高通量測序分析新生兒感染早產兒腸道菌群特點
2020-07-17 03:01:02孫倩姜春明
醫學綜述 2020年13期
孫倩,姜春明
(哈爾濱醫科大學附屬第一醫院新生兒重癥監護二科,哈爾濱 150001)
腸道是新生兒出生后菌群定植的主要場所,微生物的種類、分屬、數量及相對豐度等的動態平衡,在保證嬰幼兒生長發育、促進免疫系統發育成熟、參與體內物質代謝等方面起重要作用。近年來有研究表明,嬰幼兒所患疾病(感染性疾病、呼吸道疾病等)與腸道微生態的失衡有密切聯系[1]。新生兒期是腸道微生物群落建立的關鍵階段,在某些特定條件下,如機體抵抗力下降、過度使用廣譜抗生素等,腸道內的次要菌群在腸道內大量繁殖,轉變為腸道內的新優勢菌群,使得腸道穩態失衡從而致病[2]。腸道微生態的生物學功能主要是保持腸道的正常生理結構及相關功能,避免致病微生物的入侵及定植,調節人體的免疫系統發揮功能[3]。而影響新生兒腸道菌群定植的因素主要包括胎齡、分娩方式、喂養方式等。當腸道微生態失衡時,其表現的致病性包括:①導致自身感染或原定植細菌轉移到其他部位而誘發感染;②導致胃腸道功能紊亂,表現為排便困難、腹瀉等;③導致或參與某些腸道器質性病變(如壞死性小腸結腸炎)的發病[4]。因此,深入研究新生兒腸道微生態的動態變化,可針對性調整腸道內的定植菌群,有效降低新生兒感染的發病率[2]。本研究通過高通量測序分析方法對早產兒糞便樣本中所有細菌進行DNA測序,研究早產兒腸道微生態的分布特征,分析患有新生兒感染早產兒的腸道菌群特點。
1 資料與方法
1.1一般資料 選取2017年8月至2018年12月在哈爾濱醫科大學附屬第一醫院產科分娩且符合入組標準的26例早產兒作為研究對象,患兒出生后均在本院新生兒重癥監護病房住院治療。新生兒感染標準:①出生7 d出現氣促、發紺、三凹征等臨床表現。②出生7 d后血常規示白細胞計數<5×109/L,或>20×109/L;血小板計數<100×109/L;血清降鈣素原>2.0 μg/L等。③臨床癥狀為皮膚發花、四肢末梢涼、拒乳、發熱、腹脹、黃疸進行性加重等,但血培養為陰性。④血培養為陽性。按照是否感染將患兒分為新生兒感染組(12例)和未感染組(14例)。
1.2納入與排除標準 納入標準:①早產兒,胎齡為30~37周;②均為剖宮產;③出生體重為1 000~3 000 g;④住院時間超過7 d;⑤均為人工喂養;⑥收集標本前均未給予抗生素及益生菌。排除標準:①排除患嚴重消化道畸形、遺傳代謝病及重度窒息的早產兒;②排除母親孕期患有宮內感染、給予抗生素治療的早產兒。
1.3試驗方法 分別于出生后第1天(即24 h內)和出生后第7天收集患兒糞便。采集樣本時要留取早產兒糞便的中部,每份標本重量≥200 mg,將樣本裝入無菌的EP管中,并立即放入液氮中進行凍存。
利用美國丹諾爾Denovix超微量紫外可見分光光度計DS-11和適當濃度凝膠電泳對早產兒糞便中的微生物DNA做總質檢。選擇特定16S核糖體DNA為擴增片段,合成具有特異性的引物。用稀釋后的DNA序列作為模板,使用Taq DNA多聚酶進行聚合酶鏈反應,選用適當濃度的凝膠進行電泳檢測聚合酶鏈反應,其產物利用Denovix超微量紫外可見分光光度計DS-11和凝膠電泳進行相應文庫質檢[5]。
檢驗合格后,使用熒光定量系統對文庫進行定量,并依據每個樣本的數量要求,進一步做對應要求的混合。使用美國Illumina公司的HiSeq PE2500測序平臺進行上機測序[6]。由上海銳翌基因科技有限公司完成相應樣本提取、建庫、測序、分析。
1.4生物信息學分析
1.4.1測序數據統計 對原始數據進行過濾、鏈接和去嵌合體處理,得到的有效序列按照97%相似度進行歸類,得到操作分類單元(operational taxonomical unit,OTU),每個OTU對應一個不同的細菌(微生物)種群。然后對每個樣本的序列數據進行處理,進一步提取相應的OTU序列。對每個OTU序列進行種群相應的分類,依據每個分類單元中序列的數目,獲得相應的豐度表,最終依據該豐度表進行后續相應數據分析[7]。
1.4.2物種分類和豐度分析 依據物種歸類的結果,分別在各分類等級(門、綱、目、科、屬)對各個樣本做種群相對豐度圖,可直接查看在每一等級上,物種的相對豐度及所占比例。
1.4.3物種多樣性分析 運用Beta多樣性分析,利用各樣本序列間的豐度信息來計算樣本間距離,從而反映組間是否具有顯著的微生物群落差異。采用多響應置換過程(multi-response permutation procedure,MRPP)分析組間菌群組成的差異性,P<0.05 為差異有統計學意義。MRPP分析包括加權與非加權兩種:非加權時,不考慮物種的豐度進行加權處理,會擴大或縮小物種豐度的差異;而加權統計后,考慮了物種的豐度,保留了物種原始豐度特征,對低豐度菌群反映更好。
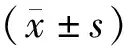
2 結 果
2.1一般情況 26例早產兒共收集52份糞便樣本,成功提取DNA并完成測序25份,將標本信息納入生物信息分析。進入測序分析的兩組早產兒的胎齡、出生體重比較差異無統計學意義(P>0.05),見表1。
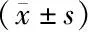
表1 兩組早產兒的胎齡和出生體重比較
2.2測序數據結果和OUT統計 本次測序得到的有效序列總數為828 388,總堿基數為351 852 086 bp,平均長度為424.35 bp,其中長度為420~440 bp的優質序列占總序列的92.68%。25個標本共得到79個OTU,說明本次測序的深度足夠,測序所得數據量合理。
2.3物種多樣性的分析
2.3.1單個樣品的菌群分類及相對豐度 共測得門水平6個、綱水平12個、目水平17個、科水平29個、屬水平40個,菌群種類共40個。豐度較高的菌群包括門水平為變形菌門、擬桿菌門、厚壁菌門、放線菌門,綱水平為γ-變形菌綱、擬桿菌綱、桿菌綱、梭菌綱、放線菌,目水平為腸桿菌目、擬桿菌目、乳酸桿菌,科水平為腸桿菌科、擬桿菌科、紫單胞菌科、腸球菌科、消化鏈球菌科,屬水平為克雷伯桿菌屬、埃希桿菌屬/志賀桿菌屬、擬桿菌屬、副桿菌屬、腸球菌屬,見圖1~5。
2.3.2早產兒生后第7天腸道菌群對新生兒感染情況的影響 運用物種MRPP分析提示,新生兒感染組與非感染組的腸道微生物群落結構差異有統計學意義(P<0.05),見表2。
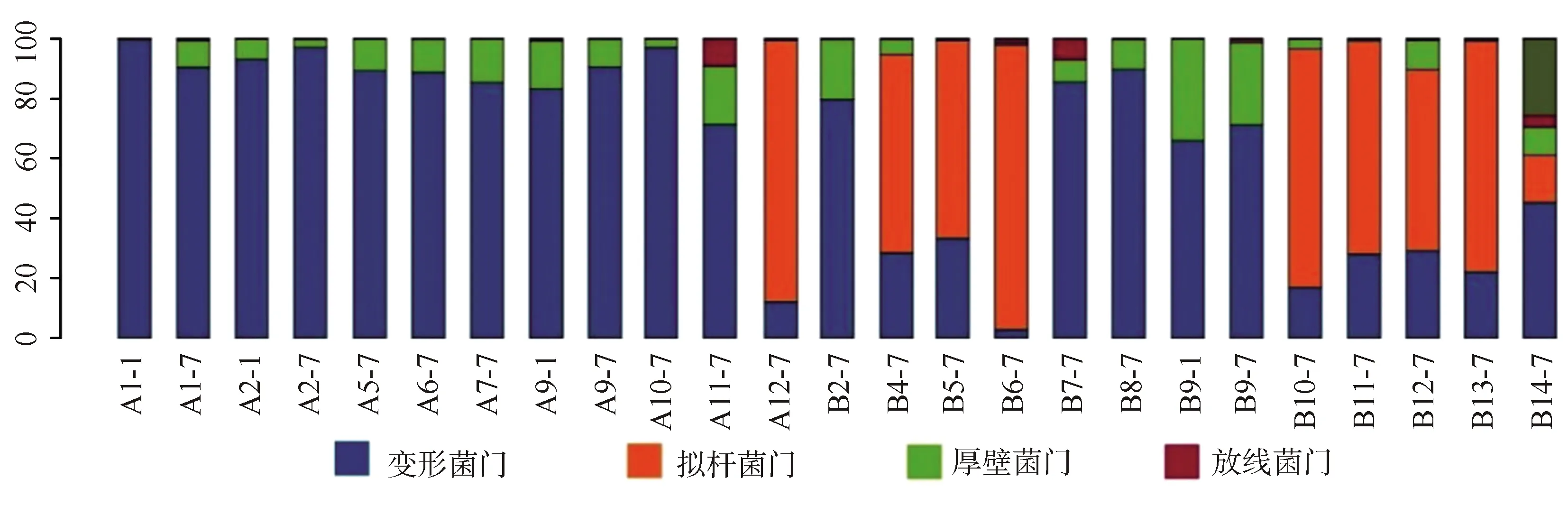
A:感染組;B:非感染組
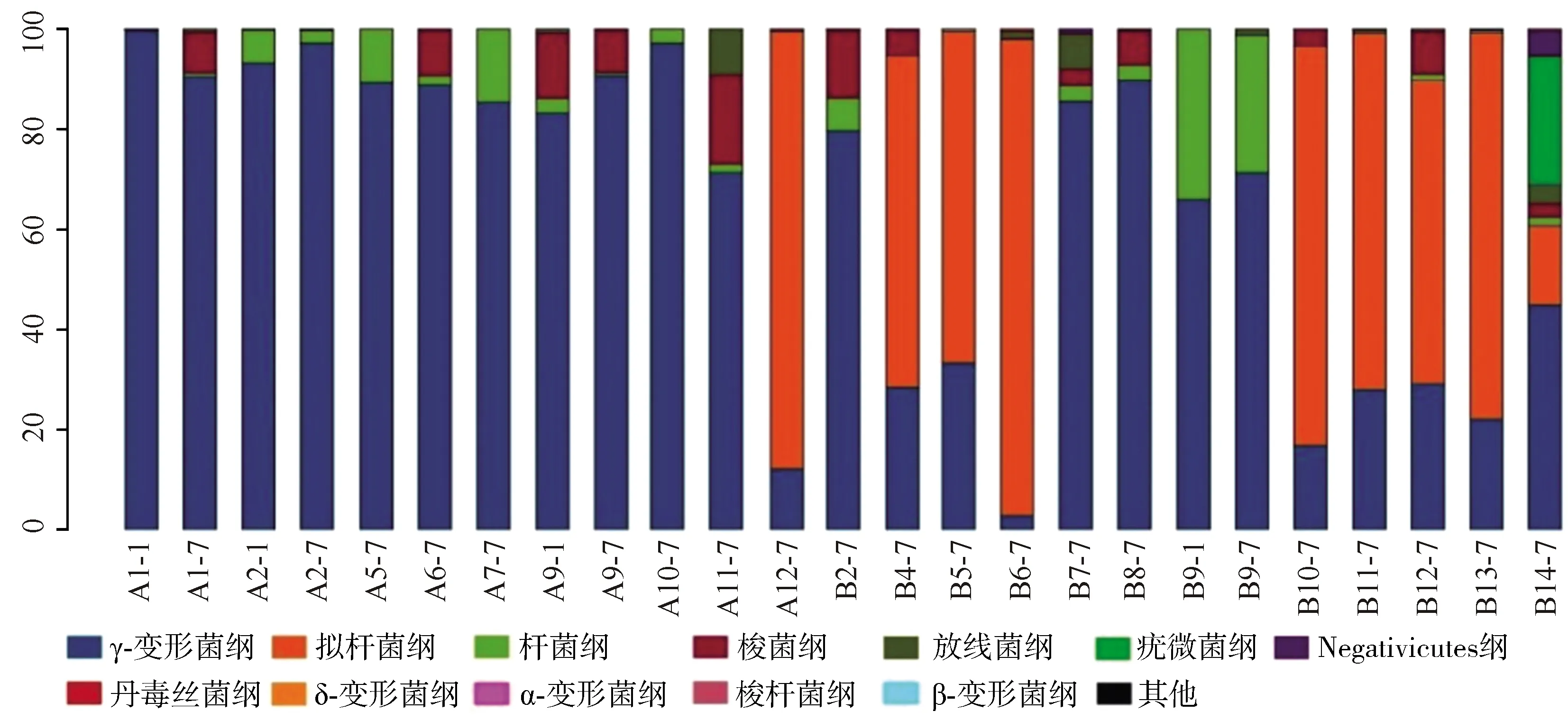
A:感染組;B:非感染組
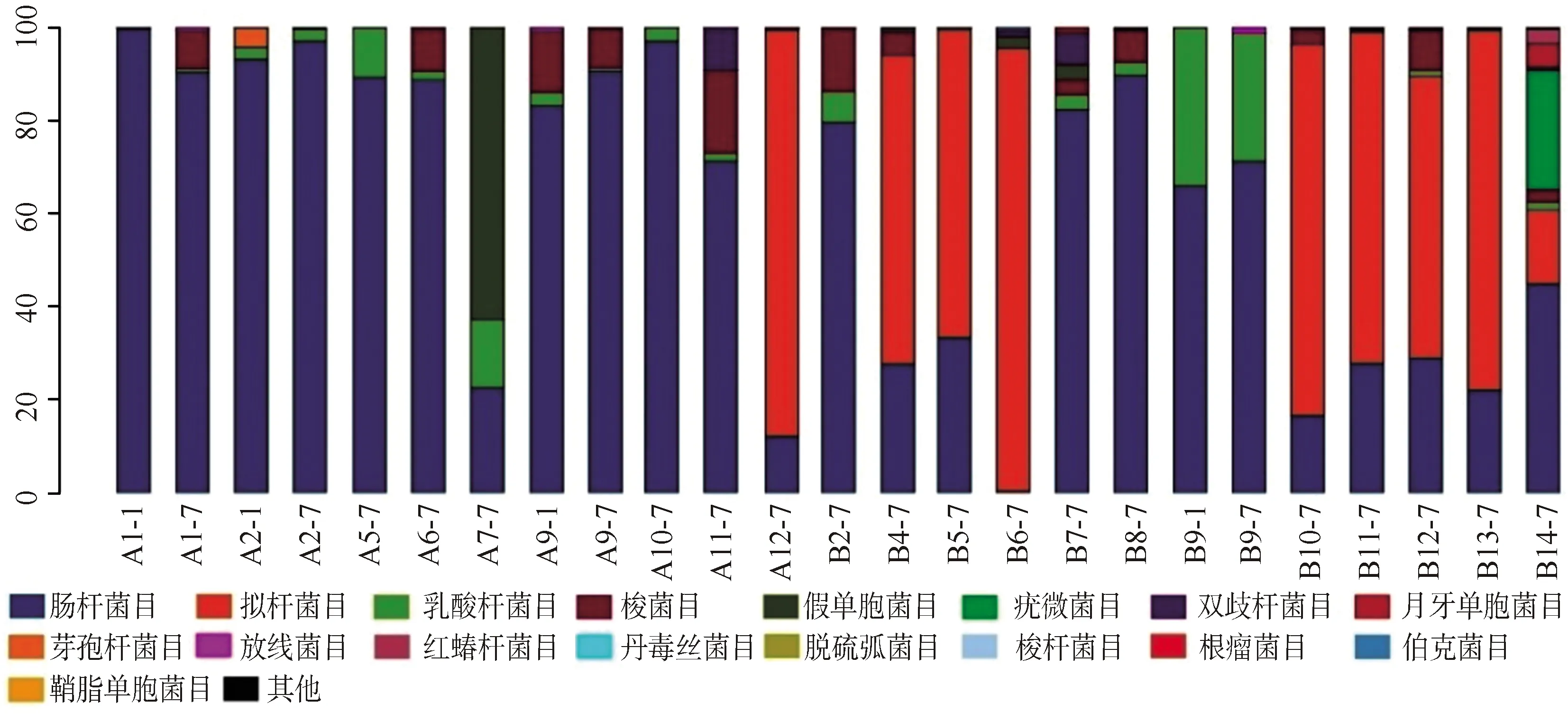
A:感染組;B:非感染組

A:感染組;B:非感染組
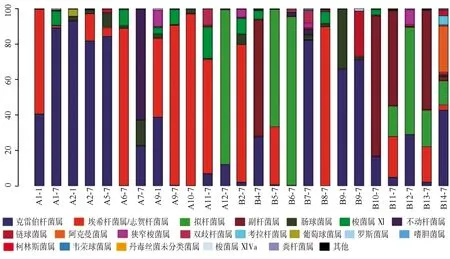
A:感染組;B:非感染組

表2 兩組早產兒的MRPP差異比較
3 討 論
新生兒腸道微生態動態平衡是維持新生兒正常生長發育的重要因素,影響新生兒腸道菌群分布的因素較多,主要包括胎齡、分娩方式、喂養方式、新生兒抗生素使用情況等。當腸道微生物群失衡時,極易誘發新生兒感染性疾病,常見的有宮內感染、敗血癥、壞死性小腸結腸炎等。
有研究表明,母乳喂養以雙歧桿菌、乳酸桿菌為優勢菌群,且可以為有益菌群的定植提供合適的環境[8];陰道分娩的胎兒腸道菌群多樣性較高,且無定植延遲,較少發生腸道菌群失調并引發感染性疾病[9]。因此,本研究選取剖宮產出生、人工喂養的早產兒作為研究對象,以減少分娩方式及喂養方式對腸道菌群的影響。本研究共收集了52份新生兒糞便樣本,DNA提取失敗共27份,其中22份出生后第1天及5份出生后第7天糞便標本因濃度過低未提取成功。由于新生兒出生時腸道內接近無菌狀態,出生后才逐漸獲得各種菌群定植,所以本研究中患兒生后第1天糞便樣本DNA提取成功率較低。既往研究表明,胎齡是新生兒腸道微生態形成的影響因素,早產兒身體各器官發育均不成熟,抵抗力與適應性差,故出生體重越輕,胎齡越小,腸道成熟度越低,腸道菌群的豐富度及多樣性越差,各種感染的發生率越高[10]。本研究對象是胎齡為30+2~36+3周,平均胎齡為34 周的早產兒,因此糞便的微生物含量可能較低,影響DNA的提取。
本研究將提取成功的25份樣本的基因進入測序程序,由Alpha稀釋曲線可知,每一份樣本的曲線均隨Reads數目的增加而趨于平坦,所以本試驗測序深度足夠,且測序數據量比較合理。本研究測得的早產兒早期腸道菌群,按細菌分類表分類包括門水平6個、綱水平12個、目水平17個、科水平29個、屬水平40個。研究表明,由于早產兒胃腸道在宮內發育不成熟、出生后腸道由宮內的低氧環境轉為宮外富氧環境、腸道抗氧化系統未成熟等,早產兒的腸道微生態形成延遲,所以其豐度較足月新生兒明顯降低[11]。
本研究對感染與非感染狀態下早產兒生后第7天的腸道菌群分布進行比較,其中新生兒感染組共測得門水平4個、綱水平6個、目水平10個、科水平13個、屬水平35個,未感染組共測得門水平5個、綱水平10個、目水平14個、科水平28個、屬水平40個,未感染組腸道菌群的豐富度明顯高于新生兒感染組。經MRPP組間差異分析,兩組細菌物種的組成差異有統計學意義。通過主坐標分析對各樣品菌種的相對豐度進行分析發現,與新生兒感染組相比,未感染組的新生兒腸道菌群相對豐度更高,腸道微生態多樣性更加豐富。
本研究結果顯示,新生兒感染組的腸道菌群主要分布如下:門水平為變形菌門,綱水平為γ-變形菌綱、桿菌綱,目水平為腸桿菌目、梭菌目,科水平為腸桿菌科、腸球菌科,屬水平為克雷伯桿菌屬、大腸埃希菌屬/志賀菌屬及擬桿菌屬;未感染組的腸道菌群主要分布如下:門水平為擬桿菌門、變形菌門,綱水平為γ-變形菌綱、擬桿菌綱、桿菌綱,目水平為腸桿菌目、擬桿菌目、乳酸桿菌目,科水平為腸桿菌科、擬桿菌科、紫單胞菌科,屬水平為克雷伯桿菌屬、大腸埃希菌屬/志賀菌屬、擬桿菌屬、副桿菌屬。可見,新生兒感染組菌群分布范圍較小、菌種含量較少、變形桿菌豐度較高;而未感染組的菌群分布更為廣泛、多樣性較高、更有均一性。但是菌群多樣性過高同樣也會增加條件致病菌的致病機會,所以恰當的菌群多樣性才可以對宿主起保護作用[12]。
新生兒感染可發生在三個階段:①出生前感染,可發生于妊娠期各階段,最常見的途徑為病原菌經母親血流通過胎盤感染胎兒,又稱宮內感染;②出生時感染,剖宮產、胎膜早破、產程延長等均可導致胎兒感染;③出生后感染,較上述兩種感染更為常見。病原體通過呼吸道、消化道等途徑傳播,導致新生兒敗血癥、新生兒感染性肺炎等。而新生兒腸道微生態的建立是一個緩慢發展的動態演變過程,直至2周歲時才逐漸趨于穩定,且與正常成人的腸道微生態相似[13]。針對早產兒細菌培養的研究已發現,早產兒腸道細菌定植延遲,其潛在條件致病菌的優勢菌群包括腸桿菌、擬桿菌及已知病原體(梭菌、葡萄球菌、假單胞菌和克雷伯菌)[14]。早產兒腸黏膜屏障功能并不成熟,造成腸壁通透性增加,故可能導致腸道微生態中的病毒、細菌和內毒素進入腸黏膜上皮細胞,造成腸道感染,甚至可以穿過腸壁進入血液、器官或組織,造成全身感染[15]。
感染發生及其嚴重程度與致病微生物的毒性有極大關系。感染是否發生,一方面依據細菌的數量、感染方式、毒力等,另一方面與機體當時的免疫狀態也有極大關系。有研究表明,腸道內腸桿菌科的大腸埃希菌可以在極低水平時造成感染,當其內部的Rcs系統啟動時,細菌對宿主細胞的侵襲力增強,致使感染加重;同屬腸桿菌科的克雷伯桿菌亦是如此,菌內的Rcs系統啟動使得莢膜多糖合成,進而產生高毒力,易導致敗血癥、傳染病鼠疫、肺炎等感染性疾病的發生[16]。所以,當腸道內定植大量變形桿菌(腸桿菌)時,條件致病菌群的相對豐度增加,可造成腸道微生態失衡,易誘發新生兒感染。當腸道內微生物多樣性增加時,致病菌的相對豐度就會降低,且與其他菌群形成相對穩態,降低了感染性疾病的發生率。此外,包含敗血癥和全身炎癥反應綜合征在內的嚴重新生兒感染均與微生物定植-宿主相互作用有關。有研究表明,新生兒敗血癥可能是由腸黏膜防御功能低下,通透性高,腸道細菌移位侵入血液循環引起[17-19]。由于早產兒開奶延遲或不能母乳喂養,雙歧桿菌、乳酸桿菌、擬桿菌等益生菌無法在腸道內正常定植,進而導致病原菌在腸道內定植或作為優勢菌群在腸道內進行大量繁殖,侵襲腸道,引起腸黏膜損傷,最終導致壞死性小腸結腸炎[20]。
本研究結果顯示,腸道內定植大量變形桿菌(腸桿菌),增加了菌群的相對豐度,由次要菌群變為優勢菌群,同時降低了有益菌群的相對豐度,造成腸道生理穩態的失調,極易誘發新生兒感染。當腸道定植菌群的多樣性增加時,可以維持腸道菌群的動態平衡,促進了免疫系統的建立和成熟,降低新生兒尤其是早產兒感染性疾病的發生率。
綜上所述,關于新生兒感染時早產兒腸道菌群的分布仍需大量試驗驗證,目前腸道微生態與新生兒疾病的關系已成為研究熱點。隨著試驗技術及設備的不斷進步,基因庫的不斷完善,新生兒腸道微生物群落的變化在新生兒感染性疾病發生中所起的作用將得到充分證實,同時腸道微生態也會對新生兒相關疾病的早期診斷和治療起到一定作用,極大改善新生兒,尤其是早產兒感染性疾病的預后并對遠期生長發育產生影響。
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
保健醫苑(2021年7期)2021-08-13 08:48:02
學生天地(2020年36期)2020-06-09 03:12:30
小學科學(學生版)(2020年5期)2020-05-25 07:11:32
家庭醫學(下半月)(2019年9期)2019-10-12 08:04:06
家庭醫學(下半月)(2019年8期)2019-09-25 09:02:00
人大建設(2019年12期)2019-05-21 02:55:32
媽媽寶寶(2017年3期)2017-02-21 01:22:12
領導文萃(2015年4期)2015-02-28 09:19:05
中國衛生標準管理(2015年8期)2015-01-26 18:08:35
