環境脅迫及相關植物激素在水稻根毛發育過程中的作用
2020-07-20 09:33:08李斌黃進王麗李瑾梁越洋陳稷
中國水稻科學 2020年4期
李斌 黃進, 王麗 李瑾 梁越洋 陳稷,
(1成都理工大學 生態環境學院,成都 610051;2四川農業大學 農學院,成都 611130;3中國林業科學研究院 資源昆蟲研究所,昆明 650224;4四川農業大學 水稻研究所,成都 611130;*通信聯系人,E-mail: huangjin18@cdut.edu.cn; jichen@sicau.edu.cn)
水稻是一種單子葉禾本科植物,對人類的重要性主要體現在兩個方面:1)水稻是人類主要的糧食作物之一,對人類生存有重要意義。水稻的果實可供食用或者作為食品生產及工業生產的原料,而水稻的秸稈也可作為飼料或者工業生產的原料;2)由于水稻是二倍體植物,并且具有易培養、生長迅速、基因組相對較小且測序已經完成等優點,它成為理想的單子葉模式植物[1]。隨著人口的增長與社會的發展,人們對稻米的需求日益增長,但受到有限耕地的制約。雖然當前陸地上有成片的閑置土地,如高原、沙漠、鹽堿等地區,但在這些地區存在水分、元素缺乏,滲透壓過高等環境脅迫,水稻在以上地區無法種植,或者在分蘗、開花、抽穗等階段受到抑制,以至于產量受到極大的影響[2-4]。因此,對于育種學家們來說,除了關注如何提高品種的產量以外,如何培育出抗逆品種以適應非傳統適耕土地如干旱少水地區、鹽堿甚至海灘,也是需要思考的問題。目前,海水稻等可在鹽堿或者淺海地區生存的水稻品種的育種工作也有所報道[2]。植物的根作為重要的營養吸收器官,不僅在水分和營養元素的吸收中發揮著重要作用,在植物應對干旱、高滲透壓等環境脅迫的過程中也同樣發揮著重要作用。當外界土壤干旱時,作為感受器的植物根系首先感受到危險并迅速發出信號,使植物對干旱脅迫做出一系列反應。首先根源脫落酸(abscisic acid, ABA)水平上調,引起葉片氣孔關閉,降低蒸騰失水,從而盡可能保持細胞水分,實現植物對干旱的耐受性。此外,干旱脅迫導致的 ABA代謝水平的變化也會改變根系的形態結構,間接影響地上部分生長發育,降低干旱對植物的影響[3]。而當植物處于水淹環境時,乙烯(ethylinne, ETH)在 植植物體 內的水水平升高, 導導致ABA在植物物體內的水平平下降,進而保保證植物可以以抵抗外界環境境的變化[4]。目前針對水稻稻如何應對環環境脅迫的機理研研究開展得較較多,也取得了很大的進展展[5],但研究對象象主要集中于水稻植株地上上部分或根系系,對于水稻根根毛在應對環環境逆境中的的作用及變化化卻很少涉及[6-77]。在水稻生生長發育過程中,根毛作為為水稻根部表皮皮細胞的特化化結構,占根系系表面積70%%以上,是水稻稻根系對水分分和養分吸收的主要貢獻者者。現有研究表表明,水稻根毛的發育主要要由遺傳因子子主導,即相關關基因通過調調節生長素(indole-3-accetic acid, IAA)等等植物激素的的代謝、ROS(reactive oxyygen species)及CCa2+的濃度及及細胞壁的合合成與水解等等實現對植物根根毛發育的時時空調控[8]。目前也有少量量研究表明植物物根毛的發育育也同時受到到植物所處的的環境條件的共共同調控[9]。基基于水稻根毛毛在水分與養養分吸收中的重重要作用及特特殊的結構和和發育的高度度動態性,可推推測它們在植物處于逆境時時,發揮重要要的作用,但對對于水稻根毛如何在逆境中發揮作用,以及植物激素素、遺傳系統如何調控根毛毛的生長發育育以應對逆境的的系統性總結卻幾乎未見報報道[10]。本文文正是從根毛的的結構、發育特點、調節根根毛的發育相相關植物激素(IAAA、ABA、ETH)與遺傳傳因子等角度度出發,整理“環環境—激素與與遺傳因子——根毛形態發發育”的相互關系系,并就其未來研究的方向向和方法提出出新的思路。
1 水稻根毛結結構和發育育過程
水稻的根為為須根系,即即由主根、側側根和須根構構成。。其按照生長長發育程度不不同又可以分分為四部分,即根根冠、分生區區、伸長區、成熟區[11]。水稻的根系系不僅僅是支撐器官官和營養器官官,同時也是是水稻感知外外界環環境如水分、營養元素水水平、有毒元元素等的感受受器。。不僅如此,在感受到外外界環境變化化時,水稻根根系通通過動態改變變根系內部基基因表達水平以做出相相應的的生理、生化化反應,進而幫幫助植物適應應環境脅迫。因此此,根系在水稻稻抗逆反應過過程中發揮著重重要作用[12],如在在干旱、營養養缺乏情況下下,通過提高高根系和根毛毛的長長度或者提高高根系和根毛毛的密度以獲取更多的的水分分和營養物質質[13]。水稻的的根毛是其根根系表面向外外延伸伸的單細胞結結構,不僅參參與水稻營養養吸收,同時時在水水稻根系與外外界微生物相相互作用及應對外界環環境變變化的過程中中發揮著極其其重要的作用用。在面臨外外界環環境脅迫時,包括水稻在在內的植物通通過對器官、組織織發育的調控控來加以應對對,而根毛對對外界環境的的響應應是植物通過過對根毛生長長發育的三個階段的分分別調調控實現的[12]。
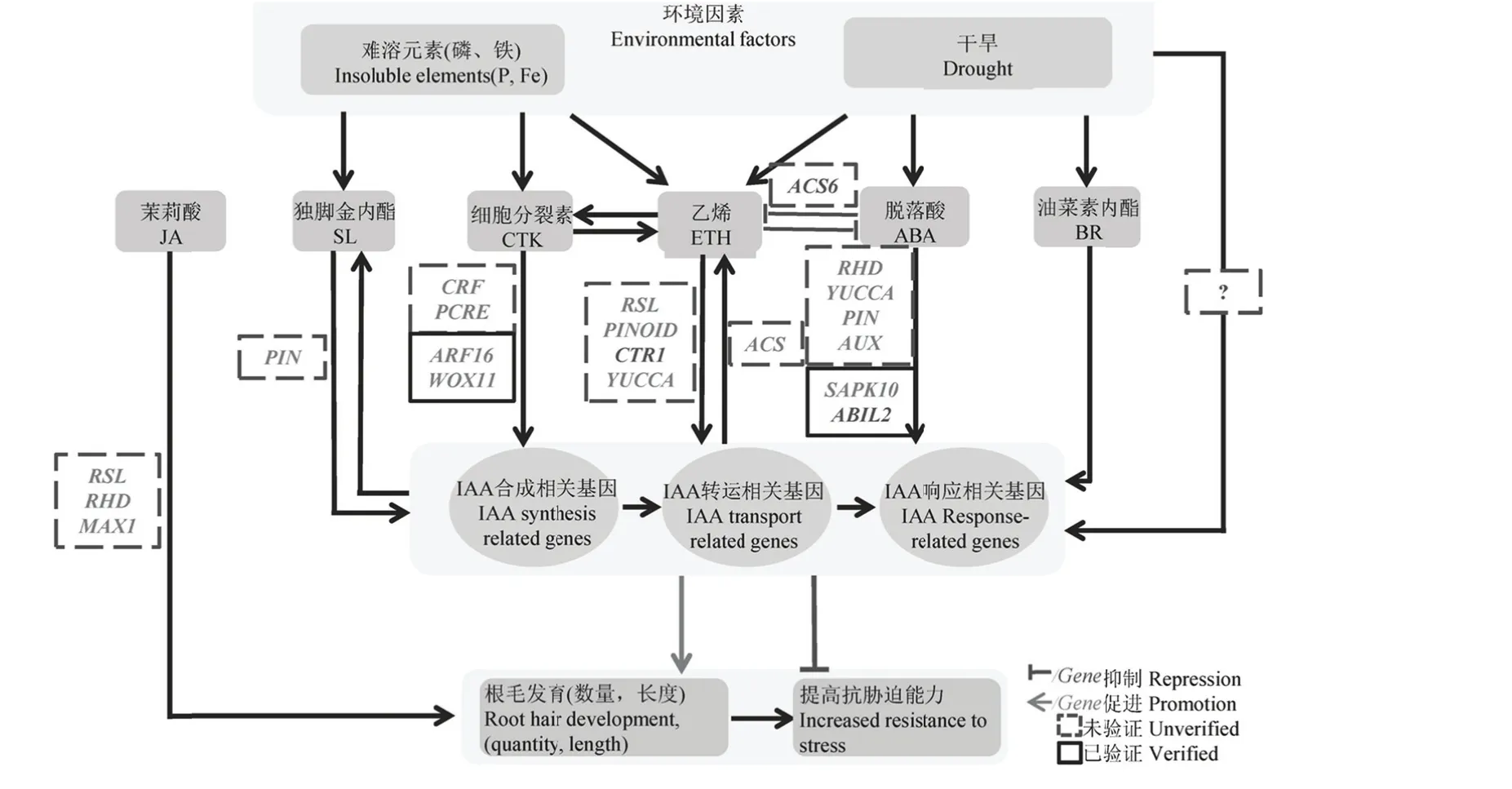
圖1 環境脅脅迫與植物激素素調控水稻根毛毛發育Fig. 1. Pathwaays of environment factors aand phytohormmone regulatedd rice root hair development..
水稻根毛的的生長發育同同時受到遺傳和環境因素的的嚴格控制,共分為三個個階段,分別別為:1)根毛毛細胞胞命運決定階階段:植物中中根毛細胞確確定,按其排排布模模式共分為三三種,水稻屬屬于第Ⅱ種(隨隨機模式),被確確定的細胞稱稱作H細胞(hhair cell)[14];2)根毛尖端端起始階段:指即將發育為根毛的根表皮細胞的細胞壁一側開始膨脹, 并在其后逐漸變寬、變長,形成管狀突起[12];3)根毛尖端伸長階段:指根毛細胞突起后,進入頂端生長階段。隨著復雜的生化反應,根毛細胞的頂端會由分泌小泡構成一個細胞致密區域,同時新合成的質膜和細胞壁物質向頂端運輸。當根毛頂端生長到適當程度, 根毛停止生長并進入成熟階段[12]。通過對擬南芥的研究,我們已經對根毛生長的這三個階段所涉及的各種生理、生化、細胞過程有了一定認識,大量相關基因及其功能也已被確認[12]。但與擬南芥相比,對于水稻或單子葉植物的根毛生長發育在分子水平上的認識仍然相當有限。到目前為止,在基因或者分子水平上證明參與水稻根毛發育的基因包括根毛細胞特異表達的 OsCSLD1(cellulose synthase-like D1)基因[15]、OsRHL1 (root hairless1)[16]以及 RHS (root hair specific)、RHEs (root hair-specific cis-elements)、EXPA7 (expansin A7)、OsSRH (short root hair)家族的OsSRH1、OsSRH2 以及OsFH1 (formin homology 1 )等[9,11]。但就整個根毛發育調控網絡而言,尚有很多的環節缺失,并且水稻中激素對根毛發育作用相關研究也非常不足[11]。
2 環境脅迫對植物根毛形態的調控作用
已有研究表明,在植物處于光照不良、干旱、重金屬脅迫、營養元素缺乏等環境脅迫下,根毛的表型會產生相應的變化[11,17]。一般來說,在外界環境變化時,植物通過改變根毛與非根毛細胞的比例、改變根毛長度等方式增加或減少根表面積以適應外界的各種脅迫,從而提高植物的生存能力[18]。更具體地講,在干旱或者營養缺乏的情況下,植物通過增加根毛長度或者密度去獲得更多的水分和養分;當植物處于外界重金屬脅迫時,則降低根毛長度或者密度以盡量減少對重金屬的接觸和吸收。如在干旱脅迫下,受到 ETH、ABA調控的 RHD4(root hair defective 4)根毛生長相關的基因表達量變化,伸長根毛以獲得更多水分(表2)[12,19-20]。營養元素缺乏時,植物通過伸長根毛增加根表面積,增加對土壤養分的吸收,如磷、鐵和鉀等在土壤中相對固定的營養元素[21]。擬南芥中根毛的密度會隨著生長環境中錳或鐵的減少而增加[22],而缺氮脅迫通過調控bZIP轉錄因子TGA1 (TGACG-binding Factor 1)/ TGA4和CPC (Caprice),共同作用促使擬南芥根毛長度、密度增加[23]。在外界有毒重金屬脅迫時,植物通過改變根毛長度減少重金屬對植物的毒性,如重金屬鎘脅迫導致擬南芥根毛伸長等[24]。在各種營養元素中,磷元素與根毛發育關系最為密切,約90%的磷吸收是由根毛完成的[25]。由于土壤中磷元素多以非水溶狀態存在,植物根系無法依靠離子交換和離子遷移來完成吸收,因此只有盡量增加根系與土壤的接觸面積來提高對磷的吸收,而更多、更密的根毛對植物來說是效率更高且成本最低的適應策略。事實上,研究人員已經清晰描繪了擬南芥在磷脅迫下根部發育的三個主要變化過程。首先,主根停止向下生長,隨后側根數量增加。隨著側根增加,根的表皮細胞數量增加也使根毛細胞的數量顯著增加,同時也增加了根毛的長度。最后,根表皮細胞分泌的酸性磷酸酶催化土壤中的有機磷轉化為無機磷酸鹽,從而使擬南芥獲取更多磷元素。在缺磷狀態下,無論是擬南芥還是水稻,由IAA合成、轉運及IAA響應的轉錄因子構成的調控網被激活,似乎都是決定根毛生長關鍵因素[26]。因此,外界環境變化與根毛細胞生長發育之間存在著明確的聯系,而其他一些研究也明確了這一點。因此,在這些環境調控根毛表型的過程中,一般來說都會伴隨著植物激素表達量的變化[17,19-20]。
3 與環境脅迫相關植物激素參與水稻根毛調控
水稻作為一種模式植物,對于其抗逆相關研究已經有若干進展,大量水稻抗逆相關路徑已闡明。但與擬南芥相比,水稻根毛的生長發育以及抗逆相關的分子機理研究較少,尤其是環境脅迫影響根毛發育的分子機理仍需要深入的研究。現有研究表明在環境影響植物根毛發育的過程中,一些受到影響或者參與其中的基因也同時與激素的代謝、轉運或者信號通路相關基因存在著重疊,即環境因子對根毛發育的調控作用很大程度上是植物激素相關基因通過對激素的合成、轉運、降解等過程的調控來實現的。當水稻感受到干旱或者營養元素缺乏時,通過體內ABA、IAA、ETH等激素水平變化間接調控根毛生長,以提高植物對相應環境逆境脅迫的耐受能力。如缺磷脅迫情況下,水稻通過調控OsAUX1基因改變根部IAA的含量,從而影響根毛的伸長(表1),促進水稻根毛伸長以獲取更多的磷元素[27];而缺鐵脅迫下,水稻或者增加根毛數量[28],或者利用ETH促進根毛的伸長生長,從而得以最大限度吸收鐵元素[29]。因此,在正常條件下植物激素在水稻根毛發育中扮演著重要角色;而在逆境條件下,根毛也是植物激素發揮其“抗逆指揮官”作用的重要環節之一。目前在水稻中的研究已經證實參與根毛與逆境脅迫互作的植物激素主要可能是 IAA和ABA[30-31],而ETH則在擬南芥中的研究較為透徹[32]。

表1 水稻中參與“環境-激素與遺傳因子-根毛形態發育”的基因Table 1. Rice genes involved in the root hair development regulated by both environmental stress and phytohormones.

表2 擬南芥中參與“環境-激素與遺傳因子-根毛形態發育”的基因Table 2. Arabidopsis genes involved in the root hair development regulated by both environmental stress and phytohormones.
3.1 生長素(IAA)對水稻根毛發育的調控作用
IAA是水稻根毛發育的關鍵因素,IAA不僅誘導水稻根毛的伸長[32],并且在水稻抗脅迫過程中也起著極為重要的作用[33]。IAA是在植物生長旺盛部位合成的極性小分子弱酸性化合物(吲哚類似物),其本質是吲哚乙酸。IAA參與細胞壁的形成和核酸代謝,并在調控植物的生長的過程中發揮重要作用[34]。適當濃度的IAA為水稻根毛發育所必需,但IAA濃度過高時,反而會嚴重抑制水稻根毛的伸長生長[35-36]。進一步研究發現IAA主要通過合成、降解和運輸等方式改變其在植物體內積累的部位和濃度,以此調控植物的生長發育以及根毛的生長[37-38]。植物體內IAA合成路徑有兩條,分別為色氨酸途徑和非色氨酸路徑。而在植物合成IAA主要依賴的色氨酸途徑中,YUCCA家族是負責將吲哚丙酮酸IPA(in-dole-3-pyruvate)轉化為IAA的限速酶基因[39-40],同時也參與植物中根毛生長調控[28]。在擬南芥中過表達YUCCA1可促進根毛生長,而通過施用IAA合成抑制劑,則可以抑制根毛的形成,而在恢復實驗中對受到抑制劑抑制的植株施用外源IAA,根毛的表型則又可得到恢復。以上研究結果證明YUCCA在調控水稻根毛生長中IAA合成路徑的重要作用[40-41]。
IAA在植物體內的運輸方式主要有兩種:1)由濃度決定的自由擴散,主要是在韌皮部;2)依賴載體和能量的極性運輸,主要在細胞內和細胞間進行。通過調控IAA運輸載體的活性和細胞內定位,植物可以調控體內不同部位 IAA濃度,實現對生理、生化反應與生長發育的調控作用,以面對外界環境的變化[26]。在IAA的極性運輸過程中,主要參與的基因家族包括 IAA輸入載體 AUX1/LAX(AUXIN1/like AUX1)和輸出載體,包括 PIN(PIN-formed)、類 PIN 蛋白和 ABCB/PGP (ATP binding cassette B/P-glyco-protein) 等。其中OsPIN、OsAUX1/LAX家族也是調控水稻根系生長的主要基因家族。
在 OsAUX基因家族中起關鍵作用的是OsAUX1,實驗發現Osaux1突變體根毛長度變短,且2,4-D (2,4-Dichlorophenoxyacetic acid)和NAA(1-naphthylacetic acid )也不能恢復其短根毛的表型,證實了OsAUX1在水稻根毛生長發育中的關鍵作用[42]。該結果也在其他研究中得以證實[43]。雖然OsAUX1表達模式與擬南芥AtAUX1類似,但對水稻根毛的研究發現OsAUX1只存在于初形成的根毛細胞中,在成熟根毛細胞中和非根毛細胞中均不表達,該結果表明OsAUX1可能在根表皮細胞是否成為根毛的這一決定階段發揮作用,并且這個作用是通過影響生長素的運輸來實現的[27,42]。此外,OsAUX1還參與植物應對外界磷缺乏或者鎘脅迫的反應。在磷缺乏條件下,OsAUX1促使根毛伸長,而在植物面對鎘脅迫時,則參與氧化應激反應(表1)[43]。OsAUX家族成員OsAUX2也在根毛中表達,因此很有可能也參與根毛生長調控[42],而OsAUX3,OsAUX4和OsAUX5等基因是否參與對根毛生長發育的調控則還需要進一步探索。
WindML的圖形界面基于像素,用顏色匹配表說明位圖中的顏色。首先在顏色匹配表上設置一種顏色的紅、綠、藍亮度值,然后用位圖陣列中的每個像素值索引值代表該點在顏色匹配表中的顏色。
OsPIN家族共有12個成員,研究認為OsPIN家族成員在通過改變他們在細胞膜上的分布決定IAA的極性運輸[44-45]。其中,OsPIN1定位于質膜并在維管組織和根原基中表達(表1)[42]。OsPIN1在IAA處理后表達量增加并誘導根毛的形成,而OsPIN3參與應答干旱脅迫,但具體的機制還不明確[46]。
ABCB/PGP家族是ABC轉運蛋白家族的1個亞家族,在擬南芥中對該家族蛋白的研究較深入。擬南芥中PGP家族共有22個成員[47],對AtPGP1、AtPGP4、AtPGP19基因的功能已有相關的研究,三個基因都參與對IAA的極性運輸,其中AtPGP1負責編碼IAA輸出載體,而在 AtPGP1或AtPGP19兩者發生突變后擬南芥 IAA運輸都受到影響[48]。AtPGP4則同時具有輸入和輸出功能,其功能缺失后擬南芥根毛變長,而過表達AtPGP4使植物根毛變短(表 2)[49]。水稻也有 22個ABCB/PGP家族成員,但只有OsABCB14被證明在生長素轉運及維持植物體內鐵平衡的過程中發揮作用[50],對于水稻中其他OsABCB成員的功能還有待進一步探索。當然還有一些基因參與調控IAA通路,如OsWOX3A通過調控 IAA轉運相關基因表達調控水稻根毛的形成[51]。ARF(Auxin response factors)家族作為生長素的響應因子參與 IAA調控路徑,如 ARF家族的ARF12對根毛有調控作用[52],ARF16受CTK調控參與根毛生長,并且參與磷脅迫以及鐵脅迫途徑(表1)[53]。但對于ARF家族中其他成員在水稻根毛生長中是否發揮作用及其相關機理的研究卻并沒有太多進展。因此,整體而言,當前對水稻中IAA調控根毛生長機制的研究仍有很大的探索空間。
3.2 脫落酸(ABA)對水稻根毛的影響
脫落酸(abscisic acid,ABA)是一種抑制生長的植物激素,多在植物應對外界不良環境的過程中發揮作用,其在環境脅迫調控植物根毛伸長的過程中起著重要的作用。有研究表明,適量的外源 ABA可以誘導水稻根毛的形成,促進水稻根毛的伸長,并伴隨新蛋白的合成[54-55],但過量的 ABA卻抑制水稻根毛的伸長[56]。
擬南芥中ABA的核心信號通路已基本清晰[57],當ABA受體PYR1 (Pyrabactinresistance1)/PYL(Pyraba ctin resistance-like)/RCAR (Regulatory Components of ABA Receptor)與ABA結合時,形成的聚合體再與下游的PP2C (A type 2C protein phosphatase)結合,抑制SnRK2 (SNF1-related protein Kinase 2)磷酸化激活bZIP轉錄因子,從而實現對ABA響應基因表達的調控[31]。環境脅迫極有可能通過該路徑間接調控植物根毛形態。
水稻中 ABA信號通路應基本和擬南芥相同,目前已鑒別出OsPYL/RCAR家族10個成員、PP2C家族9個成員、SARK(stress/ ABA-activated protein kinase)10個成員以及10個b-ZIP轉錄因子成員。此外,還有在水稻 ABA信號通路中起著抑制作用的OsABIL2 (OsABI-Like 2)基因。ABIL2和負責去磷酸化的SAPK10形成復合體,通過影響b-ZIP轉錄因子,對ABA響應基因進行調控[57]。而當前研究發現過表達OsABIL2會顯著導致根毛長度變短[57],而過表達SAPK10則導致長根毛表型(表2)[56],說明ABA極有可能通過PYR/PP2C/SNRK2信號通路調控水稻根毛伸長,而擬南芥中發現ABA通過調控生長素運輸載體PINOID的表達改變根毛表型[58-59]。因此,ABA很可能最終也通過IAA運輸通路實現對水稻根毛生長的調控。
3.3 乙烯(ETH)對水稻根毛的影響
ETH是一種氣體植物激素,也是最簡單的烯類化合物。ETH以蛋氨酸為合成原料,1-氨基環丙烷-1-羧酸(1-aminocyclopropane-1-carboxylic acid,ACC)合酶和ACO (ACC Oxidase)為關鍵酶,通過簡單的線性途徑在植物的各個器官均可合成[60]。在植物根毛的生長發育過程中,植物內、外源ETH或者ACC都能促進植物的根毛生長[32,61]。
ETH對根毛生長調控很有可能是通過ETH的信號通路實現的[32]。在擬南芥中,參與 ETH信號通路的成員主要包括 CTR1 (constit-utive triple response 1), EIN2 (ethylene insensitive 2), EIN3(ethylene insensitive 3),以及EIN3-like 1,RTE1(reversion to ethylene sensitivity 1), EBF (EIN 3 binding F box) 1和2,ETP1 (EIN2- Targeting protein 1)和ETP2等。當ETH和ETH受體結合后激發CRT1活性[62-63],導致EIN2的低磷酸化,進而激活EIN2蛋白水解酶,EIN2蛋白被水解[64]。而位于EIN2下游的是負責激活ETR (ethylene receptor)的PIN3以及具有類似功能的ELI 1(EIN3-like 1)[65],由此構成反饋抑制通路。當EIN2未被水解或磷酸化水平較低時,部分活化的EIN2通過抑制EIN3連接酶EBF1(B-cell Factor 1)和 EBF2,保護 EIN3/ER1(endoplasmic reticulum)免于被蛋白酶降解。而對擬南芥的研究已表明,過表達 ETH合成相關基因會造成植物細胞中EIN3/EIL1蛋白量的增加及CTR1基因的失活,最后促使根毛伸長(表2)[62-63]。而CRT1在鹽脅迫下高表達,參與鹽脅迫反應[62,66]。同時EIN3/EIL1與 RHD6/RSL1相互作用調控 RSL4、RSL2,進而改變根毛表型(表2)[67]。EIN2基因突變會導致短根毛表型,并降低植物對 IAA的敏感度[68-69]。在鎘和砷等重金屬脅迫下,EIN2通過改變根毛長度以減少重金屬攝取量(表 2)[70]。因此,擬南芥中EIN1、EIN2、EIN3是ETH調控根毛表型的關鍵基因[56,71]。大豆中的研究表明ETH合成上游基因 ETR1 (ethylene response mediator)、ETO1(ethylene overproduction 1)的變化對根毛表型的影響可能是通過改變ETH水平實現的(表2)[72]。EIL1則已被證明參與鐵脅迫調控(表 2)[73]。此外,通過對擬南芥的研究也發現 ETH 以“雙重作用”的方式影響根毛發育[61]。在決定細胞是否發育成根毛的階段,ETH促進更多根毛細胞突起和促進頂端生長,如 ACC處理可誘使非根毛細胞轉變為根毛細胞,使處理部位異位根毛顯著增加[71,74],而在根毛伸長的階段,ETH則繼續促進根毛的頂端生長[75]。此外,ETH同樣也是植物應對環境脅迫的關鍵激素,在外界環境脅迫下植物內源 ETH合成量增加[17]。這些研究結果證明 ETH在植物受到外界環境脅迫時,可能通過發揮其在調控根毛生長中的重要作用來幫助植物應對不良環境。
相比于擬南芥,水稻中 ETH的研究尤其是與根毛相關的研究較少。但相關研究表明,植物細胞內 IAA的大量積累會導致水稻產生過量的 ETH,進而對水稻根毛的伸長進行抑制[36]。水稻 RHL家族中受到OsERF1調控的RHL1及RHL4都與根毛發育相關(表2)。相關研究認為ETH在調控根毛生長過程中,很有可能需要RHL家族成員的參與,從而在水稻根毛形成過程中發揮重要的作用,但這一假設還需要分子遺傳學和細胞生物學相關數據的支持[78]。
3.4 環境脅迫、IAA、ABA及ETH共同調控水稻根毛生長發育
許多研究表明脅迫相關激素ABA、ETH和IAA共同調控植物根部生長,如有研究發現擬南芥參與IAA介導的反應的IAA響應因子ARF2突變后,植株對ABA會更加敏感,而ABI4介導的ABA抑制側根形成途徑也對IAA極性轉運有影響,降低根部IAA的積累[79]。以上研究結果表明IAA和ABA在調控根毛發育過程中很可能是共同發揮作用的。
在水稻 ABA信號通路研究中發現 OsSAPK10的過表達會造成水稻根毛變長,而OsABIL2基因的過表達則抑制根毛伸長,由此可以確定 ABA參與調控根毛生長(表1)[56]。ABA通過PYR/PP2C/SnRK信號通路調控IAA在根部特定部位的轉運和濃度,從而間接促進根毛的伸長。如 ABA處理后,水稻植株IAA合成相關基因YUCCAs表達量與IAA濃度升高,促進根毛伸長。有關研究也確定在水稻受到外界脅迫時,ABA通過與IAA極性運輸相關基因協同作用改變IAA濃度從而促進根毛伸長,以對抗外界不良環境[56]。經ABA處理后,水稻IAA轉運載體OsPIN和AUX/LAX、PINOID基因表達量升高,因此 ABA應是通過調節 IAA運輸相關蛋白PIN、AUX的動態平衡從而調控根毛生長[56]。對擬南芥進行ABA處理后也會導致處理部位IAA濃度增加,也佐證了ABA調控根毛生長是通過調控IAA合成和轉運相關基因進行的[56],即ABA位于IAA上游對根毛生長發育進行調控。進一步的研究發現,敲除ARF1會導致水稻萌芽期對ABA的敏感性升高,而擬南芥在外界滲透壓改變的時候 ABA調控PIN2表達。此外,PINOID在擬南芥中已經被證實在根毛細胞中的表達對控制根毛的伸長有著重要作用,并受到IAA和ABA的共同影響[80-81]。而受到 IAA和 ABA共同調控的 RopGEF4和RopGEF10在擬南芥根毛中通過 ROS系統負調控ABA信號途徑參與調控根毛的生長[82]。因此,在調控根毛生長的過程中,ABA和IAA之間相互調控的詳細作用方式可能未必這么簡單,仍需要進一步的探索,而且這也是一個值得深入研究的領域。
除了ABA和IAA之間相互影響的研究外,ETH和 IAA在控制植物根毛生長的過程中的相互關系也是一個值得研究的問題。在擬南芥中,IAA和ETH通過一系列復雜的相互作用參與對根系生長發育的調控,也包括控制根毛的細胞位置確定、根毛生長的起始和伸長等[35,83]。擬南芥中,ETH對根毛生長的調控已被證明是通過 EIN3/EIL1和RHD6/RSL1a(RHD 6 LIKE1a)路徑[35],并且可能是通過調節植物體細胞內 IAA濃度梯度從而間接調控根毛的生長[84]。
隨著研究的深入,人們發現IAA和ETH之間的關系并不是簡單的上下游調控關系,而更有可能是相互調控的模式。相關研究發現,在擬南芥中IAA促進ACS (1-carboxylic acid synthase) 基因的轉錄和ETH合成[85],而外源ETH也可以促進IAA合成相關基因的表達,并提高IAA水平[33]。進一步的研究發現IAA合成相關的基因,如ASA1(anthranilates synthase α1),ASB1(anthranilates synthase β1)、YUCCA、TAA1、TAR1(tryptophan amino transferaserelated protein 1)都受到ETH的調控[61,83]。此外,IAA不敏感突變體(axr2,axr3)也對ETH不敏感,具體表現為根毛起始數量減少[86]。ETH的合成過表達突變體eto1的長根毛表型也可通過阻斷IAA轉運載體AUX1進行抑制[35]。在根毛生長發育相關基因RHD6缺失突變中施加外源ACC或IAA可以恢復該基因突變導致的根毛數量變少的表型[87]。當ETH信號通路中關鍵基因EIN2發生突變,其突變體ein2-1根毛生長受到抑制,對ein2-1突變體使用NAA進行處理時,可以恢復 ein2-1突變體短根毛表型[32]。AUX1和EIN2雙突變體的根毛數量要少于單一基因的突變體,因此根毛生長的起始和延長很可能是ETH和IAA共同決定的[35]。但在對控制根毛的數量和長度的 IAA信號傳導相關基因 AXR2(IAA7), AXR3 (IAA17), SLR1 (IAA14) 和IAA28的突變體檢測后發現,這些突變體對過量 ETH并不敏感,即這些突變體在 ETH處理后沒有出現根系生長受到抑制的表型[88]。從這個角度來說,ETH和IAA又未表現出相互調控作用,因此很難簡單定義ETH和IAA之間的相互作用。目前,雖然對于ETH和 IAA之間的相互作用以及各自獨立的作用都有一些研究報道,但二者之間的關系還遠沒有明確。但根據目前已經掌握的信息,我們幾乎可以確定,與ABA和IAA簡單的上下游關系不同,ETH和IAA之間可能是更為復雜的相互作用關系,它們在調控植物根毛發育的過程中可能不是一個直線關系,或者在不同的條件下,它們的關系會發生一定變化。
ETH和ABA之間的相互作用是比較復雜的,目前在根毛生長發育的調控過程中,對二者之間的關系的研究相對較少,在擬南芥中,etr1-1和ein 2突變體根系對ETH和ABA都表現出抗性,在氧化應激下,ABA下游元件ABI1調控ETH合成基因ACS6,改變 ETH水平[89]。ABA也可以誘導 ETH下游ERF4的產生,而當過表達ERF4時, 植物對ABA敏感性降低[90]。這些都表明ETH和ABA之間存在相互調控作用。但ETH和ABA之間的互相調控作用是否在他們調控根毛的生長發育過程中也按照同樣的模式作用還未見相關研究結果。但我們現在可以確定的是外部環境,尤其是環境脅迫可影響 ABA、ETH以及有其他與環境相關激素的水平,ABA通過改變水稻根部IAA濃度調控水稻根毛的生長發育,而ETH通過和IAA相互作用調控植物根毛的生長發育,更通過雙重作用影響根毛生長的起始和伸長。而ABA和ETH在植物抗脅迫中相互影響,那么二者在外界生物脅迫和非生物脅迫條件下以某種方式共同調控 IAA水平的可能性也是不能排除的。這其實也是非常值得研究者深入挖掘的領域。
3.5 其他植物激素調控水稻根毛生長發育
除了現在已經探明在外界生物脅迫、非生物脅迫中起著主要作用的ABA、ETH和IAA以外,還有其他一些激素與水稻根毛生長相關,其中一些可能也參與抗脅迫信號通路或者與 IAA信號通路有著協同作用。
油菜素內酯(Brassinolide,BR)屬于油菜素甾族化合物,是植物生長發育的必需激素。研究發現BR可以增加水稻的耐旱性,在一定的濃度范圍內促進水稻根毛的生長發育,尤其濃度低時促進作用非常明顯。但也有研究表明在擬南芥中 eBL (24-Epibrassinolide)即使濃度低至0.001 nmol/L也只表現為對根毛發育的抑制作用,而水稻中一定濃度的eBL促進水稻根毛生長,利用BR抑制劑BRZ后會抑制水稻根毛生長[91-92]。這些線索表明BR也在水稻根毛發育中起著一定作用,參與生物和非生物脅迫環境下根毛生長調控,但BR調控水稻根毛的具體調控路徑還不清楚,而擬南芥研究中證明BR可能是在 ETH上游促進根毛生長,因為施用 GR24會促進植物根毛伸長,而施用ETH抑制劑AVG則引起根毛減少。而且 AVG引起的根毛減少不能通過施用GR24恢復也間接證明BR可能是在ETH的上游發揮作用[68]。但我們依然不能排除BR在調控細胞伸長過程中也是通過影響 IAA的調控網絡發揮作用的[93]。當前BR在水稻和擬南芥根毛發育中的不同表現,僅僅是兩類植物對BR敏感程度的不同,抑或是它們的 BR-根毛調控信號通路存在著很大的不同,仍是未解之謎。
茉莉酸(Jasmonate,JA)是植物體內一類非常重要的脂肪酸衍生物,它們參與調控植物的生長發育并對外界生物、非生物脅迫作出響應。JA調控擬南芥根毛發育的分子機理已有相關報道,如施用過量JA促進擬南芥根毛伸長,阻斷JA調控通路則會導致短根毛表型[93]。相關研究表明,JA通過抑制RHD6轉錄,促進擬南芥根部根毛伸長[84]。加之ETH相關基因 EIN3/EIL1也和RHD6/RSL1共同作用促進擬南芥根毛生長, 因此JA和ETH很可能通過在同一信號通路或者共享一些關鍵基因來共同調控植物根毛表型。RHD6屬于 bHLH (basic helixloop-helix protein)基因家族,在水稻中有 165個bHLH基因,其中部分基因在耐低溫,調控營養元素(如鐵離子)的吸收等方面發揮作用[94]。但目前對水稻bHLH家族研究較少,并且對該家族和植物激素相互作用的研究也有待深入。在擬南芥中發現的JA通過調控RHD6/RSL1蛋白控制根毛發育的調控通路,在水稻中也可能存在類似機制,只是尚未有相關的研究報道[84]。
通過對水稻中細胞分裂素(Cytokinin,CTK)的研究,發現CTK和IAA通過相互調控[95],影響水稻根毛的發育。外源6-BA在抑制水稻根生長的同時,根毛的發育也受到抑制[96]。其可能是通過水稻中信號轉導調控[97]。水稻中 CTK雙元組分信號系統已鑒定 37個基因,其中,CTK響應因子 CRF(cytokinin response factor) 通過一個特殊的PINCTK響應元件 PCRE (pin cytokinin response Element) 調控IAA運輸載體PIN的轉錄[98],即CTK通過PIN調控水稻生長素的轉運。用6-BA處理水稻后可影響水稻根毛的表型[96],下調水稻根部OsAUX的表達,推測 CTK可能通過同時調控OsAUX1和 OsPIN調控根毛的表型[96],但在OsAUX1啟動子上并未找到CRF的結合序列,由此判斷,CTK對OsAUX1的調控可能是以間接的方式,或者需要其他激素的參與,但目前尚缺少直接數據支持。水稻中也發現受到參與 CTK調控水稻根毛生長的OsWOX11在干旱脅迫下表達增加并提高水稻抗旱性,說明 CTK可能在環境調控根毛這一過程中起著重要作用(表1)[99]。
在擬南芥中CTK能夠增加ACC穩定性并提高ETH的水平,IAA和ETH也能夠恢復CTK過表達造成的短根毛表型。此外,在IAA和ETH缺失的情況下,CTK也能夠促進根毛的伸長[35]。以上結果暗示著IAA、ETH、CTK組成的網絡在擬南芥調控根毛的生長發育過程中的作用是相當復雜的,對該調控機制的了解還遠遠不夠。而在在水稻中是否有類似的調控網絡,其與擬南芥對應的調控網絡相比有何區別,有哪些基因參與其中等仍有待探索。
獨腳金內酯(Strigolactone,SL)是近年來才發現的一類通過類胡蘿卜素合成途徑合成的植物激素[100]。研究表明,SL可能通過控制細胞內IAA輸出載體PIN的水平來調控植物的分枝數量、不定根的數量及根毛的數量和長度[68,101]。反之,SL的合成亦受到IAA的調控,由此兩者的合成、轉運構成了一個反饋抑制通路[102]。
此外,有研究表明SL在植物面對外界磷水平變化時,在決定植物根冠比、植物側芽及側根的生長等過程中也發揮著關鍵性的作用。在磷充足的條件下,SL抑制植物側根的形成;而在磷不足條件下,SL則會促進側根的形成以幫助植物獲取更多的磷[103]。在低磷水平下,IAA受體基因TIR1 (transport inhibitor response 1) 的表達水平會升高以提高植物對 IAA的敏感性(表 1),而在 SL信號通路 max2 (More axillary growth 2 )突變體內,TIR1的表達水平反而會降低,這說明SL與IAA在磷不足條件下共同控制根毛伸長[29]。不止于此,外源SL可促進擬南芥根毛的伸長生長,而且這種促進作用除了必須有MAX1 (more axillary growth 1)基因的參與,還要依靠IAA和ETH信號通路的共同參與,但MAX2 或者SL對于IAA發揮對根毛生長發育的調控作用卻不是必需的[68,101,103-104]。整體而言,作為一種新發現的植物激素,目前認為SL在水稻中的作用主要是控制分蘗和不定根的數量及在氮、磷不足條件下控制主根及不定根的長度和數量[105-106],至于SL是否如在擬南芥中一樣參與根毛發育的調控,又是如何實現這種調控的都有待于進一步的研究。
4 展望
水稻生長發育的過程也是它們不斷對抗、適應生物脅迫和非生物脅迫等不利環境的過程。這些環境的變化激活植物體內與脅迫相關的植物激素如IAA、ABA、ETH、GA3、JA、BR、SL等,隨后這些激素調控水稻體內的細胞、生理、生化反應。這個過程對于保障水稻的生存及生長發育有著極其重要的意義。當前關于生物和非生物脅迫影響水稻生長的研究集中于植物地上部分和地下部分整體的研究,對于根毛的研究相對較少。根毛的延伸對于水稻獲得更多的營養元素,提高營養物質吸收效率,增加水稻的抗逆性有著極為重要的作用。深化“環境-激素與遺傳因子-根毛形態發育”三者之間聯系的研究對于突破水稻根毛研究的瓶頸,了解水稻的抗逆機理,為育種學家的定向育種工作提供更多的遺傳學材料和依據都有重要的意義。
當前通過研究不同外界環境變化導致水稻的根毛表型及相關信號通路變化,發現ABA、ETH、IAA激素在外界脅迫改變根毛表型中可能起著核心作用。ETH與IAA共同作用控制根毛定位、數量,ABA與IAA共同作用控制根毛伸長[56]。在以上激素調控根毛發育的過程中,主要環節包括水稻體內的ABA、ETH激素水平變化、調控IAA信號通路中運輸載體PIN和AUX基因表達量或者蛋白質特性的變化及根部IAA水平的變化等。這些變化最終促進水稻根毛的形成或者伸長生長,當然 YUCCA基因也可能是其中重要的一環[107]。
在根毛發育調控發揮核心作用的三種激素中IAA信號通路研究最為清楚,ABA和ETH其次,其中關于ETH和IAA相互作用的研究以及ABA與根毛相關的研究經驗值得借鑒。此外,研究中也發現CTK也可能通過雙元組分系統和IAA、ETH相互作用[98],那么 CTK是否又會在脅迫改變水稻根毛表型中起著一定的作用,這些都是需要我們進一步了解的。
一般認為,在激素調控網絡下游的基因,尤其是負責細胞壁松弛基因如 OsEXPA17,負責細胞骨架構成的OsFHs,負責細胞壁多糖酶OsCSLD1及負責磷脂轉運的OsSNDP1等基因由于在激素下游,突變后是不能通過植物激素恢復它們根毛缺陷表型的,但在我們研究中發現OsFH1突變后在不同環境影響下,根毛表型有一定變化[9],因此在植物抗脅迫路徑中可能還有一些我們尚未關注到的植物激素或者激素以外的因素參與調控根毛表型[108-109]。與此同時,隨著測序技術的發展,水稻根部特異表達的基因也經過測序、GO富集分析篩選到一些可能參與根毛生長調控的基因,但這些基因的具體功能還需要通過遺傳學的研究加以驗證和分析[110]。
與擬南芥相比,無論是對于參與調控根毛發育基因的研究還是環境因子和激素在水稻根毛發育中的作用,相關研究都有很多空白。因此未來對于“環境-激素與遺傳因子-根毛形態發育”之間關系的研究大致可集中為以下幾個方向:1)探索在逆境脅迫條件下,ETH、ABA、IAA三者構成的復雜調控網絡中尚未發現的關鍵基因和調控通路;2)深入分析JA、BR、SL等在根毛發育方面研究相對較少的激素;3)進一步深入研究無法用已知的激素信號通路或者遺傳調控通路解釋的基因及環境因子對水稻根毛的調控作用。
猜你喜歡
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
軍事文摘(2021年22期)2021-11-26 00:43:51
小讀者(2021年2期)2021-03-29 05:03:48
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
華人時刊(2019年13期)2019-11-17 14:59:54
文苑(2018年22期)2018-11-19 02:54:14
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
