來源于Paenibacillus campinasensis SK13.001的β-環糊精葡萄糖基轉移酶在大腸桿菌中的表達和反應條件優化
2020-07-06 13:25:38姚小琳張濤江波
食品與發酵工業 2020年12期
關鍵詞:產量
姚小琳,張濤,江波
(食品科學與技術國家重點實驗室(江南大學), 江蘇 無錫, 214122)
環糊精葡萄糖基轉移酶(cyclodextrin glycosyltransferase,CGTase)是一種典型的淀粉水解酶,該酶的主要工業應用價值在于其高效的轉糖苷反應,CGTase的3種轉糖苷反應包括環化、耦合、歧化,其活性均顯著高于水解反應[1],該酶的獨特之處在于其可以通過環化反應將淀粉或淀粉類似物轉化為非還原性環狀麥芽寡糖[2]。常見的環狀麥芽寡糖為含有6、7、8個葡萄糖單元的分子,分別為α-環糊精(α-cyclodextrin,α-CD)、β-CD和γ-CD[3-5]。環糊精分子為外部親水內部疏水的中空圓筒形立體結構,這使得環糊精能與很多特定大小的疏水性分子或基團形成穩定的包合物,從而改變客體分子的理化性質,因此環糊精在食品、醫藥、化妝品等很多領域有廣泛的應用[6-7]。隨著環糊精的廣泛應用,生產環糊精的CGT酶的制備已成為當今研究的熱點[8]。目前,已發現的能分泌CGT酶的微生物種類很多,大多數為芽孢桿菌屬。但通常野生菌所產CGT酶轉化淀粉生產環糊精的轉化率較低,這與酶的菌種來源以及反應條件相關;包括底物的種類、濃度、加酶量、反應溫度及pH等條件[9]。很多研究通過優化培養基的成分和培養條件來提高菌種產酶能力,但由于野生菌本身的調控機制存在很多限制;為了提高菌種的產酶量,可以采用基因工程的手段將CGTase進行異源表達[10]。
目前,只有β-CD實現了工業化生產,但其原料的轉化率局限在50%~60%,其他2種環糊精的轉化率則更低[11]。本研究將來自于PaenibacilluscampinasensisSK13. 001的β-CGTase基因連接到大腸桿菌表達載體pET-22b(+),獲得重組質粒pET-22b(+)/β-CGT,將其轉化到EscherichiacoliBL21(DE3)中,并進行搖瓶發酵,測定菌體沉淀中β-CGTase的環化活力,并對該酶轉化淀粉生產環糊精的工藝條件進行優化,獲得了高效生產環糊精的工藝條件。
1 材料與方法
1.1 菌種與質粒
PaenibacilluscampinasensisSK13.001,保藏于本實驗室;克隆宿主大腸桿菌(Escherichiacoli)DH5α、表達宿主菌EscherichiacoliBL21(DE3),生工生物工程(上海)股份有限公司;表達質粒pET-22b(+),本實驗室保藏。
1.2 試劑及儀器
可溶性淀粉、玉米淀粉、馬鈴薯淀粉、木薯淀粉、酵母提取物,胰蛋白胨,國藥集團;基因組DNA提取試劑盒、DNA膠回收試劑盒、質粒抽提試劑盒,生工生物工程(上海)股份有限公司;PCR引物,由金唯智(蘇州)生物科技有限公司合成;工具酶,TaKaRa(大連)生物有限公司;標準α,β,γ-環糊精,Sigma(上海)公司;其他試劑均為國產分析純。
A360紫外可見分光光度計,北京普析通用儀器有限責任公司;HS-800D電熱恒溫水浴鍋,上海浦東物理光學儀器廠;GL-10MD高速臺式大容量離心機,湖南湘儀集團;高速冷凍離心機,美國Eppendorf有限公司;e2695高效液相色譜儀、RID-10A示差折光檢測器,美國沃特世公司;Agilent ZORBAX NH2色譜柱(5 μm, 4.2 mm×250 mm),美國安捷倫公司;蛋白電泳儀、核酸電泳儀、PCR儀,德國伯樂公司產品。
1.3 培養基
LB液體培養基(g/L):酵母提取物5,胰蛋白胨10,NaCl 10;LB固體培養基為LB液體培養基添加10~15 g/L瓊脂粉。
TB培養基(g/L):甘油4,胰蛋白胨12,酵母提取物24,K2HPO4·3H2O 16.4,KH2PO42.3。
1.4 實驗方法
1.4.1 重組質粒pET-22b(+)的構建與篩選
提取PaenibacilluscampinasensisSK13.001的全基因組DNA,并以此為模板設計引物PCR得到β-CGTase(包括信號肽)目的片段。PCR擴增條件:95 ℃ 3 min,95 ℃ 15 s,55 ℃ 15 s,72 ℃ 120 s,循環34次,72 ℃ 5 min;PCR引物如下:F1(5′-3′): CGCCCATGGATGAAAAGATTTATGAAACTAACAGCCG(正向引物),R1(5′-3′):CCCTCGAGAGGCTGCCAGTTCACATT(反向引物)。2條引物均在酶切位點前加入保護堿基以方便質粒構建。F1,R1分別含有NcoI和XhoI的限制性酶切位點;以F1,R1為引物PCR擴增得到β-CGTase目的片段,將其純化、回收后采用酶切位點雙酶切、酶連的方法連接到含有6個組氨酸標簽的大腸桿菌表達載體pET-22b(+)中,構建重組質粒pET-22b(+)/β-CGT,轉化到克隆宿主大腸桿菌DH5α感受態細胞中,涂布于LB固體培養基上,篩選陽性克隆子并接種LB液體培養基,37 ℃,200 r/min過夜培養。提取質粒pET-22b(+)/β-CGT進行PCR雙酶切驗證和測序鑒定。
1.4.2 β-CGTase的誘導表達及鑒定
將驗證成功的重組質粒pET-22b(+)/β-CGT轉化到E.coliBL21(DE3)感受態細胞中,pET-22b(+)空質粒處理同上作為對照,涂布LB固體培養基篩選陽性克隆子。將鑒定正確的單菌落接種于LB液體培養基中,添加適量的氨芐青霉素,200 r/min,37 ℃培養12 h,以2%接種量接種于50 mL TB培養基中,加入適量的氨芐青霉素,37 ℃,200 r/min,培養2 h后,加入終濃度為0.9 mmol/L的異丙基-β-D-硫代半乳糖苷(IPTG),25 ℃誘導培養34 h,離心收集菌體,加入裂解液超聲破碎后收集上清,上清液經過聚丙烯酰胺凝膠電泳(sodium dodecyl sulfate polyacrylamide gel electroresis,SDS-PAGE)進行蛋白表達鑒定。SDS-PAGE參照LAEMMLI[12]的方法。
1.4.3 β-CGTase的分離純化
將菌體沉淀破碎,離心得到的上清液即為粗酶液。粗酶液參照七海生物Ni-NTA親和層析柱說明書進一步分離純化,得到純化的β-CGTase,將純化后的蛋白經過SDS-PAGE進行蛋白表達鑒定。
1.4.4 環糊精葡萄糖基轉移酶環化活力的測定及轉化率的計算
β-CGTase活性主要指環化活力。測定方法為用100 mmol/L磷酸鹽緩沖液(pH 7.0)配制質量濃度10 g/L淀粉,加熱糊化,降溫至55 ℃,加入適量的粗酶液,反應10 min,煮沸滅酶,12 000 r/min離心5 min,上清液過0.22 μm濾膜,高效液相色譜法(high performance liquid chromatography,HPLC)測定環糊精含量。單位酶活力(U)定義為1 min生成1 μmol環糊精所需要的酶量。
HPLC色譜條件:Agilent ZORBAX NH2色譜柱;示差折光檢測器(RID-10A);流動相為體積分數為65%的乙腈水溶液;柱溫30 ℃;流速1 mL/min;進樣量10 μL。
環糊精轉化率如公式(1)所示:
(1)
1.4.5 重組β-CGTase反應條件優化
1.4.5.1 底物種類對環糊精產量的影響
用100 mmol/L磷酸鹽緩沖液(pH 7.0)配制不同種類的質量濃度為30 g/L的底物,包括玉米淀粉、馬鈴薯淀粉、可溶性淀粉、木薯淀粉,將底物溶液煮沸糊化并不斷攪拌,降溫后加入3 U/g干淀粉的酶液,55 ℃反應8 h后,沸水浴滅酶,反應液12 000 r/min,離心5 min,上清液過0.22 μm濾膜,HPLC檢測產物含量并計算轉化率。
1.4.5.2 底物質量濃度對環糊精產量的影響
用100 mmol/L磷酸鹽緩沖液(pH 7. 0)配制質量濃度為30、50、70、100 g/L的玉米淀粉,將溶液煮沸糊化并不斷攪拌,降溫后加入3 U/g干淀粉的酶液,55 ℃反應8 h后,沸水浴滅酶,樣品處理同 1.4.5.1。
1.4.5.3 加酶量對環糊精產量的影響
用100 mmol/L磷酸鹽緩沖液(pH 7.0)配制質量濃度為30 g/L的玉米淀粉,將淀粉溶液煮沸糊化并不斷攪拌,降溫后分別加入1、3、5、7、10 U/g淀粉的酶液,55 ℃反應8 h后,沸水浴滅酶,樣品處理同1.4.5.1。
1.4.5.4 反應時間對環糊精產量的影響
用100 mmol/L磷酸鹽緩沖液(pH 7.0)配制3%的玉米淀粉,將淀粉溶液煮沸糊化并不斷攪拌,降溫后加入5 U/g干淀粉的酶液,55 ℃下反應,間隔時間取樣,沸水浴滅酶,樣品處理同1.4.5.1。
2 結果與分析
2.1 重組大腸桿菌BL21/pET-CGT的構建與表達
以來源于PaenibacilluscampinasensisSK13. 001的基因組DNA為模板,PCR擴增得到成熟的CGTase片段,如圖1-A所示,長度大小為2 kb左右,將CGTase基因片段采用酶切位點雙酶切、酶連的方法連接到表達載體pET-22b(+)中,轉化克隆宿主大腸桿菌DH5α感受態細胞中,挑選陽性克隆子,提質粒進行雙酶切鑒定,結果如圖1-B所示,重組質粒經NcoI和XhoI兩個酶切位點進行雙酶切驗證,顯示2條明顯的條帶,條帶大小與預期相符,證明β-CGTase基因成功連接到表達載體pET-22b(+)上。
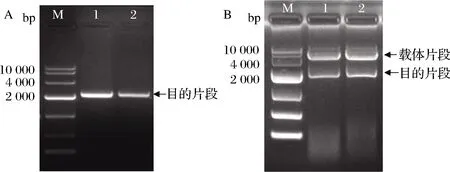
A-基因組DNA的PCR產物;B-重組質粒的雙酶切驗證
圖1 目的片段的PCR結果及重組質粒的酶切驗證
Fig.1 PCR results of target fragments and verification of recombinant plasmid digestion
將連接成功的重組質粒轉化大腸桿菌表達宿主BL21(DE3)后,進行搖瓶發酵培養,檢測發酵液上清液以及菌體沉淀中的酶活,發現上清液中環化活性較低,菌體沉淀破碎后上清液有較高酶活,從圖2可以看出隨著發酵時間的延長,β-CGTase的環化活力逐漸升高,當誘導培養時間為34 h時,環化活力達到15 U/mL,所以誘導培養時間為34 h。
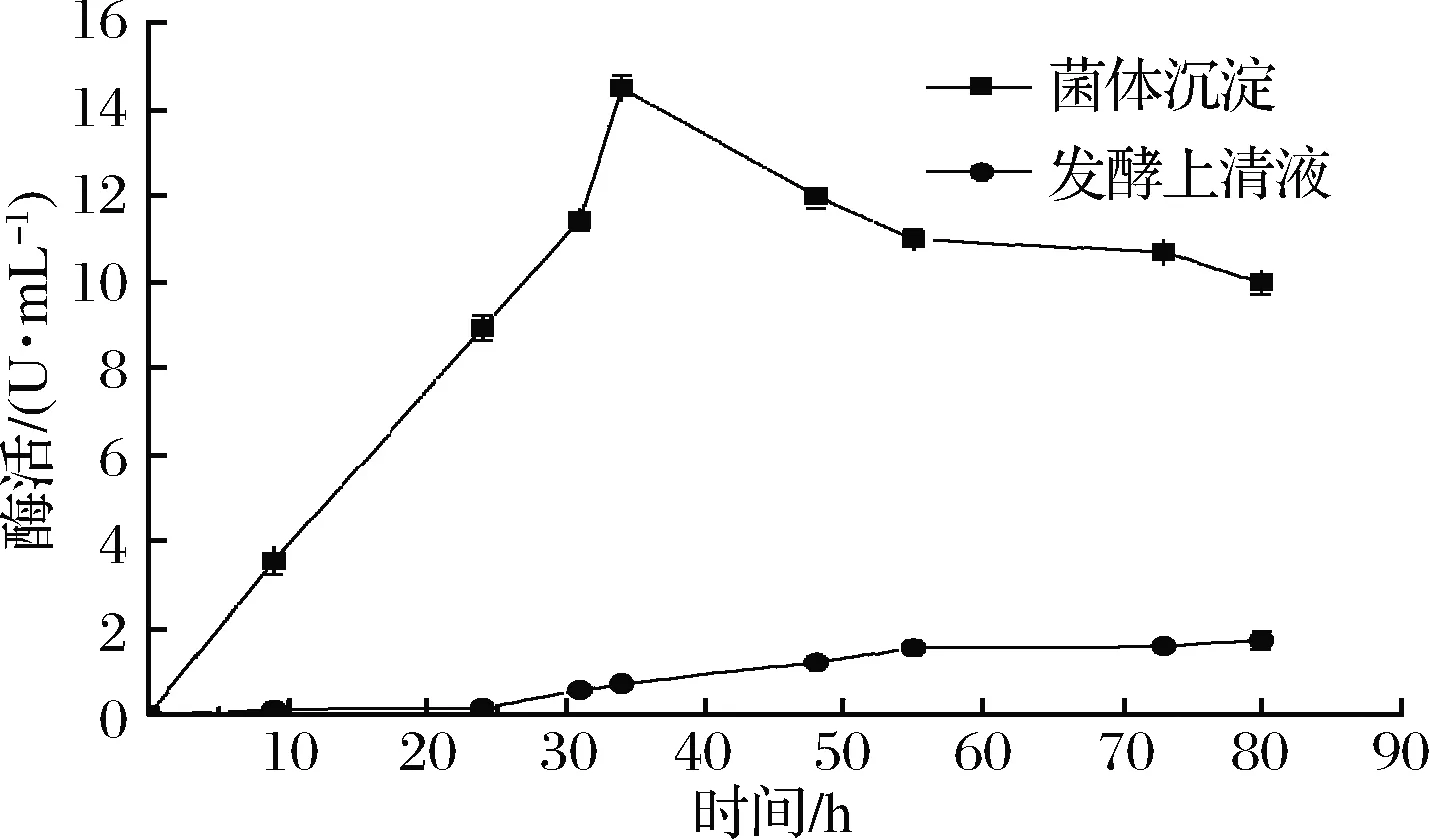
圖2 重組菌酶活隨時間變化
Fig.2 Recombinant enzyme activity changes over time
圖3為重組大腸桿菌BL21/pET-CGT蛋白表達的SDS-PAGE圖,觀察到目的蛋白條帶,預測目的蛋白分子質量為70 kDa。大多數報道的CGT酶是單體蛋白,分子質量在33~110 kDa[13];本實驗得到的CGT酶的分子質量與同物種的其他菌株分子質量相似[14]。
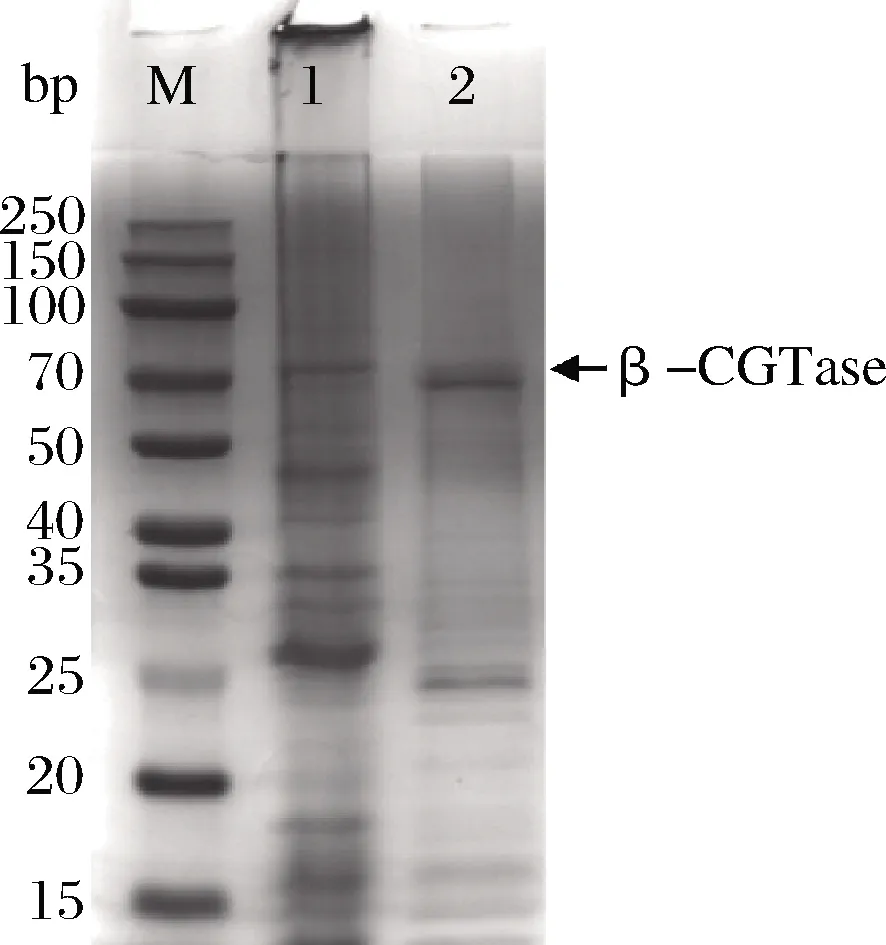
M-蛋白Marker;1-發酵胞內上清液蛋白條帶;2-純化后的蛋白
圖3 重組菌產酶SDS-PAGE
Fig.3 Recombinant bacteria producing SDS-PAGE
2.2 重組β-CGTase反應條件優化
2.2.1 底物種類對環糊精產量的影響
由圖4可知,不同淀粉類型對環糊精的轉化率有所不同。當反應作用底物為玉米淀粉時,環糊精的產量及轉化率最高為36%,馬鈴薯淀粉、可溶性淀粉、木薯淀粉分別為33%、30%、28%。產物中3種環糊精的比例為α∶β∶γ=7∶75∶18。酶法生產環糊精通常以淀粉作為底物,淀粉由直鏈和支鏈淀粉組成,不同種類的淀粉中直鏈和支鏈淀粉的比例有所不同,所以環糊精的轉化率不盡相同,在實際生產中根據不同的情況選擇合適的底物[15]。

圖4 不同種類淀粉對環糊精產量的影響
Fig.4 Effect of different types of starch on cyclodextrin yield
2.2.2 底物質量濃度對環糊精產量的影響
由圖5可知,底物濃度對環糊精的轉化率有很大的影響,隨著底物質量濃度的升高,環糊精的轉化率逐漸下降。當玉米淀粉質量濃度為30 g/L,環糊精的轉化率最高為36%;隨著底物質量分數升高,環糊精的轉化率迅速下降。造成這一現象的原因可能有:隨著底物質量分數的升高,加熱糊化后淀粉的黏度變大,使得底物與酶蛋白無法充分的接觸,導致反應速率下降;其次是酶與底物的結合逐漸達到飽和,較高的底物濃度對酶與底物的反應不會有促進作用,此外,較高的底物濃度可能會使酶反應所得產物中小分子糖含量升高,而這些小分子糖會抑制CGTase的環化作用,而增強其耦合作用,從而降低環糊精的轉化率[9]。

圖5 底物濃度對環糊精產量的影響
Fig.5 Effect of substrate concentration on cyclodextrin yield
2.2.3 加酶量對環糊精產量的影響
由表1可知,環糊精的轉化率隨加酶量的升高呈現先升高后下降的趨勢,當CGTase的添加量為5 U/g淀粉時,環糊精的轉化率最高為45.63%,其中β-CD的轉化率為35%,γ-CD轉化率為8.4%。隨著加酶量的增加3種環糊精的比例有所改變,α-CD的含量逐漸增加,β-CD、γ-CD的含量呈現先增加后降低的趨勢。隨著加酶量的增加,環糊精的轉化率并沒有增加,可能的原因是CGT酶是一種多功能酶,可以催化4種反應,其中歧化反應和耦合反應可以降解淀粉產生小分子糖,隨著加酶量的增加,這2種反應強度增強,小分子糖增多,而小分子糖的存在會使得該酶的耦合反應加劇,使環糊精解環,從而降低環糊精的產量[16]。

表1 加酶量對環糊精產量的影響Table 1 Effects of enzyme addition on cyclodextrin yield
注:括號內百分數代表每種產物所占百分比
2.2.4 反應時間對環糊精產量的影響
由圖6可知,環糊精的轉化率隨著反應時間的延長呈現先增加后下降的趨勢,隨著反應時間的延長,γ-CD的比例逐漸下降,α-CD的比例逐漸提高;當反應8 h時產量基本穩定,環糊精的轉化率最高為45.63%,其中β-CD的轉化率為35%,γ-CD的轉化率為8.4%。同種屬的菌株中,已發現的分泌CGT酶的菌株不多,其中包括Paenibacillusillinoisensis[17]、Paenibacillussp. xw-6-66[18]、Paenibacilluscampinasensissp.nov[19]以及Paenibacillusmacerans[20]等,環糊精的轉化率在30%~40%。有研究表明,隨著反應時間的延長,反應會向著產生小分子環糊精的方向移動,即α-CD的比例逐漸增加,γ-CD的比例逐漸下降[21]。該酶生產環糊精的最佳反應時間為8 h,環糊精的轉化率可達到45.63%,較少的反應時間以及較高的環糊精轉化率有利于降低生產成本,提高反應效率。
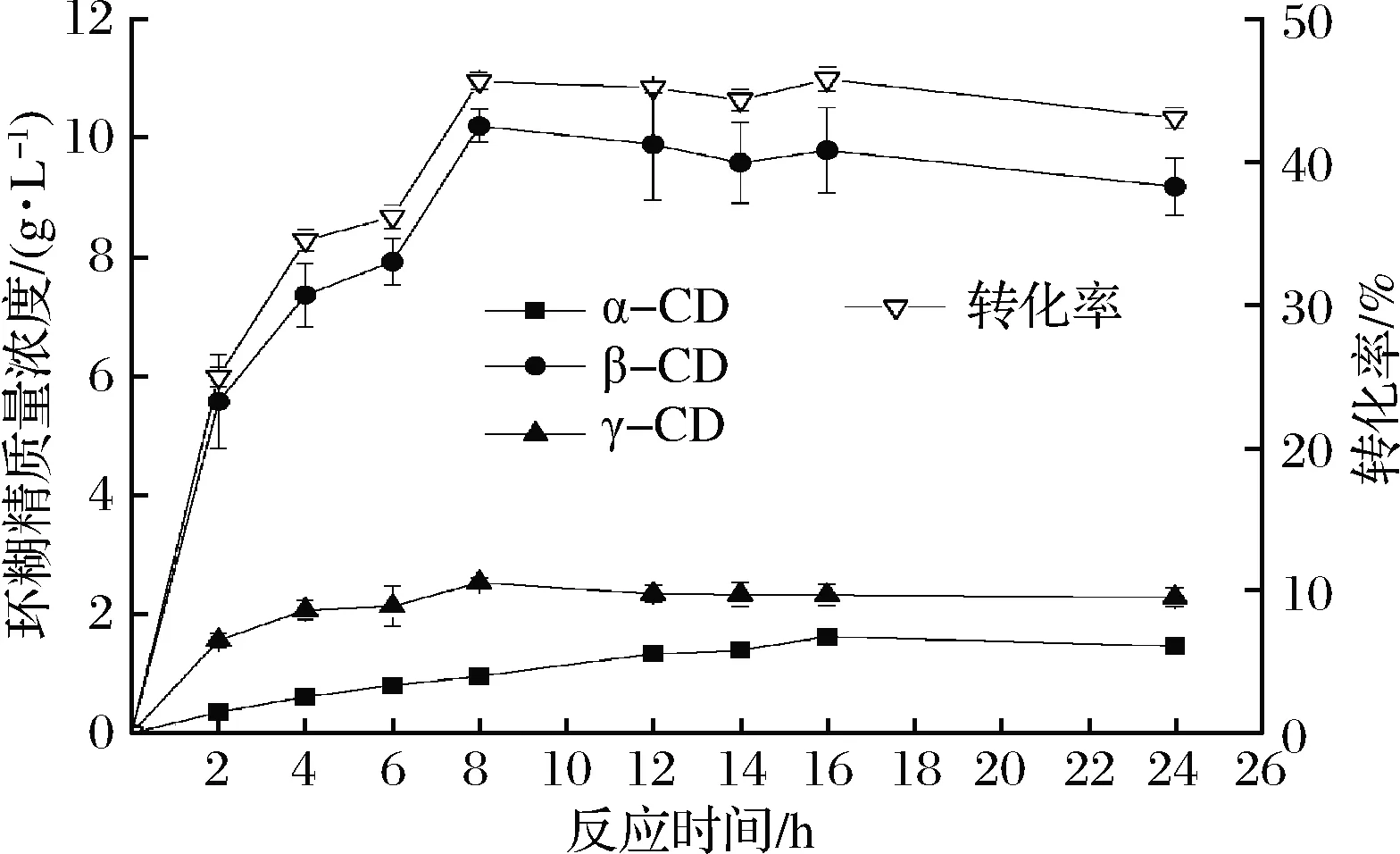
圖6 反應時間對環糊精產量的影響
Fig.6 Effect of reaction time on cyclodextrin yield
3 結論
環糊精是一種重要的環狀低聚糖,隨著其在食品、醫藥、化妝品等領域的廣泛應用,對其需求也日益增加。本文對來源于PaenibacilluscampinasensisSK13. 001的β-CGTase基因克隆到表達載體pET-22b(+)上,并轉化大腸桿菌表達宿主BL21(DE3),經過34 h的搖瓶培養,其環化活力達到15 U/mL,這為進一步實現β-CGTase的工業化生產具有重要意義。
通過對重組β-CGTase的酶反應工藝條件的優化,研究其對3種環糊精的產量、比例以及轉化率的影響。結果表明,以100 mmol/L磷酸鹽緩沖液(pH 7. 0)配制的質量濃度為30 g/L玉米淀粉,55 ℃,加酶量為5 U/g干淀粉,反應8 h;優化反應條件后環糊精的轉化率達到45.63%,其中α-CD的轉化率為2.23%,β-CD的轉化率為35%,γ-CD的轉化率為8.4%;這為環糊精的工業應用提供參考。
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
當代水產(2021年10期)2021-12-05 16:31:48
今日農業(2021年14期)2021-11-25 23:57:29
今日農業(2021年13期)2021-08-14 01:37:56
石油與天然氣地質(2021年1期)2021-02-22 14:14:44
今日農業(2020年20期)2020-11-26 06:09:10
中國果業信息(2019年10期)2019-11-13 01:21:34
中國化肥信息(2019年2期)2019-01-18 15:24:35
中國化肥信息(2019年1期)2019-01-17 21:31:12
中國化肥信息(2019年4期)2019-01-17 18:47:06
