(diǎn)PAM 序列-20 處發(fā)生堿基缺失,共缺失32 bp,在第二靶點(diǎn)PAM 序列-13 處由堿基C 替換為堿基G,如圖4(c)所示.#4 編輯突變體是一個(gè)含有插入和缺失的雙等位突變體,其中一條同源染色體在第一靶點(diǎn)PAM 序列-4 處缺失了1個(gè)堿基,另一條同源染色體序列在第一靶點(diǎn)PAM 序列-4 處插入了1個(gè)堿基,如圖4(d)所示. #5 編輯突變體是一個(gè)單堿基插入的純合突變體,在第二靶點(diǎn)PAM 序列-6 處插入了1個(gè)堿基,如圖4(e)所示.
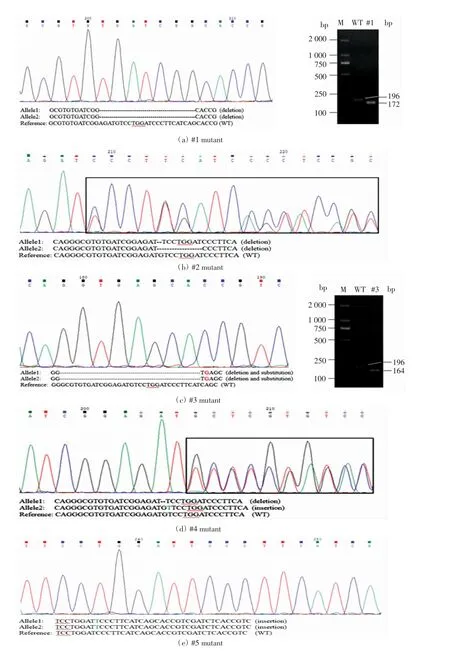
圖4 編輯突變體的測(cè)序分析及PCR 鑒定Fig.4 Sequencing analysis and PCR identification of editing mutants
2.5 突變植株的氨基酸序列分析
OsDTH13 基因的編碼序列長558 bp,編碼的蛋白質(zhì)含有186個(gè)氨基酸.利用DNAMAN 軟件對(duì)3個(gè)純合編輯突變體植株的氨基酸序列進(jìn)行分析,發(fā)現(xiàn)與野生型植株相比,突變體所產(chǎn)生的突變導(dǎo)致了部分氨基酸缺失或蛋白質(zhì)翻譯提前終止,如圖5 所示.
在氨基酸水平上,#1 突變體由于缺失24個(gè)堿基導(dǎo)致翻譯后缺失了8個(gè)氨基酸(圖5 中的#1-allele),并未造成移碼.#3 突變體在第一靶點(diǎn)PAM 序列-20 處缺失32 bp,在第二靶點(diǎn)PAM 序列-13 處由堿基C 替換為堿基G,導(dǎo)致移碼突變,從第10個(gè)氨基酸開始與野生型不同,并且終止于第19個(gè)氨基酸(圖5 中的#3-allele),最終產(chǎn)生一個(gè)僅有19個(gè)氨基酸的截短的多肽.#5 突變體是一個(gè)單堿基插入的純合突變體,插入造成移碼突變,并提前終止于第30個(gè)氨基酸處(圖5 中的#5-allele),產(chǎn)生一個(gè)30個(gè)氨基酸的截短的多肽.后續(xù)本課題組將利用這些突變體對(duì)OsDTH13 基因功能進(jìn)行研究.

圖5 野生型與3個(gè)突變體的氨基酸序列比對(duì)Fig.5 Comparison of amino acid sequences between wild type and three mutants
2.6 突變體植株的遺傳模式分析
將上述T0代純合編輯突變體獲得的種子進(jìn)行種植,得到T1代突變體植株.以#3 純合突變體為例,對(duì)所得的T1代突變體植株提取DNA,用設(shè)計(jì)的檢測(cè)引物進(jìn)行PCR 擴(kuò)增,擴(kuò)增產(chǎn)物通過2%瓊脂糖凝膠電泳進(jìn)行檢測(cè).結(jié)果顯示,所有T1代植株均表現(xiàn)為純合缺失突變,如圖6 所示.經(jīng)進(jìn)一步的測(cè)序驗(yàn)證,測(cè)序結(jié)果與膠圖相符,說明編輯突變體可以穩(wěn)定遺傳.
3 討論與結(jié)論
植物中PEBP 基因家族成員的主要功能是控制植物開花時(shí)間和生長發(fā)育.現(xiàn)已明確了擬南芥和水稻中幾個(gè)PEBP 基因?qū)﹂_花時(shí)間的調(diào)控作用.擬南芥中的FT 和TFL1 可以與14-3-3 蛋白及一個(gè)帶有堿性亮氨酸拉鏈蛋白結(jié)構(gòu)域的轉(zhuǎn)錄因子FD1 相互作用形成FAC 復(fù)合體(florigen activation complex),并隨之結(jié)合在AP1 等一些開花基因啟動(dòng)子區(qū)域,從而誘導(dǎo)植物開花[4,15].水稻中的成花素基因?yàn)镠d3a 和RFT1,是擬南芥開花激活子FT 基因在水稻中的同源基因.水稻中2個(gè)成花素的作用機(jī)理與擬南芥中的成花素作用機(jī)理保守,成花素首先在細(xì)胞質(zhì)中與受體14-3-3 互作,然后轉(zhuǎn)移到細(xì)胞核中與FD1 互作形成FAC 復(fù)合體,促進(jìn)下游OsMADS15 基因的表達(dá),從而促進(jìn)水稻的開花[15].Qin 等[16]在短柄草中鑒定出一個(gè)短日照特異表達(dá)的基因FTL9,F(xiàn)TL9 在短日照條件下促進(jìn)開花,而在長日照條件下抑制開花.目前,在大豆中發(fā)現(xiàn)了10個(gè)FT 的同源基因,其中GmFT2a 和GmFT5a 已被證實(shí)為主要的開花促進(jìn)因子,2個(gè)基因在不同光照條件下對(duì)開花的促進(jìn)作用存在差異,在短日照條件下GmFT2a 的開花促進(jìn)效應(yīng)比GmFT5a 更強(qiáng),而GmFT5a 在長日照條件下的開花促進(jìn)效應(yīng)更強(qiáng)[17].在進(jìn)化過程中,F(xiàn)T 基因的功能逐漸出現(xiàn)分化,一些基因不僅僅調(diào)控植物開花.最近研究表明,水稻的成花素Hd3a 還可以負(fù)向調(diào)控水稻的抗病性,hd3a 突變體提高了水稻對(duì)白葉枯病菌的抗性[18].基于此,研究水稻中PEBP 家族不同成員的詳細(xì)功能至關(guān)重要,對(duì)于全面理解該家族成員的功能具有重要意義.為揭示水稻中PEBP 家族成員的功能,本研究利用CRISPR-Cas9 技術(shù)創(chuàng)建了OsDTH13 基因的編輯突變體,最終得到了5 株突變體植株,其中2 株為單堿基插入,3 株為小片段的缺失.這些編輯突變體的獲得為OsDTH13 的生物學(xué)功能研究奠定了基礎(chǔ). 后續(xù)本課題組將利用獲得的能夠穩(wěn)定遺傳的純合突變體材料進(jìn)行表型觀察及基因功能研究,詳細(xì)揭示該基因在水稻中的生物學(xué)功能.
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學(xué)版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農(nóng)業(yè)(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農(nóng)業(yè)(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農(nóng)業(yè)(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
