小葉楊不同種源一年生扦插苗速生期表型性狀變異定量分析
2020-06-10 00:48:48葛會碩宋躍朋蘇雪輝張德強張曉宇
浙江林業科技 2020年1期
葛會碩,宋躍朋,蘇雪輝,張德強,張曉宇
(1.北京林業大學 理學院,北京 100083;2.北京林業大學 生物科學與技術學院,北京 100083;3.焦作市農林科學研究院,河南 焦作 454001)
小葉楊Populus simonii具有控制土壤荒漠化、減少土壤侵蝕和穩定沙丘的優勢[1]。同時,小葉楊還具有較高的經濟和藥用價值[2]。已有學者對小葉楊的生長特性,包括節間長度變量之間的關系和表型變異進行分析[3]。節間長度是幼苗生長的重要指標,節間長度變量間的關系對育種遺傳具有重要意義。2016 年,Yamamoto 等[4]基于圖像分析的機器學習方法探究了番茄Lycopersicon esculentum幼苗節間長度估計,能夠檢測到3 個番茄幼苗的時間序列圖像中358 個節點的72%。2017 年,李慧敏等[5]研究了玉蜀黍Zea mays穗下節間長度之間的關系,發現了兩個與產量和抗倒伏性相關的農藝性狀。2018 年,王健等[6]探討了普通小麥Triticum aestivum節間長度與株高的關系,結果表明矮稈小麥是通過縮短各節間長度來降低株高的。
在小葉楊的生長過程中,由于地理位置和生長環境的不同,小葉楊幼苗樣本可能具有不同的表型特征。沈德勝等[7]利用K-means 聚類方法提取了林木冠層孔隙度。考慮到K-means 聚類對初始聚類中心敏感和易陷入局部最優解,Wan 等[8]在K-means 聚類算法中引入了多種基于群體智能的優化算法,發現人工蜂群算法(ABC)的搜索能力與粒子群算法相當,前者收斂速度更快。所以采用基于人工蜂群算法的K-means 聚類(KABC)[9]可以有效防止陷入局部最優,進而達到準確分類的效果。
由于表型變異是復雜遺傳多樣性的外在表現,所以是遺傳多樣性的重要研究領域之一[10]。陳美熹等[11]以馬尾松Pinus massoniana58 年生采種母樹林為研究材料發現母樹間在球果表型及種子性狀均存在極顯著差異。2010 年衛尊征等[12]以東北和華北地區5 個種源的小葉楊為材料,分別對17 個表型及生理性狀進行了比較分析,發現小葉楊各性狀在種源間和種源內均存在廣泛的遺傳變異。2014 年,Du 等[3]研究了毛白楊Populus tomentosa的自然表型變異,發現各性狀在個體間存在著較大范圍的表型變異。
國內外學者對節間長度的研究主要在農作物方面,而林業上幼苗期的樹木節間長度直接反映了幼苗的生長狀況。此外,擴大種源范圍對研究小葉楊種源間和種源內遺傳變異具有重要意義。本文以小葉楊幼苗為研究材料,在2017 年4 月29 日至6 月17 日,對來自15 個種源的與生長、葉片和節間長度有關的性狀進行了相關性分析和遺傳變異分析,為幼苗的生長研究提供參考。
1 材料和方法
1.1 試驗材料
2017 年,在北京林業大學科技溫室(40°0' N,116°20' E)共測量并記錄了77 株產自我國15 個種源地的小葉楊一年生扦插苗個體。種源地自然概況見表1。測定時間為小葉楊全生長季(從2017 年4 月29 日至6 月17日),使用同樣生理年齡的插穗進行無性系擴繁,每個系號4 次重復。即每隔15~ 20 d 利用直尺測量、記錄一次樣本生長的表型性狀,包括株高、葉片數、節數和節間長度,共測量和記錄4 次,其中節間長度是從地徑部分開始從下往上用直尺測量節與節之間長度。

表1 小葉楊種源地的地理位置與氣候條件Table 1 Location and climatic conditions of different provenances of P.simonii
1.2 數據處理與統計分析
為了分析小葉楊幼苗節間長度之間的相關性和減少計算復雜度,借助R 語言軟件進行因子分析,提取代表大部分特征的公因子(旋轉后的因子分析其累積方差貢獻率達到65%以上即可)。在進行因子分析之前,首先通過KMO(Kaiser-Meyer-Olkin)檢驗[13]是否適合做因子分析。KMO 檢驗值在[0,1]區間。定義如下:

KMO 檢驗值可以通過R 軟件中的psych 包(1.8.4 版)[14]進行計算。
其次,基于Matlab 2016b 軟件,將3 個表型特征(株高、葉片數和節數)和因子分析得到的公因子作為輸入值代入KABC 進行聚類分析。參數設定:根據R 軟件MClust 包中的14 個指標,確定最佳聚類數 。小葉楊的樣本集X=(X1,X2,……XN),N(樣本量)分別等于80,78,79 和77(4 個時期)。Xi(i=1,2,……N)表示1*7 的向量(株高、葉片數、節數和4 個公共節間長因子)。種源大小(SN)為20,最大迭代次數為100,同一食物源(聚類中心)重復使用次數為10[9]。經過人工蜂群算法的一次聚類中心的優化結合K-means 聚類算法的一次聚類,交互進行,直到算法收斂或達到最大迭代次數即輸出聚類中心(K×7)和聚類結果。通過聚類結果和聚類圖可以將不同地區的樣本進行明確分類,以分析樣本和地區間的關系。
最后利用SPSS 21.0 軟件對小葉楊幼苗進行統計檢驗以分析其變異性,試驗測定其表型性狀的平均值、標準差、變異系數CV和表型分化系數(Vst),以方差的百分比表示群體間的成分,并解釋了群體間和群體內表型數據的分化程度。CV和Vst計算如下:
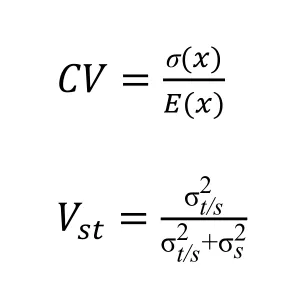
式中,σ(x)代表種源間樣本標準差,E(x)代表種源間樣本均值,代表種源間的方差分量,代表種源內的方差分量[15]。
2 結果與分析
2.1 小葉楊幼苗節間長度性狀定量分析
經計算,4 個時期(4 月29 日、5 月20 日、6 月4 日、6 月17 日)的KMO 值均屬于[0.7,0.8]區間,說明樣本數據適合做因子分析。分析計算公因子個數,結果如圖1。由圖1 表明,主成分分析(PCA)建議提取四個公因子;因子分析(EFA)建議提取三個公因子。從EFA 的結果來看,前四個特征值(三角形)在Scree 測試中高于拐點,因此,為了最大限度地保留原始數據信息,決定提取四個公因子(表2)。
由表2 可知,4 個時期的累計貢獻率為69.07%~76.69%,所以對應的4 個公因子代表了原始數據的大部分信息。4 個時期因子分析載荷值明顯分為4 個區域,D 和C 部分的方差貢獻率在4 個時期均比B 和A部分高,說明D 和C 部分的幼苗生長速度較快。且每一個公因子都由相鄰節長變量組成(載荷較大),表明相鄰節長度具有較強的相似性。

圖1 評估因素數量Figure 1 Number of assessment factors

表2 小葉楊4 個時期節間長度變量的因子分析Table 2 Factor analysis of internode length variables of P.simonii at 4 measurement days
2.2 小葉楊幼苗表型性狀聚類分析
首先利用R 軟件中mclust 包的14 個索引獲得推薦聚類結果,結果顯示BIC 值在4 個時期中值均為2。因此,確定最佳聚類數為2,如圖2。然后利用KABC 算法進行迭代計算,最終得到了4 個時期內小葉楊幼苗樣本的聚類結果,如圖3。
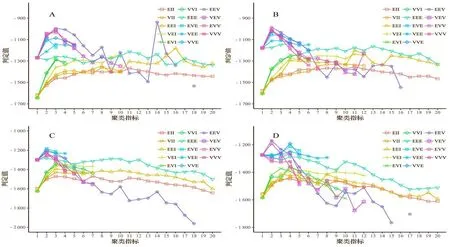
圖2 4 個日期的小葉楊幼苗推薦聚類數Figure 2 Recommended cluster number of P.simonii seedlings at 4 measurement date
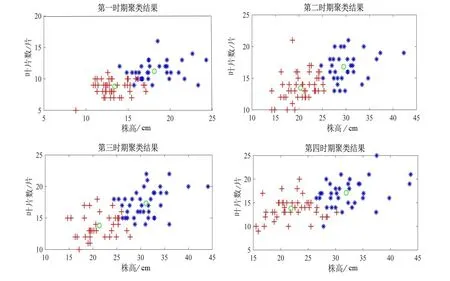
圖3 KABC 算法對4 個時期小葉楊幼苗的分類結果Figure 3 Classification maps by KABC algorithms for P.simonii seedlings at 4 measurement dates

表3 4 月29 日和5 月20 日小葉楊幼苗平均株高、葉片數和節數Table 3 Average height,number of leaf and internode of P.simonii seedlings on April 29th and May 20th

表4 6 月4 日和6 月17 日小葉楊幼苗平均株高、葉片數和節數Table 4 Average height,number of leaf and internode of P.simonii seedlings on June 4th and June 17th
2.3 小葉楊幼苗不同種源間表型變異分析
由表5 可知,株高、葉片數和節數3 個性狀種源間差異明顯。陜西麟游縣種源在株高、葉片數和節數上指標值均最大,平均海拔為1 271 m;陜西富縣株高、葉片數和節數在種源間差異次之,平均海拔為1 200 m。對同在西北地區的青海省和寧夏回族自治區而言,寧夏中寧縣種源在株高性狀上差異均值僅略大于陜西麟游縣,平均海拔為2 955 m;貴德縣種源均值在株高和節數性狀上小于麟游縣,平均海拔為2 200 m,都蘭縣和興海縣種源在株高性狀上表現相似且均值均小于貴德縣,平均海拔分別為3 180 m 和3 924 m。整體表現為西北地區的這些種源株高較高和節數較多,隨著海拔升高,株高和節數值均降低。遼寧蓋州種源在株高和葉片數性狀上最小,表現為遼寧地區種源株高較矮,葉片數較少。

表5 小葉楊表型性狀的描述性統計分析(平均值±標準差)Table 5 Phenotypic traits of different provenances of P.simonii
小葉楊節長性狀在種源間也存在很大差異,遼寧蓋州種源中下部節長均值在種源間差異最大,但標準差差異最小;而西北地區陜西富縣、麟游縣和寧夏中寧縣種源中下部節長均值次之,標準差差異明顯;青海門源縣種源中下部節長標準差最大,表現為遼寧地區種源中下部節長種源間占植株比例波動較小,位于西北地區的4個種源中下部節長占比差異較大。北京陶然亭公園種源中上部節長均值較大,平均海拔為43.5 m,河北赤城縣和內蒙古包頭市種源次之,且赤城縣種源均值大于內蒙古包頭市種源,兩地區平均海拔分別為945 m和2 000 m,表現為華北地區種源中上部節長占植株比例較大,隨著海拔的升高,中上部節長比例呈減小的趨勢。
2.4 小葉楊幼苗種源內表型變異分析
從表6 可知,性狀變異都受地區差異影響,以葉片數變異系數波動較小,中上部節長變異系數最大,中下部節長次之。株高、葉片數和節數變異系數小于4 個節長公因子(下部節長、中下部節長、中上部節長和上部節長)變異系數,表明株高、葉片數和節數性狀較穩定,受較高的基因控制,4 個節長公因子穩定性差,有較多變異可能性。

表6 小葉楊表型性狀的變異系數Table 6 Coefficient of variation of phenotypic traits of different provenances of P.simonii
對不同種源變異綜合分析可知,平均變異系數為6.09%~ 40.22%,變化幅度較大。其中西北地區寧夏中寧縣、青海門源縣和陜西麟游縣種源性狀變異系數較大,平均變異系數分別為40.22%,39.48%和38.18%。說明西北地區表型多樣性較豐富,遼寧蓋州縣平均變異系數最小(6.09%),表明東北地區種源多樣性較低。河北赤城縣、張家口和內蒙古包頭市平均表型變異系數在22.65%~ 29.46%,波動不大,三個地區年均降水量在400~421.8 mm,表明這些地區種源多樣性相似,與相似的氣候條件有關。
2.5 小葉楊幼苗種源間表型分化分析
由表7 可以看出,株高在種源內方差分量較大,葉片數和節數次之且差異相似,4 個節長公因子分化幅度較小。各性狀分化幅度為6.9%~ 12.2%,其中株高的分化系數最大為12.2%,節數的分化系數也達到了11.9%,葉片數的分化系數最小為6.9%。各個性狀在種源內的方差明顯大于種源間方差約4.56 倍,說明表型特征在種源內變異大于種源間變異且主要來源于種源內變異。7 個性狀的平均表型分化系數為17.6%,株高和節數性狀占較大比例,表明株高和節數在幼苗種源內變異較大。
3 討論
目前對小葉楊幼苗節間長度性狀的分析尚未見報道,而節間長度也反映了幼苗的生長狀況,如王健等[6]探討了普通小麥節間長度與株高的關系,結果表明矮稈小麥是通過縮短各節間長度來降低株高的。所以通過探究小葉楊節間長度之間的關系可以得到影響幼苗生長的重要部位,由于試驗得到4 個時期上部節長(D 部分)和中上部節長(C 部分)的方差貢獻率均比中下部節長(B 部分)和下部節長(A 部分)高,可推測這兩部分在幼苗生長中占據重要地位,對下一步群體基因組變異研究具有借鑒意義。
對小葉楊幼苗種源進行表型性狀的變異分析發現,15 個種源的7 個性狀在種源內和種源間均存在很大差異,且種源內變異大于種源間變異,這與衛尊征等[12]對小葉楊5 個群體的研究結果和Du 等[3]對毛白楊表型性狀的變異分析結論一致。這表明表型變異是小葉楊種源甚至楊屬Populus的普遍遺傳特性。這種形態特征的變異有利于小葉楊幼苗適應環境,種源內基因型的豐富性表現為表型性狀的多樣性,使種源能夠適應不同的環境[16],這也許造就了小葉楊生長優良、壽命長、能在復雜氣候條件下生存的特點。
4 結論
對15 個種源小葉楊一年生幼苗表型性狀分析發現節間長度性狀間存在很大聯系,將23~ 26 個節長性狀提取為4 個公因子,將植株整體分為了4 部分,且每部分之間都存在較強的相關性,即由相鄰節長組成(載荷較大),說明幼苗相鄰節間長具有較穩定的遺傳特征。4 個時期上部節長(D 部分)和中上部節長(C 部分)的方差貢獻率均比中下部節長(B 部分)和下部節長(A 部分)高,說明幼苗D 和C 部分形態特征變化較大,這與植株自身生長情況相符。
通過對小葉楊幼苗均值和標準差統計結果得到西北地區種源株高較高和節數較多,隨著海拔升高,株高和節數值均降低;遼寧蓋州種源在株高和葉片數最小,表現為遼寧地區種源株高較矮,葉片數較少。此結果和文中KABC 聚類算法得到的結果相同,聚類結果中第二類樣本主要分布在西北的陜西、青海和寧夏等地,平均株高、平均葉片數和平均節數值在4 個時期明顯比第一類樣本大。
研究結果也表明,小葉楊幼苗種內也存在極其豐富的種源間和種源內變異,15 個種源幼苗的7 個表型性狀平均變異系數在6.09%~ 40.22%,變異幅度較大,其中以中上部節長度變異系數最大,株高、葉片數和節數變異小于4 個節長公因子變異系數,表明株高、葉片數和節數性狀受較高的基因控制,4 個節長公因子變異較豐富。其次,7 個表型性狀分化幅度為6.9%~ 12.2%,株高的分化系數最大為12.2%,各個性狀在種源內的方差分量明顯大于種源間方差分量,說明表型性狀在種源內變異大于種源間變異,即種源內的多樣性程度大于種源間的多樣性程度。
