鹽度對大麻哈魚幼魚存活率、鰓ATP酶活力及其組織結構的影響
2020-06-12 12:51:38李培倫王繼隆魯萬橋崔康成高文燕
水生生物學報 2020年3期
李培倫 劉 偉 王繼隆 魯萬橋 崔康成, 高文燕,
(1. 中國水產科學研究院黑龍江水產研究所, 哈爾濱 150070; 2. 上海海洋大學水產與生命學院, 上海 201306)
一般來講, 魚類為更好地適應其生存的水環境,在長期進化過程中形成了良好的滲透壓調節機制,面對水體環境的變化, 能夠及時做出適應性調節以便更好地生存, 其中鹽度作為水體環境中重要的生態因子, 會對魚類的生存、生長發育、新陳代謝和滲透壓等產生一定程度的影響[1,2]。魚鰓不僅是絕大多數硬骨魚類的呼吸器官, 同時還具備調節機體滲透壓平衡、體液pH、離子轉運和排泄氨氮等重要功能[3,4]。諸多研究表明[5—10], 廣鹽性魚類具有較高的鹽度生存范圍, 在其海淡水適應過程中, 魚鰓的結構、生理功能等會產生明顯的適應性改變, 主要體現在鰓絲寬度、鰓小片大小及間距、線粒體豐富細胞的數量及其頂膜形態、ATP酶活力等方面。
大麻哈魚(Oncorhynchus ketaWalbaum), 俗稱太平洋鮭、秋鮭、大馬哈魚等, 屬鮭形目(Salmoniformes)、鮭科(Salmonidae)、大麻哈魚屬(Onco-rhynchus), 其自然分布于北緯35°—73°, 東經120°—123°的太平洋、北冰洋海域及其有出海口的河流,終生只進行一次繁殖, 繁殖后親體死亡, 為典型的溯河生殖洄游型冷水性魚類[11,12]。大麻哈魚在進入其出生地淡水河流產卵繁殖之前需要在海洋中度過大部分生命旅程, 淡水中孵化生長的幼魚在體重1 g左右時開始由淡水進入海水進行鹽化過渡以逐步適應高鹽度的海水環境[13]。目前, 國內外有關大麻哈魚的研究報道較多, 主要集中在種群結構生物學[14—17]、耳石標記技術[18,19]、嗅覺印記[20]、幼魚生長特征[21—23]等方面, 而有關在不同鹽度條件下大麻哈魚幼魚鰓絲結構形態、ATP酶活力等方面的報道較少。本文對不同鹽度條件下大麻哈魚幼魚成活率、魚鰓組織結構及其ATP酶活力響應情況等進行了研究, 不僅能夠豐富大麻哈魚在降海洄游方面的生理生態資料, 還可為進一步探索其降海洄游過程中滲透壓調節機理提供科學基礎。
1 材料與方法
1.1 試驗材料
2017年10月下旬, 在烏蘇里江海清鄉江段采捕大麻哈魚繁殖親本, 運至湯原縣大麻哈魚孵化繁育基地進行采卵授精孵化。2018年4月上旬, 將大麻哈魚魚苗運至中國水產科學研究院黑龍江水產研究所養殖車間, 在全自動循環控溫水族箱(80 cm×60 cm×50 cm)內進行飼養。
1.2 試驗方法
試驗設計試驗共設置4個鹽度組S0、S8、S16和S24, 對應鹽度分別為0、8‰、16‰和24‰,其中S0為淡水對照組。試驗用水為地下井水與海水素調配而成, 鹽度計校正各試驗組鹽度。挑選生長狀況良好且個體接近的大麻哈魚幼魚隨機分配放入各組, 每個試驗組各設3個平行(每個平行30尾), 每個鹽度組為90尾實驗魚。采用逐步過渡法進行鹽度馴化, 每天上升4個鹽度, 直至達到各試驗組鹽度設定, 試驗周期為42d, 期間統計大麻哈魚幼魚存活率。實驗魚初始叉長為(5.27±0.24) cm, 體質量為(1.01±0.14) g。試驗期間水溫控制在(14±0.50)℃, 每天投喂鮭魚顆粒飼料2次, 每3天換水一次, 試驗期間養殖車間其他管理條件一致。
魚鰓ATP酶活力測定取樣前24h停止投喂,用MS-222麻醉劑麻醉魚體。每個鹽度組隨機取6尾實驗魚置于冰盤上進行解剖, 取出魚鰓后用預冷生理鹽水快速沖洗并用吸水紙吸干水分, 保存于冰箱(-80℃)中。測定前對組織解凍, 準確稱取組織重量, 按重量(g)∶體積(mL)=1∶9的比例加入9倍的生理鹽水, 冰水浴條件下機械勻漿, 2500 r/min, 離心10min, 取上清再用生理鹽水5倍稀釋, 取樣50 μL進行總蛋白及酶活力測定。Na+/K+-ATPase酶和Ca2+/Mg2+-ATPase酶活力采用試劑盒(南京建成)檢測, 按照說明書進行操作。
鰓絲組織結構觀察波恩試液固定各鹽度組魚鰓用于常規組織學觀察, 固定24h后轉入70%酒精中保存。后經梯度酒精脫水、常規石蠟包埋與切片(厚度6 μm)、HE染色、二甲苯透明, 中性樹膠封片保存。Olympus BX53顯微鏡觀察,Olympus DP27顯微拍攝系統進行拍照并測量鰓絲寬度、鰓小片長度、鰓小片寬度和鰓小片間距。
鰓絲掃描電鏡制樣用2.5%濃度的戊二醛(pH 7.2)對各鹽度組魚鰓進行固定且在4℃條件下保存。固定好的樣品先用磷酸緩沖溶液沖洗3次,然后用1%濃度的鋨酸再固定2h, 再用磷酸緩沖溶液沖洗3次。梯度酒精脫水, 叔丁醇置換, 冷凍干燥儀干燥后鍍膜, 最后用SU-8010型掃描電子顯微鏡觀察拍照。
1.3 數據處理
運用Excel2007和SPSS19.0對實驗數據進行統計分析, 采用單因素分析法進行方差分析, Duncan法進行組間多重比較, 數據結果用平均值±標準差(Mean±SD)表示,P<0.05為差異顯著, 最后用Graph-Pad Prism 5軟件作圖。
其中: 存活率(Survival rate, %)=100×(Nt/N0)。式中,t為實驗天數(d),Nt為大麻哈魚幼魚終末尾數,N0為大麻哈魚幼魚初始尾數。
2 結果
2.1 鹽度對大麻哈魚幼魚存活率的影響
如表1所示, 在試驗結束后S0組共死亡5尾幼魚, 存活率為94.45%; S8和S16組死亡數量最少, 均為1尾, 存活率達到98.89%; 而S24組幼魚死亡數量最多, 為15尾, 存活率為83.34%, 大麻哈魚幼魚存活率隨著鹽度的升高呈現出先升高后下降的趨勢。其中在鹽度馴化過渡期間, 除S24組死亡1尾大麻哈魚幼魚之外, 其余各組均正常, 無死亡現象。
2.2 鹽度對大麻哈魚幼魚魚鰓ATP酶活力影響
從試驗結果可以看出, 大麻哈魚幼魚魚鰓Na+/K+-ATP酶活力為0.2924—1.1294 U/mg, 而Ca2+/Mg2+-ATP 酶活力為0.3665—0.8353 U/mg, 各鹽度組之間的酶活力單因素方差分析結果見圖1。隨著鹽度的升高, 大麻哈魚幼魚魚鰓中Na+/K+- ATP和Ca2+/Mg2+-ATP 酶活力均呈現出先升高后下降趨勢。其中Na+/K+-ATP酶活力最高的是S8組, 最低的為S24組, 且S8組與S0組、S24組之間具有顯著性差異(P<0.05), 而S16組與其余各組之間均無顯著性差異(P>0.05); 不同鹽度組之間Ca2+/Mg2+-ATP酶活力由高到低依次: S8組、S16組、S0組和S24組; 單因素方差分析結果來看, S8組與S0、S24組之間具有顯著性差異(P<0.05), 而與S16組之間無顯著差異(P>0.05); S24組與S8、S16組之間具有顯著性差異(P<0.05), 而與S0組之間無顯著差異(P>0.05)。

表1 在不同鹽度條件下大麻哈魚幼魚的存活率Tab. 1 Survival rates of juvenile chum salmon under different salinities
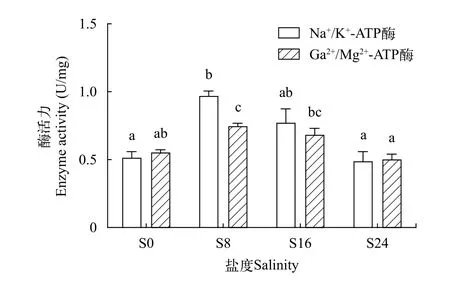
圖1 不同鹽度下大麻哈魚幼魚魚鰓ATP酶活力Fig. 1 ATP enzyme activity levels of juvenile chum salmon at different salinities
2.3 不同鹽度下大麻哈魚幼魚魚鰓組織學觀察
在光學顯微鏡下觀察到S0組大麻哈魚幼魚鰓絲主干較窄, 寬度約(12.71±1.51) μm, 由1—2層上皮細胞組成且排列緊密, 上皮細胞多呈圓形、長橢圓形; 線粒體豐富細胞主要分布在鰓絲與鰓小片交匯區域, 胞體較大但數量較少, 呈橢圓狀, 著色較深(圖版Ⅰ-1、2); 鰓絲頂端鰓小片與鰓絲基部融合,主干部位含大量軟骨細胞, 末端逐步收縮呈平滑的橢圓弧狀且內含血細胞(圖版Ⅰ-3、4); 鰓絲上的鰓小片內柱細胞多呈圓柱形或長梭形, 柱細胞之間連接形成血管腔, 內含紅細胞, 而扁平上皮細胞則位于鰓小片邊緣(圖版Ⅰ-2)。鰓小片在所有試驗組中長度最長, 約為(68.79±13.82) μm; 寬度最寬, 約為(8.35±1.32) μm; 間距最小約為(11.47±1.72) μm(表2)。
S8組大麻哈魚幼魚鰓絲主干同S0組相比稍寬,約(24.32±3.32) μm, 其內部上皮細胞層數增多且形狀不一, 多呈卵圓形、圓形、長梭形等, 線粒體豐富細胞數量增多, 體積增大, 染色深淺不一(圖版Ⅰ-5、6); 鰓絲頂端鰓小片逐漸變短直至完全同鰓絲合為一體, 軟骨細胞排列緊密, 細胞核明顯, 末端呈規則的橢圓弧狀(圖版Ⅰ-7、8); 鰓小片內部柱細胞著色較深, 呈明顯的長梭形, 扁平上皮細胞位于邊緣區域(圖版Ⅰ-6)。鰓小片大小較S0組相比有所減小, 其長度約為(65.42±5.45) μm, 寬度約為(7.77±0.89) μm, 鰓小片間距亦有所減小, 約為(12.29±2.56) μm(表2)。
S16組大麻哈魚幼魚鰓絲主干同S0、S8組相比逐漸增寬, 約(28.71±3.08) μm, 線粒體豐富細胞數量增多, 集中在鰓絲與鰓小片基部, 細胞較大, 核明顯(圖版Ⅰ-9、10); 鰓絲頂端逐步收縮呈橢圓弧狀,能夠清晰觀察到軟骨細胞和血細胞(圖版Ⅰ-11、12)。鰓小片大小較S0、S8組相比繼續減小, 其長度約為(58.11±10.99) μm, 寬度約為(6.74±0.73) μm,但鰓小片間距較S0、S8組有所增大, 約為(13.06±3.00) μm(表2)。
S24組大麻哈魚幼魚鰓絲及鰓小片的結構變化同其他試驗組相比, 鰓絲主干最寬, 約為(33.68±2.81) μm; 鰓小片最短, 約為(52.52±7.30) μm, 且其基部膨大彎曲(圖版Ⅰ-13); 線粒體豐富細胞數量增多, 主要分布在鰓絲主干邊緣部位, 鰓小片基部區域亦有分布; 柱細胞分布于鰓小片內部區域呈長梭形或卵圓形, 扁平上皮細胞位于鰓小片邊緣區域(圖版Ⅰ-14); 鰓絲頂端逐漸收縮呈規則的橢圓弧狀, 與其他試驗組無明顯區別, 其內軟骨細胞和紅細胞明顯(圖版Ⅰ-15、16)。S24組大麻哈魚幼魚鰓小片間距最小, 約為(12.02±2.68) μm, 但其鰓小片寬度較S16組有所增加, 為(6.28±0.75) μm(表2)。

表2 鹽度對大麻哈魚幼魚鰓組織結構的影響Tab. 2 Effects of salinity on the gill structures of juvenile chum salmon
2.4 不同鹽度下大麻哈魚幼魚魚鰓掃描電鏡觀察
在實驗結束時, 不同鹽度下大麻哈魚幼魚魚鰓掃描電鏡觀察結果見圖版Ⅱ。S0組大麻哈魚幼魚鰓絲表面布滿了扁平上皮細胞、線粒體豐富細胞和黏液細胞等, 其中以扁平上皮細胞為主, 線粒體豐富細胞和黏液細胞數量較少(圖版Ⅱ-1)。扁平上皮細胞界限輪廓較清晰, 呈不規則多邊形, 表面覆滿迷宮狀環形微脊, 微脊之間具有深溝; 黏液細胞位于相鄰扁平上皮細胞之間, 呈細長窄條狀, 表明凹凸不平, 起伏較大; 線粒體豐富細胞頂膜位于扁平上皮細胞之間, 大多數位于鰓絲靠近鰓小片一側,頂膜相對平整, 開口較大, 呈不規則四邊形或三角形, 且其表面存在大量的微絨毛(圖版Ⅱ-2)。鰓小片底部表層扁平上皮細胞微脊較短、呈點狀、棒狀或長弧狀, 排列密集, 其上含有大量的氣孔(圖版Ⅱ-3、4)。
S8、S16和S24組大麻哈魚幼魚鰓絲表層結構與S0組相似, 隨著鹽度的升高, 鰓絲表層線粒體豐富細胞數量有所增加, 但其頂膜開口不斷變小; 其中S8組和S16組鰓絲表面扁平上皮細胞之間的輪廓更加清楚且環形微脊條紋清晰(圖版Ⅱ-5、9), 而S24組鰓絲表面扁平上皮細胞之間界限模糊, 表面環形微脊變短, 隆起的微脊上有細小的橫突且微脊之間有融合或間斷情況(圖版Ⅱ-10); S8、S16和S24組線粒體豐富細胞頂膜呈不規則的橢圓形, 其中S8線粒體豐富細胞頂膜含有少量的微絨毛(圖版Ⅱ-6), 而S16、S24組線粒體豐富細胞頂膜平整稍向外凸且其表面沒有微絨毛(圖版Ⅱ-16、24); S8、S16和S24組鰓小片底部扁平上皮細胞迷宮樣環形微脊結構清晰, 但隨著鹽度的升高, 微脊長度明顯變短、數量增多且間距逐漸變小(圖版Ⅱ-7、11、15); 鰓小片底部扁平上皮細胞表層氣孔數量逐漸減少且孔徑變小(圖版Ⅱ-8、12、16)。
3 討論
3.1 鹽度對大麻哈魚幼魚鰓ATP酶活力影響
ATP酶是魚鰓執行離子調控功能的重要膜結合蛋白酶, 主要為離子轉運提供載體和驅動力, 其中Na+/K+-ATP酶被稱作Na-K泵, 主要作用是向胞內運輸K+的同時向胞外排出Na+, 而Ca2+/Mg2+-ATP酶能夠將胞內Ca2+轉移至胞外, 使胞內Ca2+維持在較低水平[6]。由于自然界中魚類種類繁多, 其生活環境有所不同進而導致有關鹽度對魚鰓Na+/K+-ATP酶活力影響的研究形成了兩種觀點, 一種為鹽度對魚鰓Na+/K+-ATP酶活力具有顯著的影響, 如銀鮭(Oreochromis mosambicus)[24]、銀鯧(Pampus argenteus)[6]、大菱鲆(Scophthalmus maximus)[25]等, 而另一種觀點則認為鹽度對其無顯著影響, 如多鰭南極魚(Notothenia neglecta)[26]。本文研究結果顯示,隨著鹽度的升高, 大麻哈魚幼魚魚鰓中Na+/K+-ATP和Ca2+/Mg2+-ATP 酶活力均呈現出先升高后下降趨勢且酶活力最高的均為S8組、最低的為S24組, 其中鹽度組(S8、S16、S24) ATP 酶活力隨鹽度的升高呈現下降的趨勢, 說明鹽度對大麻哈魚魚鰓ATP酶活力具有顯著的影響, 但其活力同鹽度之間的變化趨勢又與其他學者的研究結果有所不同, 如在不同鹽度中養殖60d的大菱鲆(Scophthal-mus maximus)[25]、養殖30d的四指馬鲅(Eleutheronema tetradactylum)[27]等其Na+/K+-ATP酶活力均隨著鹽度的升高而增大, 這可能與大麻哈魚的生理生態習性有關。大麻哈魚作為海淡水洄游型廣鹽性魚類, 具有較大的鹽度適應范圍, 推測體重1 g左右的幼魚在降海過程中能夠很好的適應低鹽度的海水環境, 但當鹽度超出其適應范圍后ATP酶活力反而會下降, 不能夠更好的調節細胞內外的離子濃度及其滲透壓平衡。
3.2 鹽度對大麻哈魚幼魚鰓組織結構的影響
鰓是魚體與其生存的水環境之間進行物質交換的媒介, 鰓絲形態結構及各類細胞對其發揮正常生理功能具有重要作用[7]。大麻哈魚為海淡水洄游型廣鹽性魚類, 不僅能在高鹽度海水中生存, 亦可在淡水中生活, 受棲息水環境鹽度的影響鰓組織會發生一定程度的適應性改變。本文結果可以看出生活在S0組淡水環境中的大麻哈魚幼魚, 其鰓絲寬度較窄、鰓小片長且寬、細胞排列緊密, 這些結構形態能夠保障魚鰓與水體充分接觸進而攝取其中的無機離子維持滲透壓平衡, 這與魏渲輝等[2]的研究結論相似。而生活在不同鹽度水體(S8、S16、S24)中的幼魚, 其鰓絲寬度隨著鹽度的升高逐漸增厚且各鹽度組之間呈現出顯著性差異, 鰓小片的長度和寬度亦隨著鹽度的升高發生一定程度的萎縮,鰓小片間距同淡水組相比有所增大, 這一結果與鯔(Mugil cephalus)[7]、虹鱒(Salmo cairdneri)[28]等研究相一致, 可能為適應水體高滲環境, 增加水氧交換量以滿足其生長需求而發生結構上的適應性改變。
3.3 鹽度對大麻哈魚幼魚線粒體豐富細胞和扁平上皮細胞的影響
研究表明[2,9,10], 無論是生活在海水中的魚類還是淡水環境中的魚類, 鰓絲上都有線粒體豐富細胞的存在, 但由于魚類生活的環境不同, 線粒體豐富細胞的數量和結構會存在較大的差異。大麻哈魚幼魚線粒體豐富細胞主要分布于鰓絲與鰓小片相交區域, 且隨著水體鹽度的升高向相鄰鰓小片中間擴散, 此外其數量逐漸增多、胞體變大、頂膜變小且微絨毛消失, 這些變化可有效提高線粒體豐富細胞的泌氯功能和代謝水平以便更好地適應高鹽度環境。大麻哈魚幼魚鰓絲表層絕大部分區域被扁平上皮細胞所覆蓋, 其主要以被動運輸的方式參與魚鰓的生理功能[10], 此外, 該類細胞表面存在的微嵴、溝、孔等結構致使鰓絲表面凹凸不平, 對于提高鰓絲表面的阻礙作用、延長水流的停留時間有重要的作用[29]。通過掃描電鏡觀察大麻哈魚幼魚鰓絲基部表面結構可以看出, 在不同鹽度條件下扁平上皮細胞表面迷宮狀環形微脊亦有所變化, 同S0組相比, S8組和S16組鰓絲表面扁平上皮細胞之間的輪廓更加清楚且環形微脊條紋清晰, 而S24組鰓絲表面扁平上皮細胞之間界限模糊, 表面環形微脊變短, 隆起的微脊上有細小的橫突且微脊之間有融合或間斷情況, 此外, 隨著鹽度升高鰓小片底部扁平上皮細胞表層結構亦有清晰逐漸變得混亂、氣孔數量逐漸減少且孔徑變小, 通過這些變化可直觀看出扁平上皮細胞對水環境中鹽度的響應結果,同時推測這些微嵴、溝、孔等的結構形態可作為鑒別魚體是否適應鹽度的標準, 而降海階段的大麻哈魚幼魚適宜的鹽度生存范圍可能介于8—16, 具體有待進一步的深入研究。

圖版Ⅰ 鹽度對大麻哈魚幼魚鰓組織的影響PlateⅠ The effect of salinity on the gills of the juvenile chum salmon juveniles

圖版Ⅱ 不同鹽度下大麻哈魚幼魚鰓絲表層結構掃描電鏡圖PlateⅡ Scanning electron micrographs of the surface structure of the gill filaments in chum salmon at different salinities
