楨楠幼樹光合特性對鎘脅迫的響應
2020-07-17 10:25:50鐵得祥胡紅玲喻秀艷舒奕嘉
生態學報 2020年11期
關鍵詞:植物
鐵得祥,胡紅玲,*,喻秀艷,2,舒奕嘉,張 健
1 四川農業大學林學院, 成都 611130 2 自貢市林業重點工程管理中心,自貢 643000
植物的光合作用對重金屬較為敏感[1],對于非耐鎘植物而言,光合合色素、氣體交換參數和葉綠素熒光參數等光合特性指標在Cd脅迫下均會呈現出顯著降低趨勢[2],其原因主要有以下幾方面:Cd會阻礙葉綠素的生物合成[3]、破壞光合器官的結構和功能,顯著降低植物的光合作用速率;還能干擾電子轉移和ATP的合成過程、抑制暗反應階段的關鍵酶活性[4- 5];此外,也能通過改變植物葉肉細胞的水勢和膨壓,來控制植物的氣孔行為[6],進而降低植物的氣體交換速率,使得部分氣體交換參數值下降。但對與耐Cd植物而言,較低濃度鎘脅迫下,植物的光合作用并不會受到顯著抑制,如寶山堇菜(Violabaoshanensis),龍葵(Solanumnigrum)等植物[7- 8];由此可見,Cd對植物光合作用的影響程度大小與植物對Cd的耐受程度密切相關。此外,不同耐Cd植物對鎘脅迫的光合特性響應也存在明顯差異,如龍葵和印度芥菜 (Brassicajuncea)的葉綠素含量和凈光合速率在較低Cd濃度脅迫與正常生長環境下并無顯著差異,但部分氣體交換參數值(蒸騰速率和氣孔導度)卻呈現出相反的變化趨勢[7,9];再如球果蔊菜(Rorippaglobosa)的葉綠素含量隨Cd脅迫程度的加重呈先升后降趨勢[10],而龍葵的葉綠素含量則呈逐漸下降的趨勢[7]。綜上表明,不同植物的光合特性對Cd脅迫的響應存在很大差異。
楨楠(Phoebezhennan)屬樟科(Lauraceae)楠屬(Phoebe)植物,是我國特有的珍稀樹種,在我國主要分布于四川盆地、貴州北部及湖北地區。該樹種多為高大的常綠喬木,生命周期長、幼齡期生長快,適應性較強,目前在綠化工程中廣泛應用[11]。也因其特殊的材質、紋理特征和醫藥成分,在林業、輕工業、醫藥等行業都具有較高的經濟價值[12]。目前對于楨楠光合特性的研究多集中在干旱、施肥、水淹脅迫等方面[13- 14],但都未從葉綠素熒光層面去進行更加深入的探究,作為植物光合作用與環境關系的內在探針[15],通過分析楨楠幼樹體內葉綠素熒光參數和光合作用參數的相互關系,能更加全面地了解楨楠幼樹的光合作用機理,如植物葉片的電子傳遞、轉移過程,對吸收光能的利用情況等[16- 17]。目前由于工業排放和農藥化肥的大量施用,重金屬污染風險逐步加重,尤其是在長江中下游地區Cd污染日趨嚴重,因此探討楨楠幼樹光合生理和葉綠素熒光對不同濃度Cd脅迫的響應情況,有助于更加全面的了解楨楠幼樹對Cd的基本適應能力,為深入研究楨楠抗Cd脅迫機制提供理論依據,也可為選擇適宜的楨楠栽培立地提供參考。
1 材料與方法
1.1 試驗地概況
試驗地設在四川農業大學(成都校區)教學科研基地,地理位置為東經103°51′29″,北緯30°42′18″,屬亞熱帶濕潤氣候區,四季分明,氣候溫和,雨量充沛。年平均氣溫16.8 ℃,月平均最高溫為25.9 ℃(7月),月平均最低溫為6.9 ℃(12月),極端最高氣溫38 ℃(7月),極端最低氣溫-2 ℃(1月);累年平均相對濕度達84%;全年平均降水量為759.1—1155.0 mm;年平均總日照時數為840.2 h。
1.2 試驗材料
1.2.1栽植土壤與容器
試驗用土為農田沙壤土,其理化性質為:pH6.97、有機質27.51 g/kg、全氮2.31 g/kg、全鉀3.88 g/kg、汞0.01 mg/kg、砷9.91 mg/kg、鎘0.19 mg/kg、鉛3.34 mg/kg、鉻100.75 mg/kg、銅40.27 mg/kg。栽植容器均采用規格一致的塑料盆(盆口直徑30 cm,底徑20 cm,高23 cm),每盆等量裝土8.5 kg(干土質量)。
1.2.2供試植株
為同一種源的2.5 a生楨楠幼樹,無病蟲害,平均株高118.55±8.80 cm,平均地徑13.37 ±0.20 cm,每盆1株,移栽后常規水費管理,進行為期1 a的適應性培養。期間采用HH2土壤水分測定儀(ML2x,GBR)監測土壤水分狀況,使土壤體積含水量保持在20%左右(即本試驗用土田間持水量的80%)。試驗開始前15 d,將所有待處理植株移入通風秀氣的大棚內,并停止施肥,以防肥料對鎘效應的表達產生干擾。
1.3 試驗設計
采用單因素試驗設計,根據四川省土壤環境調查數據[18],四川有較多地區土壤Cd含量達4.4 mg/kg,且少數污染嚴重地段鎘含量高達22.20 mg/kg,加之近年來西南地區土壤Cd含量增幅較大[19],因此根據四川地區鎘污染現狀并結合未來可能的發展趨勢,本試驗共設置6個Cd處理水平:CK(0 mg/kg)、T1(10 mg/kg)、T2(20 mg/kg)、T3(30 mg/kg)、T4(40 mg/kg)、T5(50 mg/kg),其中Cd以CdCl2的形式水溶施入,用量見表1。為避免苗木移入塑料盆后的適應培育過程中一些植株因外界干擾生長受阻或意外死亡影響樣本選擇,每個處理重復9次,共計54盆。所有處理植株均放置在通風透氣大棚內培養。試驗于2017年6月初開始,以CdCl2與相同體積去離子水配成母液,然后稀釋成不同處理濃度。為使鎘施入方式與現實中鎘排放過程相似,本次試驗將各處理CdCl2等分為5份,分別于2017年7、9、11月,2018年1、3月施入。施入時,將配制的溶液均勻澆灌在盆內土壤表面,另用托盤墊于盆底收集可能滲出的CdCl2溶液,并將其澆回土壤中,對照澆等量的去離子水。于2018年5月初試驗結束時,每個處理抽取3株重復,在每株新梢上相同葉位選取3片成熟功能葉,采用Li- 6800便攜式光合測定儀(Li-Cor Inc.,USA)在不同時間分別測定各處理植株葉片的氣體交換參數、光響應特征曲線以及葉綠素熒光參數,之后采集功能葉,裝入密封袋,標記好之后置于冰盒中,即刻帶回實驗室進行葉綠素含量測定。

表1 重金屬Cd處理水平
1.4 測定指標及測定方法
1.4.1光合色素含量測定
采用分光光度法(80%丙酮與無水乙醇1∶1混合提取法)測定[20]。
1.4.2光合氣體交換參數測定
凈光合速率(Pn)、蒸騰速率(Tr)、氣孔導度(Gs)和胞間CO2濃度(Ci)使用Li- 6800便攜式光合測定儀(Li-Cor Inc.USA)于2018年5月一個晴天上午9:00—11:00進行測定,設定參比室CO2濃度為400 μmol/mol,葉室溫度為30 ℃,葉室相對濕度為50%,光照強度設定為環境光強(1000 μmol m-2s-1)。葉片均選用同方位成熟的功能葉,每個處理重復測定3株,每株測定3個葉片。
1.4.3光合-光響應曲線測定
使用Li- 6800進行測定,測定時間為2018年5月多個晴天上午(8:30—11:30),葉片選擇標準與數量同上。光強梯度設定為1800、1500、1200、1000、800、600、400、200、150、100、75、50、25、0 μmol m-2s-1,葉室其他參數設定與測定氣體交換參數的設定一致,通過預試驗測定楨楠幼樹的光飽和點在800 μmol m-2s-1左右,故首先將楨楠幼樹葉片在800 μmol m-2s-1的光強下進行光誘導直至Pn穩定(大約5 min),然后使用光響應曲線自動測定程序進行測定。
1.4.4熒光參數測定
使用Li- 6800進行測定,葉片選擇標準與數量同上,葉室參數設定與測定氣體交換參數的設定一致。測量前將供試植株移入無光實驗室進行暗適應,并用錫箔紙包裹待測葉片(標記測定葉片,并充分保證暗適應),待暗適應一整夜后,在第二天8:00開始熒光參數的測定,首先用一束小于0.1μmol m-2s-1的測量光(頻率為50 Hz)照射,得到初始熒光F0;后打開一個飽和脈沖光(8000 μmol m-2s-1,100 Hz,1000 ms),得到暗適應下的最大熒光Fm,然后根據環境光強設定活化光,待F穩定后,此時的F即為穩態熒光Fs;然后再打開一個同樣強度的飽和脈沖光,得到Fm′;同樣打開遠紅光(25 μmol m-2s-1)得到F0′。根據測定數據,可計算得出PSⅡ潛在活性(Fv/F0)、PSⅡ最大光化學量子產量(Fv/Fm)、PSⅡ有效光化學量子產量(Fv′/Fm′)、PSⅡ實際光化學量子產量(ΦPSII)、電子傳遞速率(ETR)、光化學淬滅系數(qP)和非光化學淬滅系數(NPQ)[21]。
1.4.5PSⅡ吸收光能的分配比例
根據Demmig|Adams等[22]提出的方法計算:
(1)光化學耗散(P)=qP×Fv′/Fm′×100%;
(2)天線熱耗散(D)=(1-Fv′/Fm′)×100%;
(3)過剩激發能(Ex)=(1-qP)×Fv′/Fm′×100%。
1.5 數據處理
使用Excel 2016軟件(Microsoft Office, USA)進行數據處理,采用SPSS 22.0對數據進行單因素方差分析(One-way ANOVA),顯著水平α設定為0.05,用Duncan′s法進行多重比較,最后用Sigmaplot 12.5(Systat Software Inc., USA)做圖。
光合-光響應曲線測定結果采用雙曲線修正模型擬合,同時獲得最大凈光合(Pmax)、光補償點(LCP)、光飽和點(LSP)和暗呼吸速率(Rd)等光合特征參數[23- 24]。
2 結果與分析
2.1 鎘脅迫對楨楠幼樹葉綠素含量的影響
如圖1所示,隨著Cd脅迫程度的加重,葉綠素a(Chla)、葉綠素b(Chlb)、類胡蘿卜素(Car)和葉綠素總量(Chla+b)均呈下降趨勢。其中Chla、Chlb、Chl(a+b)分別在T3、T2、T3時顯著下降(P<0.05),相比CK降幅分別達45%、51%、46%,Car含量在T1時顯著下降,降幅達34%。表明低濃度(T1—T2)脅迫下,Cd對Chla、Chlb、Chl(a+b)含量無明顯影響,但對Car影響顯著;而高濃度(T3—T4)Cd脅迫顯著影響了各類葉綠素含量。由圖1還可知,Chla/b隨Cd脅迫的增大逐漸上升,可見Cd對Chla的抑制程度比Chlb更弱一些。
2.2 鎘脅迫對楨楠幼樹氣體交換參數的影響
如表2所示,隨著Cd脅迫程度的加重,楨楠幼樹的Pn、Gs、Tr均呈下降趨勢,且分別在T1、T1、T2時顯著下降了24%、31%、21%。且隨Cd脅迫程度的加重而增加,Ci呈在增加趨勢,LS呈降低趨勢,并均在T4處理時顯著高于(低于)CK,表明此時楨楠幼樹葉片的固C能力減弱,且楨楠幼樹葉片光合速率的降低主要由非氣孔因素決定。
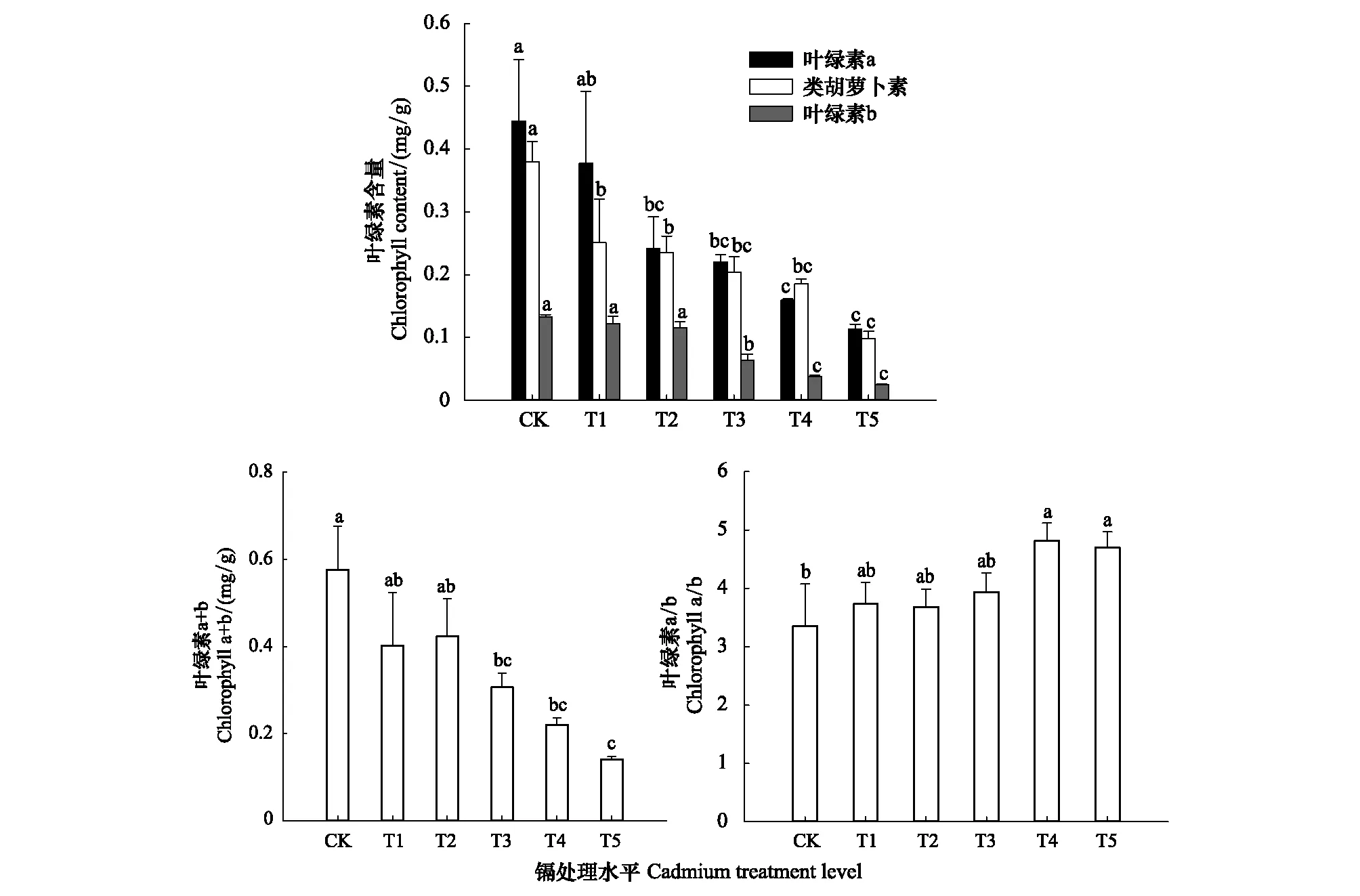
圖1 不同水平鎘處理下楨楠幼樹葉片中色素含量變化Fig.1 Changes of pigment content in leaves of P. zhennan leaves under different levels of cadmium treatment圖條上方不同小寫字母分別表示鎘處理間差異顯著(P < 0. 05)
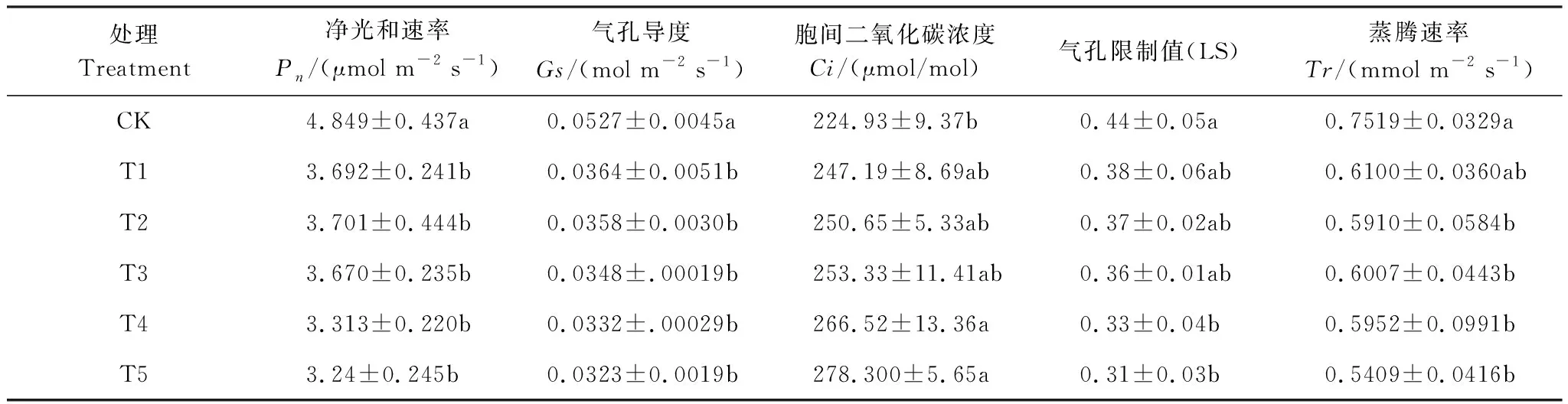
表2 不同水平Cd處理下楨楠幼樹氣體交換參數變化
2.3 鎘脅迫對楨楠幼樹葉片光合-光響應特征的影響
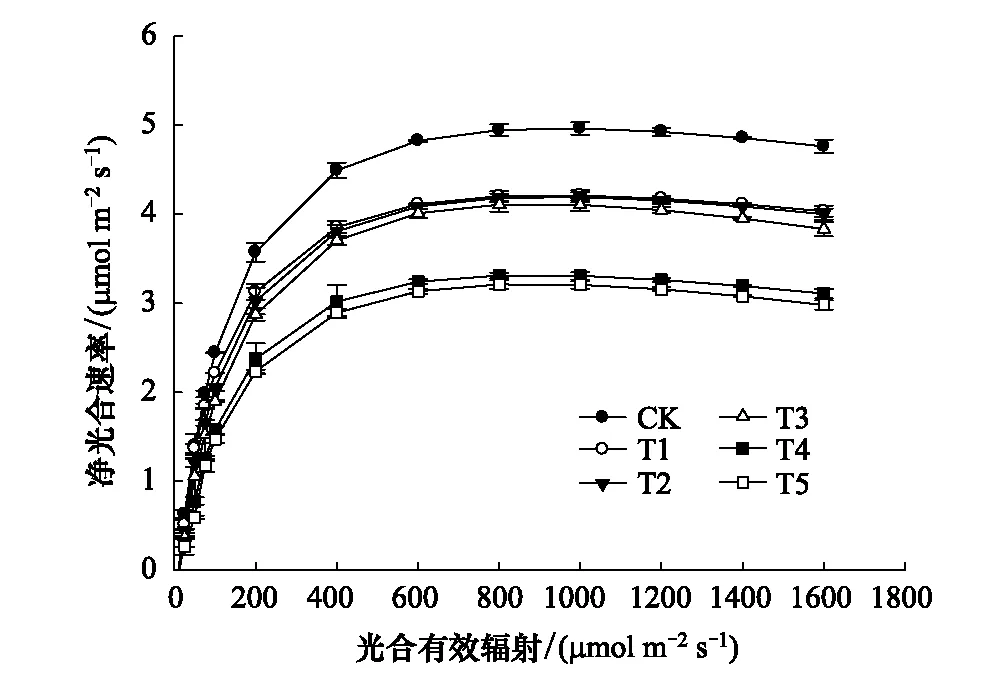
圖2 不同水平鎘處理下楨楠幼樹葉片的光合-光相響應曲線 Fig.2 Photosynthetic-optical response curves of leaves of P. zhennan var.
由圖2可知,在同一PAR下,隨著Cd脅迫程度的加重,楨楠幼樹對光能的利用率逐漸降低,最大凈光合速率(Pmax)呈下降趨勢。在同一鎘處理下,當PAR≤100 μmol m-2s-1時,Pn均隨光合有效輻射的增強而急劇增大,二者基本呈直線關系,此時PAR是限制楨楠幼樹光合作用的唯一因素;隨著PAR的持續增強,各處理植株Pn仍呈上升趨勢;當PAR≥800 μmol m-2s-1時,Pn基本趨于平緩,此時CO2濃度成為限制楨楠幼樹光合作用的唯一因素,且CK的最大Pn均顯著高于Cd處理植株(表7),T2與T3最大Pn也顯著高于T4與T5,而T2與T3,T4與T5間則無明顯差異。由表7可知,隨著Cd脅迫程度的加強,楨楠幼樹的表觀量子效率(AQY)逐漸降低,光補償點(LCP)逐漸升高,表明楨楠幼樹光合機構遭受一定程度的破壞,其對弱光的利用能力、干物質積累能力等均減弱。
2.4 鎘脅迫對楨楠幼樹葉綠素熒光參數的影響
如表4所示,Fv/Fm、Fv/F0在T1—T4處理范圍內無顯著變化,在T5處理時顯著降低;ΦPSⅡ、ETR、qP均隨Cd脅迫程度的加重呈下降趨勢,T1—T2時顯著降低;NPQ呈升高趨勢,并至T4處理時顯著高于CK。由此可知,在T1—T4處理時,楨楠幼樹葉片仍能保持正常的光能轉化效率及穩定的PSⅡ潛在光活性,但葉片PSⅡ光反應中心對Cd較為敏感,Cd脅迫下其受損明顯,開放程度降低,但楨楠幼樹也會盡力提高PSⅡ反應中心的熱耗散能力(NPQ上升),以此減輕光抑制所產生的傷害。
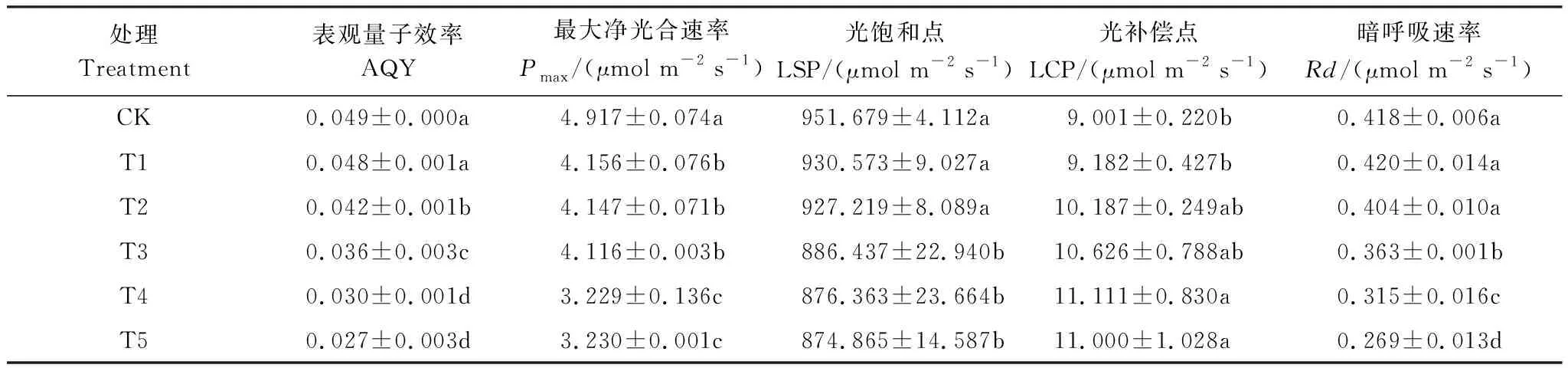
表3 不同水平鎘處理下楨楠幼樹葉片光響應曲線特征參數
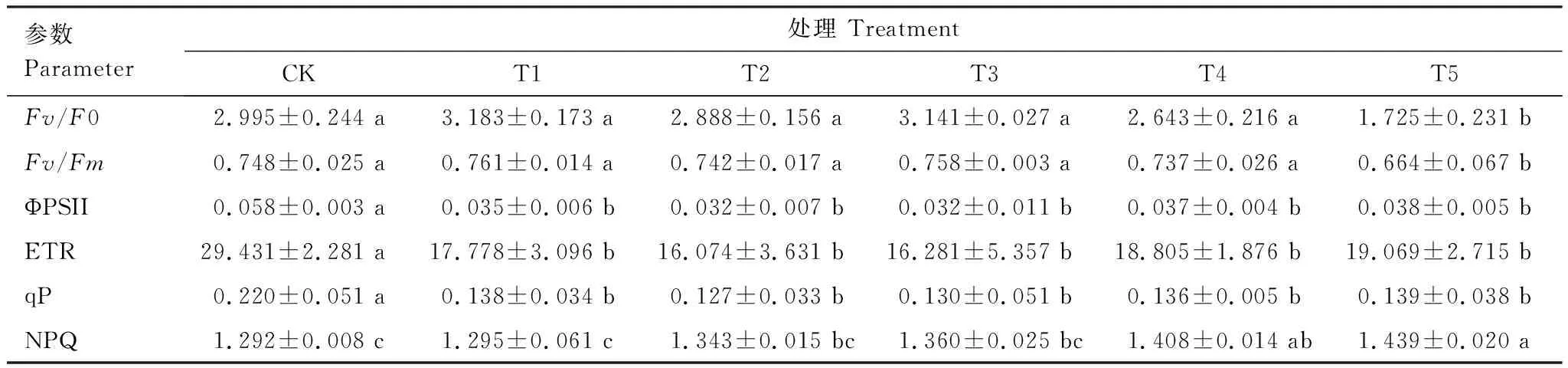
表4 不同水平鎘處理下楨楠幼樹熒光參數變化
2.5 鎘脅迫對楨楠幼樹吸收光能分配的影響
楨楠幼樹葉片對吸收光能的分配情況如表4所示。正常生長條件下,楨楠幼樹葉片吸收的能量約70%均用于熱耗散,約20%用于非光化學反應,約4%用于光化學反應,說明楨楠幼樹葉片對吸收光能的利用率較低,多通過熱耗散途徑耗散,以免光抑制。而Cd脅迫進一步減弱了楨楠幼樹葉片對吸收光能的利用能力,且在T2及以上處理時,天線熱耗散能量減小,過剩激發能增加,表明PSⅡ光化學反應中心受損,保護調節作用減弱。
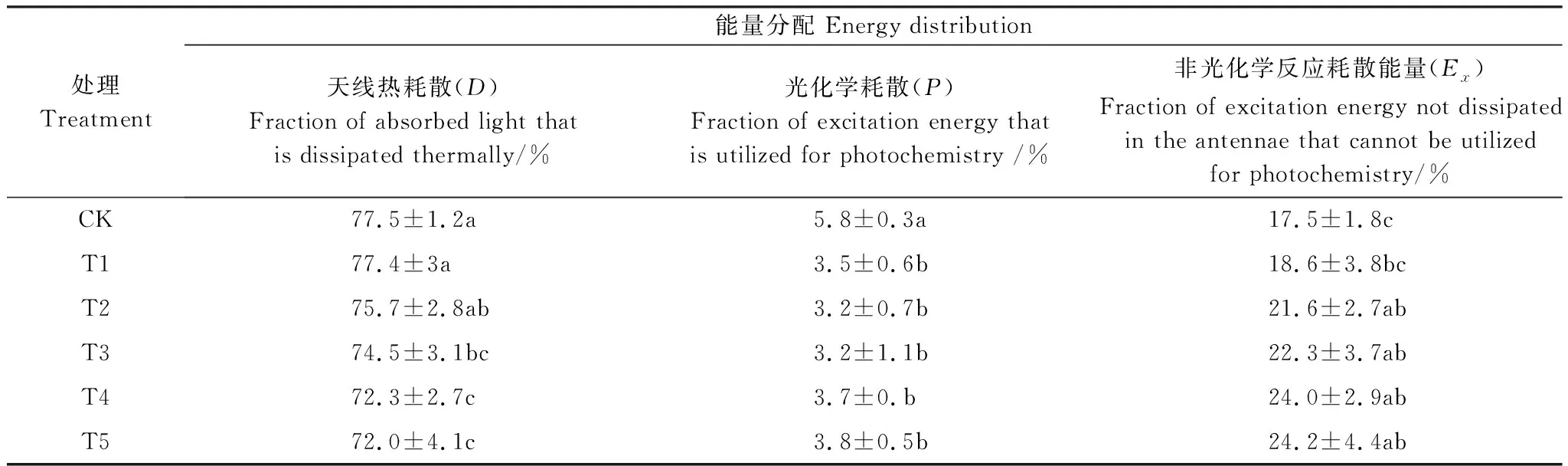
表5 不同水平鎘處理下楨楠幼樹葉片對吸收光能的耗散情況
3 討論和結論
3.1 鎘脅迫對楨楠幼樹葉綠素含量的影響
葉綠體色素是植物進行光合作用的基礎物質,其含量變化能反映植物光合作用的潛力。其中Chla、Chlb作為天線色素,在光能的吸收和傳遞中占主導作用,而Car除具備天線色素功能外,還可直接猝滅單線態氧,減小活性氧數量,以減輕逆境下植物所受的膜脂過氧化損傷。本研究中三者含量均呈下降趨勢,這可能是Cd與葉綠體中多種酶的巰基(—SH)結合或取代葉綠體的主要組成成分如Fe2+、Zn2+和Mg等,破壞葉綠體結構,加快了葉綠素的分解[7,25]。此外,我們在研究鎘對楨楠幼樹生長及抗性生理的影響(已另文發表)時發現細胞分裂素(CTK)含量有所減少,這也可能是導致葉綠素含量減少的一個重要原因[26]。當植物葉綠素含量不足時,若想維持較高的光合速率,便會調節Chla/b值,本研究中Chla/b值逐漸升高,應是楨楠對鎘毒害的一種防御機制[27],此外有研究表明,減少捕光葉綠素蛋白(Chlb)的合成數量,可以降低活性氧的積累[28],增強自身的耐性。
3.2 鎘脅迫對楨楠幼樹氣體交換參數的影響
Pn、Tr、Ci、Gs能夠反映植物光合生理對逆境的響應情況,是評價光合作用強度的基本指標。其中Pn變化的原因主要包括氣孔因素和非氣孔因素兩種[29],且根據Faqruhar和sharkeyt[30]的觀點,判斷葉片光合速率降低的主要原因是氣孔因素還是非氣孔因素,要依據兩個數值的變化方向,即Ci和LS;且Ci變化是主要判斷依據。本研究中,隨著脅迫程度的加重,Pn逐漸降低,Ci增加,LS降低,表明Cd脅迫環境中,限制楨楠幼樹光合速率的主要原因是非氣孔因素,這也與董袁媛等[27]研究鎘脅迫對黃麻光合作用的影響,譚長強等[31]研究鎘脅迫對蜆木光合生理的影響,所得到的結果相似。而引起Pn降低的原因可能較多,如葉片葉綠素含量不足、遭受光抑制,導致PSI和PSII的電子傳遞速降低等,這與簡敏菲等的研究結果一致[25],也與本文在葉綠素含量、PSII方面的研究結論相符。
3.3 鎘脅迫對楨楠幼樹葉片光合-光響應特征的影響
楨楠幼樹的Pn均隨著光合有效輻射的增強而逐漸增大,直至穩定,這與干旱、水淹脅迫下楨楠幼樹光合-光響應特征一致[32- 33]。許大全提出,AQY和Pmax同時下降是植物遭受光抑制的顯著特征[34]。本研究中,AQY、Pmax值逐漸降低,可見Cd脅迫下楨楠幼樹除對弱光的利用能力降低以外[16,35],強光及飽和光強下其光合速率也會受到明顯抑制。另外,Fv/Fm值變化也能衡量植物遭受光抑制程度[17,34],本研究中,其值在50 mg/kg時顯著降低(表4),結合AQY、Pmax和Fv/Fm值,可以認為,在Cd脅迫環境下楨楠幼樹遭受到一定程度的光抑制。而LSP降低,LCP升高則體現出楨楠幼樹對光抑制的適應能力,這與滕振寧等人的研究結果一致[36]。
3.4 鎘脅迫對楨楠幼樹葉綠素熒光參數的影響
葉綠素熒光作為PSⅡ光化學反應的指示器,對分析植物光合作用功能有重要作用。研究表明,Cd脅迫能抑制PSⅡ光合活性以及電子運轉[37]。Fv/Fm、Fv/F0在T1—T4處理時變化不明顯,這說明T1—T4處理時PSII反應中心受損程度較小,楨楠幼樹仍能維持一定的光能轉換效率以及光合活性,隨著Cd濃度的進一步增加(T5),Fv/Fm、Fv/F0顯著下降,表明PSⅡ綜合功能會有所下降,吳坤等[26]認為這與葉綠體超微結構發生改變有關。ΦPSⅡ和ETR均能反映植物在PSⅡ的實際光化學反應效率,本研究中,二者值均顯著下降,這與徐小遜等[[2]和蘇秀榮等[39]的研究結果相似,Cd脅迫導致楨楠幼樹葉片的光合電子傳遞鏈受到破壞,電子無法有效從PSⅡ向PSⅠ傳遞,同時PSⅡ反應中心也無法接受新的光子,一旦光子累積過多,葉片的細胞同化力(NADPH和ATP)則無法正常形成,C的固定和同化也無法完成,ΦPSⅡ和ETR也就因此下降,楨楠幼樹光合速率降低的原因可能也在于此,這與Vassilev和Yordanov[40]的研究結果一致。qP(光化學淬滅系數)能反映PSⅡ中天線色素吸收的光能參與光化學反應的比例。本研究中qP顯著下降,吳坤等認為這是植物處于Cd脅迫環境時,為防止自身PSⅡ原始電子受體QA被過度還原的適應現象[26],同時植物也會利用葉黃素循環途徑將過量光能猝滅耗散(NPQ上升),以降低自身光合機構的受損度[41]。
3.5 鎘脅迫對楨楠幼樹吸收光能分配的影響
植物葉片所吸收的光能通常包括3部分:天線熱耗散的能量(D)、光化學耗散能量(P)和非光化學反應耗散能量(Ex)[22,42]。本研究中,楨楠幼樹以天線熱耗散途徑為主來消耗過剩光能,Cd脅迫下光化學耗散能量減小,這與光合、熒光參數(AQY、Pmax、Fv/Fm)降低有關;而非光化學反應耗散能量的增加可防止PSII和電子傳遞鏈被過分還原,進而避免過剩光能對光合機構的過度破壞,這反映了楨楠幼樹對Cd脅迫的適應能力。
綜上所述,低濃度(T1—T2)Cd脅迫時,楨楠幼樹葉綠素含量、氣體交換參數和光響應特征參數就明顯下降,隨著Cd濃度的進一步增加(T3—T5),楨楠幼樹PSⅡ反應中心遭受一定程度的光抑制,對弱光以及吸收光的利用能力變弱,最終導致楨楠幼樹光合能力下降,因此不宜在鎘污染較為嚴重的地區大量發展楨楠人工幼林。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
