基于拉格朗日粒子追蹤的黃海中南部小黃魚幼體早期輸運初步研究
2020-07-02 02:37:16黃康康葉振江于海慶田永軍
浙江海洋大學(xué)學(xué)報(自然科學(xué)版) 2020年6期
關(guān)鍵詞:模型
黃康康,葉振江,于海慶,田永軍
(中國海洋大學(xué)水產(chǎn)學(xué)院,山東青島 266003)
小黃魚Larimichthys polyactis 是廣泛分布在我國黃海、渤海和東海的重要經(jīng)濟魚類[1],是中國、日本和韓國重要的捕撈魚種,在中國漁業(yè)中占有重要的地位[2]。其資源經(jīng)歷衰退期和恢復(fù)期2 個階段[3]。從20 世紀(jì)60 世紀(jì)中葉到20 世紀(jì)80 年代初,山東省小黃魚產(chǎn)量始終處于較低水平。為恢復(fù)小黃魚資源量,中國政府采取了諸如休漁制度及產(chǎn)卵場保護等一系列措施[2]。但是,小黃魚群體結(jié)構(gòu)趨于簡單,體型趨于小型化、低齡化且性成熟加快等問題使得種群結(jié)構(gòu)仍較為脆弱[4]。以往的研究主要關(guān)注小黃魚資源量[2,5]、生長[6-7]、攝食[7-8]、早期發(fā)育[10]等方面,對于早期生活史的報道相對較少。魚類早期的生長發(fā)育狀況往往能決定來年的補充量[11],是魚類資源保護的基礎(chǔ)[12]。
關(guān)于小黃魚早期補充的研究,李建生等[13]分析了春季長江口及鄰近海域小黃魚仔稚魚的生態(tài)分布;蔣欣等[14]對長江口及其鄰近海域的仔稚魚空間分布及其與環(huán)境之間的關(guān)系進行了分析,討論了溫度對于魚類早期階段的重要性;俞連福等[14]利用春季東海南部海域調(diào)查的仔稚魚資料同海洋環(huán)境進行分析研究,并就仔稚魚分布與溫度關(guān)系進行了討論。這些研究對于海洋魚類早期分布的影響機制尚未有深入涉及。
近年來,水動力模型結(jié)合生物粒子為研究海洋生物早期分布及影響機制提供了重要的視點[15]。李曰嵩等[16]利用三維數(shù)值模型對臺灣暖流、黑潮等驅(qū)動下的鮐魚Scomer japonicus 魚卵進行追蹤,模擬了早期魚卵和仔魚的輸運過程及豐度分布;羅曉凡等[17]通過模擬含潮汐過程和不含潮汐過程的粒子運動過程,對大型水母的運移規(guī)律進行了細致分析;LACROIX,et al[18]利用水動力模型,模擬了比目魚Solea solea 早期生長階段的種群移動動態(tài);LANGANGEN,et al[19]利用水動力模型對黑線鱈Melanogrammus aeglefinus 進行了多年粒子追蹤,探究了不同年份黑線鱈產(chǎn)卵場的變動。這些研究利用水動力模型研究了早期仔稚魚等在水體中的移動過程,對揭示魚類早期生活史過程與資源動態(tài)的環(huán)境影響機制起到了重要作用。
本文采用水動力模型對黃海中南部近岸水域產(chǎn)卵場調(diào)查的小黃魚魚卵主要分布區(qū)進行粒子追蹤模擬,研究小黃魚早期階段分布的年際變化及分布的影響機制,并就小黃魚稚魚主要聚集區(qū)1953-2018 年5-7 月平均海表溫度同山東省小黃魚捕撈量進行分析,探究小黃魚早期分布情況與資源量動態(tài)的關(guān)系。
1 材料與方法
1.1 環(huán)流數(shù)據(jù)
本文使用Finite Volume Arctic Ocean model(麻省大學(xué)海洋生態(tài)動力模擬實驗室(UMASS,MEDML)研發(fā)的有限體積海岸海洋模型)模型輸出的2011-2018 年黃渤海環(huán)流數(shù)據(jù)驅(qū)動粒子追蹤。該模型的溫鹽初始場采用HYCOM 上的數(shù)據(jù),氣象強迫數(shù)據(jù)采用美國國家環(huán)境預(yù)報中心(NCEP)發(fā)布的氣候預(yù)報系統(tǒng)的再分析數(shù)據(jù)(CFSV2,https://rda.ucar.edu/datasets/ds093.1/)。為更真實反映海洋環(huán)境變動,模型加入海表面高度(SSH)及溫度(SST)同化。該模型在開邊界深海大洋處水平分辨率為10 km 左右,在模型關(guān)注的近岸海域為1.5 km 左右。
1.2 生物數(shù)據(jù)處理
魚卵數(shù)據(jù)來源于黃海中南部近岸水域產(chǎn)卵場調(diào)查,調(diào)查時間為2015 年5 月15-25 日、2017 年5 月24-29 日、2018 年5 月25-30 日,每站采用大型浮游生物網(wǎng)(網(wǎng)口直徑0.8 m,網(wǎng)目0.505 mm,網(wǎng)口內(nèi)徑0.5 m,網(wǎng)衣2.8 m)進行水平拖網(wǎng)10 min,所采集樣品以5%福爾馬林溶液進行固定保存,樣品分析主要參考《中國近海魚卵和仔魚》及相關(guān)參考文獻[20]。圖1 為2015、2017、2018 年小黃魚魚卵豐度分布圖,小黃魚魚卵豐度為100 m3水體中出現(xiàn)的魚卵粒數(shù)(ind.·100-1·m-3),計算公式為:

式中:G 為單位體積海水中魚卵粒數(shù)(ind.·100-1·m-3);N 為各站位全網(wǎng)魚卵數(shù)量(ind.);V 代表濾水體積(m3),即網(wǎng)口面積、流量計轉(zhuǎn)數(shù)、校正系數(shù)(0.3)三者的乘積。
1953-2018 年山東省小黃魚捕撈產(chǎn)量數(shù)據(jù)來源于http://www.fishdata.cn/web。小黃魚稚魚主要聚集區(qū)判定標(biāo)準(zhǔn)為單位面積粒子密度大于4 800 的區(qū)域,稚魚主要聚集區(qū)5-7 月平均海表溫度數(shù)據(jù)來源于https://coastwatch.pfeg.noaa.gov/erddap/griddap。
1.3 拉格朗日粒子追蹤
通過對2015、2017、2018 年5 月下旬黃海中南部近岸水域產(chǎn)卵場調(diào)查采集的樣品進行分析,小黃魚魚卵主要分布在34°N 附近沿岸(圖1),因此本文的研究區(qū)域選取圖1 紅框內(nèi)海域為粒子釋放區(qū)(2011-2018 年每年5 月6 日、5 月16 日、5 月26 日各釋放2 600 個粒子,共釋放62 400 個粒子),模擬小黃魚在不同年份、不同日期從同一海域進入水體,隨水流輸運聚集的情況,模擬時間為60 天。
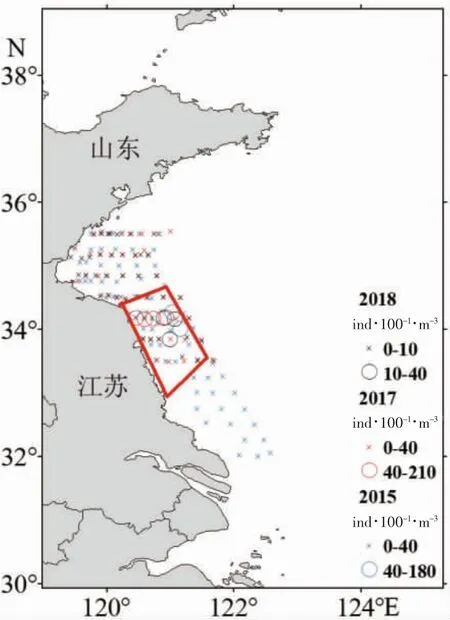
圖1 2015、2017、2018 年5 月下旬小黃魚魚卵豐度分布Fig.1 Egg density distribution of L.polyactis
本文中,假設(shè)一段時間Δt 內(nèi),魚卵從(xt,yt)位置以平均速度u、v 移動到(xt+Δt,yt+Δt)(x、y 代表經(jīng)、緯度),那么輸運過程在二維平面可簡單表示為:
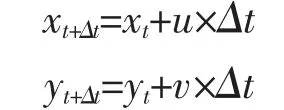
式中:xt+Δt(yt+Δt):t+Δt 時刻魚卵所在位置的經(jīng)(緯)度;
xt(yt):t 時刻魚卵所在位置的經(jīng)(緯)度;
u,v:魚卵從t 到t+Δt 時刻內(nèi)的平均流速,其中u 以東為正方向,v 以北緯正方向,單位為m·s-1。
2 結(jié)果
2.1 釋放粒子空間分布年際變化
模擬結(jié)果表明,釋放粒子均分布在33-38° N 范圍內(nèi),未超出黃海范圍(圖2)。流場和釋放日期的年際變化導(dǎo)致不同年份間粒子空間分布的差異,但同一年份不同日期釋放的粒子空間分布差異較小。所有釋放粒子表現(xiàn)出2 種分布狀態(tài):(1)2011-2016 年釋放粒子有少部分向海州灣東南部海域擴展,其余釋放粒子均分布在海州灣東北部海域;(2)2017-2018 年釋放粒子整體分布集中,覆蓋范圍較小且粒子分布位置離初始位置集中向東北、近岸。
2.2 釋放粒子溫度履歷的年際變化
釋放粒子輸運階段的溫度變化具有明顯的時間差異(圖3),主要表現(xiàn)為:(1)不同年份釋放粒子變化趨勢差異較大,2017 年粒子平均溫度變化較大,2014 年粒子平均溫度變化相對較小;(2) 不同日期釋放粒子平均溫度變化速度不同,5 月6 日釋放粒子平均溫度上升速度最快,5 月16 日釋放粒子次之,5 月26 日釋放粒子最慢。以上結(jié)果表明不同釋放年份及不同釋放日期對粒子輸運的溫度過程均有顯著影響。
2.3 模擬稚魚主要聚集區(qū)
對2011-2018 年釋放粒子運用ARCGIS 軟件點密度分析對釋放粒子最終位置進行統(tǒng)計(圖4)。結(jié)果表明,在121-123° E、34-36° N 海域內(nèi)粒子密度最高,從該區(qū)域向外密度逐漸降低。在海州灣沿岸區(qū)域粒子密度較高,但海州灣西部靠岸區(qū)域粒子分布相對較少。
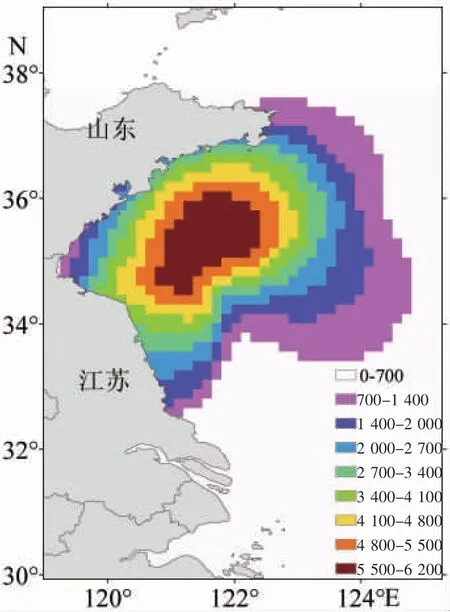
圖4 2011-2018 年輸運結(jié)束時小黃魚的稚魚空間分布Fig.4 Spatial distribution of juvenile L.polyactis at the end of the transport period from 2011 to 2018
2.4 模擬稚魚聚集區(qū)的溫度與捕撈量關(guān)系
1953-2018 年稚魚主要聚集區(qū)5-7 月份平均海表溫度與山東省小黃魚捕撈量關(guān)系如圖5 所示。20 世紀(jì)60 年代-20 世紀(jì)80 年代,隨著溫度的下降,小黃魚捕撈量呈現(xiàn)下降的趨勢,從1963 年開始,小黃魚捕撈量始終保持在較低水平,1980 年左右平均溫度達到最低點,小黃魚捕撈量也達到歷史最低點。20 世紀(jì)80 年代中葉到21 世紀(jì)初的十幾年間,溫度呈現(xiàn)出快速上升趨勢,小黃魚捕撈量于1990 年開始出現(xiàn)大幅度的增長。2000 年以后,小黃魚產(chǎn)量繼續(xù)增長,至2012 年以后有所下降;而觀察區(qū)溫度呈現(xiàn)先下降后上升的過程。

圖5 1953~2018 年山東省小黃魚捕撈量Fig.5 L.polyactis catches in Shandong Province,1953-2018
3 討論
3.1 粒子輸運的年際與季節(jié)動態(tài)
本文通過對海水表層粒子進行追蹤以探討近岸水動力過程對小黃魚稚魚分布的影響。追蹤結(jié)果表明稚魚主要分布區(qū)位于海州灣東北部海域,最北分布在38° N 附近,同一年份釋放粒子不具有明顯差異,即同樣的模擬時間下,模擬區(qū)域5 月份產(chǎn)出魚卵的輸運路徑較為統(tǒng)一(圖2)。不同年份稚魚分布存在明顯的差異,首先考慮是由風(fēng)驅(qū)動下的流場所致,黃海近岸夏季風(fēng)為南風(fēng)或東南風(fēng)[21],夏季風(fēng)的年變化使得其對海流的作用不同,較強的風(fēng)力會使得粒子向東北方向輸運的更遠。因此,粒子輸運的年際變化很大一部分原因是受到風(fēng)驅(qū)動下的流場影響。海域內(nèi)魚卵仔稚魚的種類和數(shù)量取決于整個海域的漁業(yè)資源狀況,同時又將影響整個海域魚類資源的補充[22]。輸運路徑的年際差異會對小黃魚的分布、早期生長、資源動態(tài)產(chǎn)生影響,因而對其深入研究有利于解釋資源量變動的原因。萬瑞景等[20]研究表明,7月下旬在121-123° E、35-35.5° N 區(qū)域內(nèi)存在大量仔稚魚,本次研究模擬的稚魚主要聚集區(qū)與其研究結(jié)果相一致,在一定程度上模擬了小黃魚稚魚的分布區(qū)域,該區(qū)域可能具有適宜的溫度及餌料供小黃魚早期生長發(fā)育,使得小黃魚稚魚在該區(qū)域聚集并作為棲息地。
3.2 育幼場溫度對小黃魚資源動態(tài)的影響
總體來看,小黃魚產(chǎn)量在20 世紀(jì)90 年代后的上升與溫度的上升趨勢相一致(圖5),溫度上升可能是產(chǎn)量復(fù)蘇的重要因素之一。
魚類的補充機制是一個復(fù)雜的過程,饑餓和捕食在魚類的早期生長階段非常重要[23]。但是,一些與魚類密度無關(guān)的因素,如大氣條件,海洋環(huán)境也會影響魚類的早期補充[24]。KIM,et al[25]通過研究小黃魚漁獲量與環(huán)境的關(guān)系證明了溫度對小黃魚早期補充的影響,其研究認為溫暖的產(chǎn)卵季節(jié)和穩(wěn)定的海洋環(huán)境有利于仔稚魚的生存,從而在來年產(chǎn)生較大的補充量和更高的捕撈量。MYERS,et al[26]的研究表明,幼魚期的死亡率對于資源量的補充非常重要,溫度的變化能通過影響魚類幼魚期的生長及死亡來影響成魚的漁獲量。
林龍山等[27]認為2000 年后小黃魚產(chǎn)量的增加主要是由于捕撈強度始終處于較高水平所致,并非小黃魚的資源量發(fā)生根本好轉(zhuǎn)。對于山東省小黃魚捕撈量而言,其主要影響因素由原來的受環(huán)境溫度影響轉(zhuǎn)變?yōu)槭苋祟愡^度捕撈的影響,朱曉光等[28]認為在過去的幾十年中,我國海洋漁業(yè)捕撈強度增加上百倍,對海洋魚類的利用已經(jīng)從最初的適度捕撈轉(zhuǎn)化為過度利用,人類的過度捕撈破壞了原本相對穩(wěn)定的海洋生態(tài)系統(tǒng)及食物鏈,使得漁業(yè)資源結(jié)構(gòu)更加不穩(wěn)定。本文結(jié)果顯示溫度是影響小黃魚資源量與產(chǎn)量動態(tài)的重要因素,但是小黃魚產(chǎn)量亦受其它一些人為因素如捕撈強度等影響,相關(guān)機制有待以后進一步加強研究。
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
網(wǎng)絡(luò)安全與數(shù)據(jù)管理(2022年1期)2022-08-29 03:15:20
導(dǎo)航定位學(xué)報(2022年4期)2022-08-15 08:27:00
中學(xué)生數(shù)理化·中考版(2022年8期)2022-06-14 06:55:24
新世紀(jì)智能(數(shù)學(xué)備考)(2021年9期)2021-11-24 01:14:36
成都醫(yī)學(xué)院學(xué)報(2021年2期)2021-07-19 08:35:14
新世紀(jì)智能(數(shù)學(xué)備考)(2020年9期)2021-01-04 00:25:14
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2020年10期)2020-11-26 08:24:50
數(shù)學(xué)物理學(xué)報(2020年2期)2020-06-02 11:29:24
光學(xué)精密工程(2016年6期)2016-11-07 09:07:19
