SCFAs對奶牛瘤胃上皮細(xì)胞Ca2+信號通路相關(guān)基因表達(dá)的影響
2020-07-22 07:57:32寧麗麗楊天宇姜茂成趙國琦
中國農(nóng)業(yè)大學(xué)學(xué)報 2020年8期
關(guān)鍵詞:信號
寧麗麗 成 健 詹 康 楊天宇 姜茂成 趙國琦,2,3*
(1.揚州大學(xué) 動物科學(xué)與技術(shù)學(xué)院,江蘇 揚州 225009;2.揚州大學(xué) 農(nóng)業(yè)科技發(fā)展研究院,江蘇 揚州 225009;3.揚州大學(xué) 教育部農(nóng)業(yè)與農(nóng)產(chǎn)品安全國際合作聯(lián)合實驗室,江蘇 揚州 225009;4.常熟市海虞動物防疫站,江蘇 常熟 215500)
短鏈脂肪酸(Short chain fatty acid, SCFAs)是瘤胃內(nèi)通過厭氧細(xì)菌發(fā)酵日糧產(chǎn)生的碳原子數(shù)為1~6的有機(jī)脂肪酸,主要包括乙酸、丙酸和丁酸,約占總揮發(fā)性脂肪酸95%[1]。SCFAs可部分代謝為酮體和乳酸,也可作為瘤胃上皮細(xì)胞的能量來源,反芻動物對SCFAs的依賴可以達(dá)到能量需求的70%[2]。G-蛋白偶聯(lián)受體41(G-protein-coupled receptor 41, GPR41)和GPR43已經(jīng)被證明是SCFAs的受體[3]。GPR41和GPR43在各個組織中均有表達(dá), GPR41在脂肪細(xì)胞中表達(dá)最高,GPR43在免疫細(xì)胞中表達(dá)最高[4]。并且GPR41和GPR43已經(jīng)在奶牛不同的組織中被鑒定,在奶牛瘤胃上皮細(xì)胞(BRECs)中GPR41的表達(dá)量高于GPR43[5]。GPR41和GPR43都可以與G蛋白家族的Gi/o蛋白和Gq蛋白偶聯(lián),從而增加細(xì)胞內(nèi)鈣離子濃度和降低cAMP濃度[7]。GPR41主要與Gi/o蛋白偶聯(lián)激活信號傳導(dǎo)系統(tǒng),而GPR43主要與Gq蛋白偶聯(lián)激活信號傳導(dǎo)系統(tǒng)[8-9]。
鈣離子參與機(jī)體的各項生理活動,不僅可以維持細(xì)胞膜兩側(cè)的生物電位,正常的神經(jīng)傳導(dǎo)功能,正常的肌肉伸縮與舒張功能以及神經(jīng)-肌肉傳導(dǎo)功能等,還可以發(fā)揮某些激素的作用機(jī)制。細(xì)胞內(nèi)的Ca2+濃度可以通過“G蛋白偶聯(lián)-PIP2-1,4,5-三磷酸肌醇(IP3)”信號途徑來調(diào)節(jié)[10]。磷脂酶C(PLC)和蛋白激酶C(PKC)是Ca2+信號通路中引起級聯(lián)反應(yīng)的關(guān)鍵分子[11]。PLCL1、PLCE1和PLCB2是PLC的3個家族,PKCB和PKCG是PKC的2種亞類。
在重組表達(dá)的GPR41或GPR43 CHO細(xì)胞中,SCFAs可以促進(jìn)IP3水平的上升以及cAMP含量的下降,并且還會導(dǎo)致MAPK的活化和Ca2+的釋放[4]。SCFAs刺激奶牛乳腺上皮細(xì)胞(BMECs)后,在2~4 s內(nèi)誘導(dǎo)細(xì)胞內(nèi)Ca2+濃度增加,60 s內(nèi)可以達(dá)到最大值。研究表明SCFAs是通過與GPR41、GPR43結(jié)合和活化而參與BMECs中的信號傳導(dǎo)[12]。此外,SCFAs可能通過GPR41和GPR43調(diào)節(jié)動物的神經(jīng)系統(tǒng)。如丙酸可通過GPR41加強(qiáng)交感神經(jīng)外流,但是在抑制GPR41的情況下丙酸并沒有加強(qiáng)交感神經(jīng)的活性。經(jīng)過siRNA試驗證明,交感神經(jīng)的激活是通過GPR41介導(dǎo)的[13]。有研究表明,奶牛瘤胃上皮細(xì)胞(BRECs)在缺乏SCFAs的條件下,GPR41不能被表達(dá)[14],SCFAs對BRECs 的Ca2+信號通路調(diào)控研究尚未有相關(guān)報道。因此,本研究旨在探討SCFAs對瘤胃上皮細(xì)胞的Ca2+信號通路相關(guān)分子的表達(dá)以及細(xì)胞內(nèi)Ca2+濃度的影響,為進(jìn)一步探究SCFAs對各種細(xì)胞Ca2+信號通路的影響提供依據(jù)。
1 材料與方法
1.1 試驗材料
DMEM高糖培養(yǎng)基、澳洲胎牛血清、非必需氨基酸(NEAA)、磷酸鹽緩沖液(PBS)和胰蛋白酶(Gibco,美國);乙酸、丙酸、丁酸、青霉素、鏈霉素、L-谷氨酰胺溶液和乙二胺四乙酸(EDTA)(Sigma,美國);PrimeScriptTMRT Master Mix和SYBR?Premix Ex TaqTM Ⅱ(TaKaRa,中國);熒光定量96孔板和8連管(Bio-rad,美國);總RNA提取試劑盒(Tiangen,中國);Fluo-4/AM(Beyotime,中國);FACS LSRFortessa流式細(xì)胞分析儀(BD,美國);試驗所用野生型BRECs和通過CRISPR/Cas 9系統(tǒng)敲除GPR41的BRECs均由揚州大學(xué)動物培養(yǎng)物保藏與應(yīng)用研究所(IACCA)提供[15]。
1.2 試驗方法
1.2.1試驗設(shè)計
試驗分為3個處理組,每個處理有3個重復(fù),第1組為正常野生型瘤胃上皮細(xì)胞;第2組為培養(yǎng)基中含20 mmol/L SCFAs野生型瘤胃上皮細(xì)胞;第3組為培養(yǎng)基中含20 mmol/L SCFAs且通過CRISPR/Cas 9系統(tǒng)敲除GPR41基因的瘤胃上皮細(xì)胞。放置37 ℃、5% CO2的培養(yǎng)箱中。24 h之后,消化并收集細(xì)胞進(jìn)行RNA提取。20 mmol/L SCFAs組成:12 mmol/L 乙酸、5 mmol/L丙酸和3 mmol/L丁酸。
1.2.2總RNA提取
按照總RNA提取試劑盒(Tiangen,中國)提取總RNA。最后,取1 μL提取的樣品進(jìn)行總RNA濃度和純度的測定。
1.2.3反轉(zhuǎn)錄成cDNA
按照Takara反轉(zhuǎn)錄試劑盒進(jìn)行,整個過程在冰上操作。反轉(zhuǎn)錄體系為10 μL,反應(yīng)條件:37 ℃ 15 min 和85 ℃ 5 s;PCR反應(yīng)體系為20 μL,反應(yīng)條件:95 ℃預(yù)變性30 s;95 ℃ 5 s,60 ℃ 30 s,40個循環(huán),每個樣品都有3個重復(fù)。
1.2.4Real-time PCR
熒光定量PCR反應(yīng)配置總體系為20.0 μL,其中SYBR?Premix Ex TaqTMⅡKit 10.0 μL;10 μmol/L 的PCR Forward primer和PCR Reverse Primer各0.8 μL;Water PCR grade 6.4 μL;cDNA 2.0 μL。引物詳情見表1。反應(yīng)條件:95 ℃預(yù)變性30 s;95 ℃ 5 s,60 ℃ 30 s,40個循環(huán),每個樣品都有3個重復(fù)。計算方法按照2-ΔΔCT。
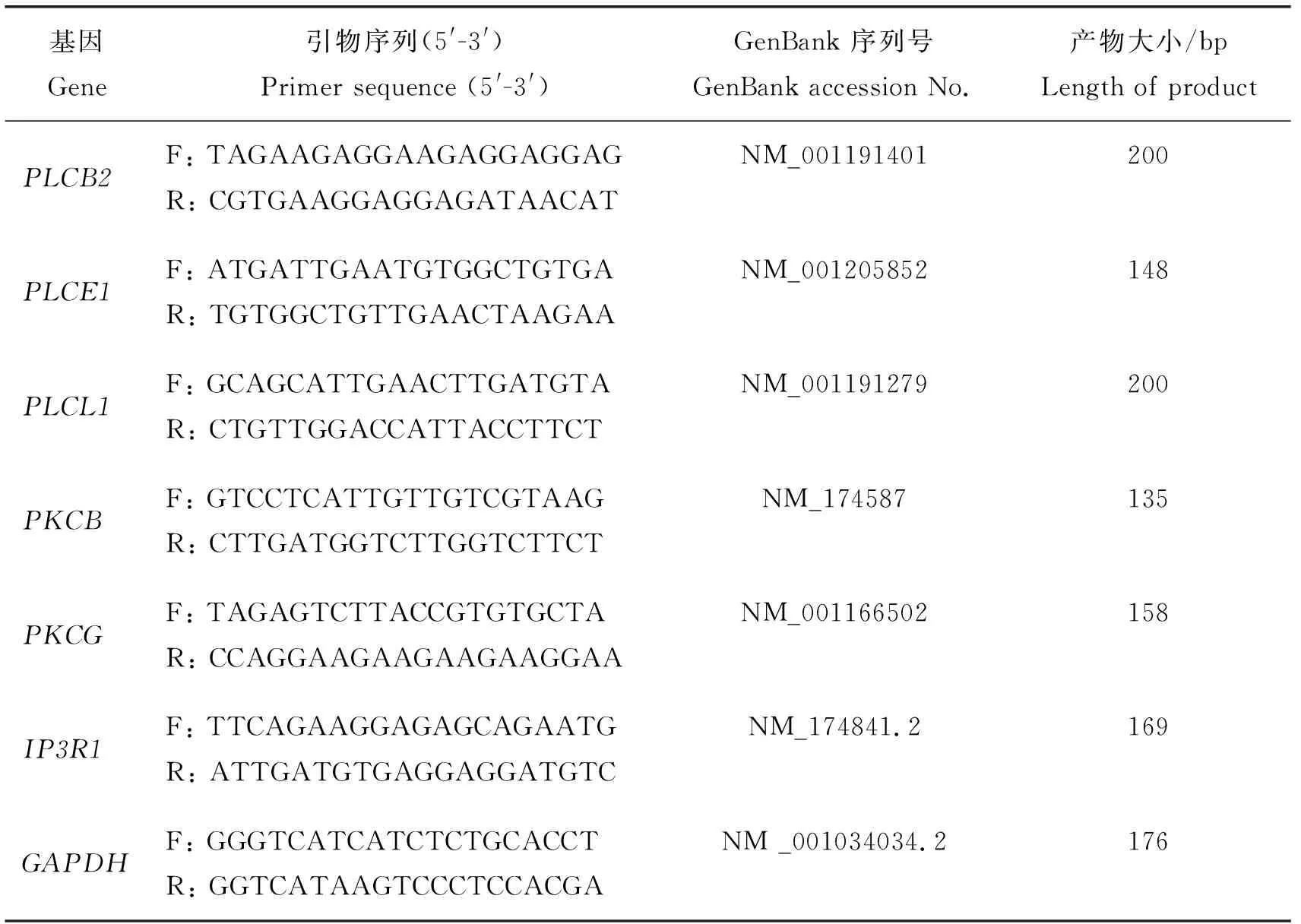
表1 熒光定量PCR引物Table 1 Reverse-transcription PCR primers
1.2.5細(xì)胞內(nèi)游離鈣離子濃度的測定
待接種細(xì)胞培養(yǎng)24 h后,第2和第3處理組分別加20 mmol/L SCFAs作用2 min。收集3組細(xì)胞,加Fluo-4/AM(終濃度2.5 μmol/L)37℃避光孵育30 min進(jìn)行熒光探針裝載,隨后PBS漂洗2次,流式細(xì)胞儀檢測。激發(fā)波長為494 nm,發(fā)射波長為516 nm,檢測的平均熒光強(qiáng)度來表示細(xì)胞內(nèi)Ca2+濃度。
1.3 數(shù)據(jù)統(tǒng)計分析
結(jié)果采用“平均數(shù)±標(biāo)準(zhǔn)差”表示。運用SPSS 16. 0統(tǒng)計軟件中的One-Way ANOVA模塊進(jìn)行單因素方差分析,顯著性檢驗應(yīng)用LSD法。P<0.05表示差異顯著,P<0.01表示極顯著差異。
2 結(jié)果與分析
2.1 SCFAs對BRECs中PLC表達(dá)的影響
采用qRT-PCR分別檢測PLCL1、PLCE1和PLCB2在不同處理的BRECs中的表達(dá)變化,結(jié)果見圖1。在缺乏SCFAs的條件下,PLCB2幾乎不被表達(dá);添加20 mmol/L SCFAs之后,PLCB2被極顯著激活(P<0.01),PLCL1和PLCE1表達(dá)量上調(diào),差異不顯著(P>0.05)。與添加SCFAs的野生型BRECs相比,敲除GPR41基因之后,PLCL1的表達(dá)量有下調(diào)的趨勢(P>0.05),而PLCE1和PLCB2的表達(dá)量極顯著增加(P<0.01)。

WT代表野生型BRECs;GPR41KO代表通過CRISPR/Cas 9系統(tǒng)敲除GPR41;數(shù)據(jù)標(biāo)注不同小寫字母表示差異顯著(P<0.05)。下同。WT stands for wild-type BRECs. GPR41KO stands for knocking out GPR41 through the CRISPR/Cas 9 system. Data annotations with different lowercase letters mean significant difference (P<0.05). The same below.圖1 SCFAs對BRECs中PLC的mRNA表達(dá)影響Fig.1 Effects of SCFAs on the abundance of PLC mRNA in the BRECs
2.2 SCFAs對BRECs中PKC表達(dá)的影響
PKCB和PKCG在不同處理的BRECs中的表達(dá)見圖2。與野生型相比,敲除GPR41之后,添加SCFAs可以上調(diào)PKCB和PKCG的表達(dá)量,但是差異不顯著(P>0.05)。
2.3 SCFAs對BRECs中IP3R1表達(dá)的影響
由圖3可知,促進(jìn)內(nèi)質(zhì)網(wǎng)鈣庫釋放的分子IP3R1在SCFAs誘導(dǎo)下,其mRNA表達(dá)量被顯著上調(diào)(P<0.05);與野生型BRECs比,敲除GPR41之后,添加SCFAs,IP3R1的mRNA表達(dá)量顯著下調(diào)(P<0.05)。
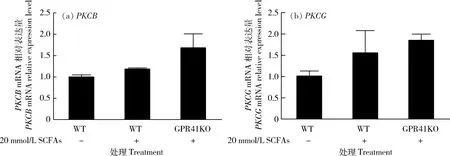
(a) SCFAs對PKCB的mRNA表達(dá)影響;(b) SCFAs對PKCG的mRNA表達(dá)影響。(a) Effects of SCFAs on the abundance of PKCB mRNA; (b) Effects of SCFAs on the abundance of PKCG mRNA.圖2 SCFAs對BRECs中PKC的mRNA表達(dá)影響Fig.2 Effects of SCFAs on the abundance of PKC mRNA in the BRECs

圖3 SCFAs對IP3R1的mRNA表達(dá)影響Fig.3 Effects of SCFAs on the abundance of IP3R1 mRNA
2.4 SCFAs對BRECs中Ca2+濃度的影響
從圖4可知,在野生型BRECs內(nèi)Ca2+濃度在SCFAs刺激后提高了21%,但差異不顯著(P>0.05);與野生型BRECs比,敲除GPR41之后,添加20 mM SCFAs細(xì)胞內(nèi)Ca2+濃度降低了15%,但差異不顯著(P>0.05)。
3 討 論
細(xì)胞內(nèi)鈣離子濃度的變化對細(xì)胞代謝和細(xì)胞增殖中的信息傳遞有非常重要的意義。細(xì)胞內(nèi)鈣信號是免疫細(xì)胞激活的關(guān)鍵因素,當(dāng)抗原受體被觸發(fā)時,鈣從細(xì)胞外流入細(xì)胞內(nèi)[16]。Kimura等[17]研究發(fā)現(xiàn),圍產(chǎn)期奶牛單核細(xì)胞內(nèi)鈣儲量較低,導(dǎo)致免疫細(xì)胞激活信號下鈣離子的釋放變?nèi)酢CFAs是反芻動物體內(nèi)主要的能量來源,并且可以調(diào)節(jié)瘤胃上皮細(xì)胞生長、瘦素水平、胰島素分泌和免疫應(yīng)答過程[18-19]。SCFAs還可以刺激交感神經(jīng)系統(tǒng)的激活,促進(jìn)機(jī)體能量消耗[13]。GPR41和GPR43已經(jīng)被證明是SCFAs的受體,奶牛瘤胃上皮細(xì)胞中GPR41的表達(dá)量高于GPR43[5]。有研究表明,添加20 mmol/L SCFAs和40 mmol/L SCFAs可以顯著增加BRECs中GPR41的表達(dá)量[6],而缺乏SCFAs的條件下,GPR41并不能夠被表達(dá)[14]。因此,本試驗通過在野生型瘤胃上皮細(xì)胞和敲除GPR41之后的瘤胃上皮細(xì)胞中添加SCFAs來檢測Ca2+信號通路中關(guān)鍵分子的表達(dá)量以及細(xì)胞內(nèi)Ca2+濃度的變化。
PLC和PKC是Ca2+信號通路中引起級聯(lián)反應(yīng)的關(guān)鍵分子。IP3R是促進(jìn)內(nèi)質(zhì)網(wǎng)鈣庫釋放的受體[11]。qRT-PCR結(jié)果表明,在缺乏SCFAs時,PLCB2幾乎不表達(dá),但是添加20 mmol/L SCFAs后顯著被激活,并且PLCE1和PLCL1表達(dá)量也有所提高,暗示PLC進(jìn)一步將IPI2分解為二酰甘油(DAG)和三磷酸肌醇(IP3)。IP3R是IP3受體,與IP3結(jié)合之后誘導(dǎo)其表達(dá)。因此,添加20 mmol/L SCFAs之后,IP3R1能極顯著激活。IP3R的表達(dá)介導(dǎo)內(nèi)質(zhì)網(wǎng)釋放Ca2+,添加SCFAs之后,細(xì)胞內(nèi)Ca2+濃度提高了21%。SCFAs刺激BMECs之后誘導(dǎo)細(xì)胞內(nèi)Ca2+濃度增加[12]。Yonezawa等[20]研究表明,GPR41和GPR43被SCFAs激活后,誘導(dǎo)細(xì)胞內(nèi)鈣離子濃度上升并且不受百日咳毒素的阻斷作用。丁酸與大鼠的腦垂體細(xì)胞在一起孵育后可以增加細(xì)胞內(nèi)Ca2+的濃度,這主要是通過GPR41和GPR43信號通路實現(xiàn)的[21]。SCFAs促進(jìn)Ca2+濃度的升高與本研究結(jié)果一致。細(xì)胞內(nèi)游離的Ca2+進(jìn)一步增加了PKC活化,添加SCFAs之后,PKCB和PKCG的表達(dá)量有所提高,和Ca2+濃度的變化趨勢相同。PKC進(jìn)一步激活MAPK信號通路,引起級聯(lián)反應(yīng),進(jìn)行細(xì)胞信號轉(zhuǎn)導(dǎo)。由此可以看出SCFAs激活細(xì)胞內(nèi)Ca2+信號通路與細(xì)胞內(nèi)MAPK信號通路是相輔相成的。丁酸不僅誘導(dǎo)細(xì)胞內(nèi)Ca2+濃度增加,而且也誘導(dǎo)GPR41和GPR43增加。在生長釋放素激素的作用下,GPR43基因沉默后,細(xì)胞內(nèi)的Ca2+濃度并沒有改變[12]。
為探究SCFAs對BRECs中Ca2+信號通路的調(diào)控機(jī)制,本文通過由揚州大學(xué)動物培養(yǎng)物保藏與應(yīng)用研究所提供的CRISPR/Cas 9系統(tǒng)敲除GPR41基因的BRECs來進(jìn)行深入研究。敲除BRECs中的GPR41基因,添加SCFAs之后,GPR41的表達(dá)量顯著下降,表明GPR41的一個等位基因被成功地敲除[11]。本試驗結(jié)果顯示,添加20 mmol/L SCFAs之后,與野生型BRECs比,敲除GPR41顯著上調(diào)了PLCB2和PLCE1的表達(dá)量;PLCL1的表達(dá)量降低,PKCB和PKCG的表達(dá)量升高,但是均無顯著差異。由此可以看出SCFAs不是通過GPR41加強(qiáng)PLCB2、PLCE1、PLCL1、PKCB和PKCG的表達(dá)。相反,IP3R1的表達(dá)量被顯著下調(diào),該結(jié)果表明SCFAs對IP3R1的調(diào)控是通過GPR41實現(xiàn)的。而且本試驗細(xì)胞內(nèi)Ca2+濃度降低了15%,是由于內(nèi)質(zhì)網(wǎng)上IP3R1的低表達(dá)抑制了內(nèi)質(zhì)網(wǎng)上鈣庫中Ca2+的釋放。
4 結(jié) 論
SCFAs可以通過激活其受體GPR41來調(diào)控BRECs上皮細(xì)胞IP3R1、PLCE1和PLCB2的表達(dá),及細(xì)胞內(nèi)Ca2+釋放,以進(jìn)一步激活Ca2+信號通路。
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
媽媽寶寶(2019年10期)2019-10-26 02:45:34
中國生殖健康(2019年3期)2019-02-01 06:12:26
鐵道通信信號(2018年11期)2019-01-19 01:15:08
電子制作(2018年11期)2018-08-04 03:25:42
鐵道通信信號(2018年2期)2018-04-18 12:18:10
鐵道通信信號(2016年11期)2016-06-01 12:11:32
鑿巖機(jī)械氣動工具(2016年3期)2016-03-01 04:00:25
中國病理生理雜志(2015年8期)2015-12-21 12:38:06
