外源褪黑素和脫落酸對干旱脅迫下葡萄生理特性的影響
2020-07-28 04:04:36王云梅周桂虹劉芯伶林芷儀
干旱地區農業研究 2020年3期
關鍵詞:植物
王云梅, 夏 惠, 周桂虹, 劉芯伶, 林芷儀, 梁 東
(1.四川農業大學園藝學院, 四川 成都 611130;2.汶川縣科學技術和農業畜牧局, 四川 阿壩藏族羌族自治州 623000)
干旱脅迫導致葡萄生長發育產生生理障礙,產量降低,漿果品質受到影響,是制約葡萄產業發展的重要環境因子之一[1]。褪黑素(melatonin,MT)是一種吲哚類小分子物質,作為一種廣譜的生理調節劑存在于絕大多數生物有機體中[2]。在植物體中,MT可以調節植物的生長發育[3],并通過清除活性氧和自由基的強抗氧化作用提高植物對干旱[4]、鹽害[5]、重金屬[6]、UV輻射[7]、高溫[8]、冷害[9]等多種逆境的抗性。脫落酸(abscisic acid,ABA)作為五大植物激素之一,主要起著抑制生長的作用;另一方面,ABA作為植物體細胞內信號傳導物質對非生物逆境響應發揮著重要作用。在植物的干旱脅迫信號傳導途徑中,有ABA-依賴型和ABA-非依賴型兩種途徑。ABA通過調節植株體內保護酶系統,增強其抵御活性氧毒害的能力,從而緩解干旱脅迫的傷害[10]。而外施ABA對植物的影響與ABA濃度密切相關[11-13]。目前,MT和ABA抗旱作用的研究方法多以PEG滲透脅迫模擬干旱為主,不能真正反映干旱這一自然過程;此外,MT和ABA兩種物質結合對干旱脅迫的效果及互作用關系還不清楚。
本研究以盆栽‘陽光玫瑰’葡萄扦插苗為材料,進行MT根灌、ABA噴施、MT+ABA混施和干旱處理,通過測定葡萄葉片MDA、H2O2、光合色素和滲透調節物質的含量以及抗氧化酶活性的變化,探究外源MT和ABA對葡萄干旱脅迫響應的生理機制以及MT和ABA之間的相互作用關系,以期為MT和ABA在果樹逆境中的生產應用提供理論依據。
1 材料與方法
1.1 試驗材料及處理
試驗材料為‘陽光玫瑰’(Shine-Muscat)葡萄扦插苗。于2017年冬季從四川農業大學試驗基地采集健壯的‘陽光玫瑰’1 a生枝條,沙藏處理。于2018年3月進行扦插,將枝條剪成長約5 cm左右、帶一個飽滿芽的枝段,扦插在裝有營養土、蛭石和沙子(體積比1∶1∶1)且直徑為13 cm、高度為15 cm的營養缽內。待生根后移入規格為(26 cm×22 cm)的塑料盆中,進行常規澆水施肥管理。
待扦插苗長至12~15片真葉時,選取長勢一致的‘陽光玫瑰’扦插苗60盆,平均分成5組,每組12盆,每盆1株,放入避雨棚進行處理:(1)正常供水對照(CK);(2)干旱對照處理(Dck);(3)100 μmol·L-1的MT根灌處理(MT);(4)50 μmol·L-1的ABA葉面噴施處理(ABA);(5)100 μmol·L-1MT處理和50 μmol·L-1ABA處理(MT+ABA)。首先,MT組和MT+ABA組用MT預處理:19∶00根灌500 ml的MT溶液,每2 d根灌1次,共4次;同時其它處理組澆500 ml清水。然后,對(2)~(5)組進行自然干旱處理,即不澆水;(1)組正常澆水,每2 d 1次,每次500 ml。ABA處理:在干旱處理第0、2、4、6、8 d和10 d的19∶00對ABA和MT+ABA處理組葉面分別噴施10 ml的50 μmol·L-1的ABA溶液;其它處理組噴施10 ml清水。在干旱脅迫處理的第0 d(以最后一次根灌MT溶液的第二天作為干旱脅迫第0d)、第18 d分別取各組植株4~8葉位的葉片,一部分用于測定相關生理指標,另一部分-80℃液氮速凍后保存于-80℃,用于后續指標的測定。
1.2 測定指標
1.2.1 土壤含水量和水勢及葉片相對含水量和水勢的測定 土壤含水量采用烘干稱重法測定;葉片水勢和土壤水勢使用型號為WP4C的水勢儀進行測定;葉片相對含水量的測定參照Zegaoui等[14]的方法。
1.2.2 丙二醛(MDA)、H2O2和光合色素含量以及滲透調節物質的測定 MDA和H2O2含量、相對電導率測定參照Bai等[15]的方法;滲透調節物質脯氨酸、可溶性糖和可溶性蛋白的含量測定參照李合生等[16]的方法;光合色素含量的測定參照Sariyeva等[17]的方法。
1.2.3 抗氧化酶活性的測定 超氧化物歧化酶(SOD)、過氧化物酶(POD)和過氧化氫酶(CAT)活性的測定采用Mirzaee[18]的方法。
1.3 數據處理及統計分析
數據均采用平均值±標準差(SD)表示,比較每個處理組平均值,以P<0.05為差異顯著,使用SPSS 20. 0進行單因素方差分析。
2 結果與分析
2.1 不同處理對干旱脅迫下葡萄形態特征的影響
干旱脅迫下植物葉片呈現出不同形態變化,發生葉片黃化卷曲、萎蔫等,脅迫嚴重時甚至葉片會脫落。從圖1可以看出,在控水處理18 d時,干旱對照處理組(Dck)的葡萄植株中、下部出現部分葉片脫落、萎蔫和黃化;MT處理組僅出現葉片黃化,無落葉;ABA處理組葡萄葉片脫落和黃化;MT+ABA處理組與MT處理組表型相似,葉片黃化。正常供水對照處理(CK)的葉片形態特征沒有發生明顯的變化。

圖1 干旱脅迫下‘陽光玫瑰’形態特征的變化Fig.1 The changes of morphological characteristics of ‘Shine-Muscat’ under drought stress
2.2 不同處理對干旱脅迫下土壤水分、葡萄葉片及根系水分的影響
表1數據表明,在控水條件下水分不斷蒸發,土壤含水量逐漸降低。控水18 d,干旱對照(Dck)處理的土壤含水量比正常供水處理降低了64.35%。土壤水勢的變化與含水量變化一致,呈顯著性降低,但各處理間無顯著性差異。
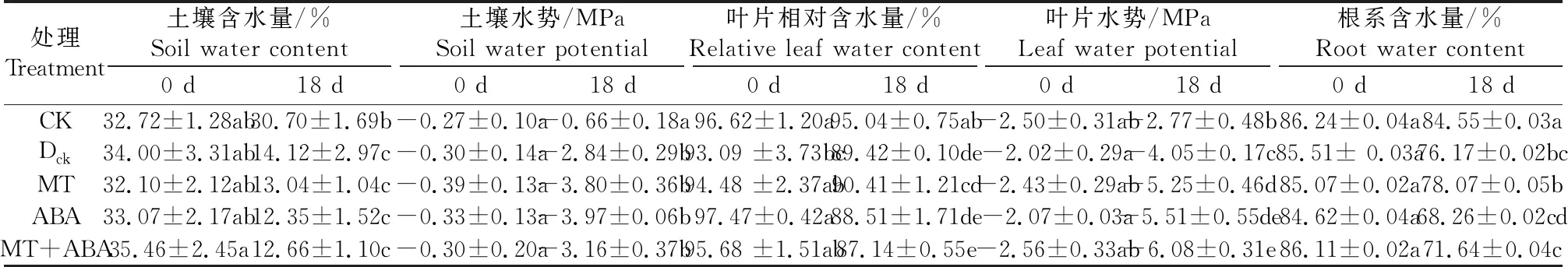
表1 不同處理對干旱脅迫下葡萄土壤含水量、土壤水勢、葉片相對含水量、葉片水勢和根系含水量的影響
干旱處理下Dck、MT、ABA和MT+ABA處理的葉片相對含水量均顯著低于對照(CK)處理。其中MT+ABA處理組最低,僅為CK處理的91.66%,其次是Dck和ABA組,兩者無顯著差異,MT組的葉片相對含水量高于Dck、ABA和MT+ABA。在干旱脅迫下葉片水勢較CK顯著降低。與干旱對照Dck相比,MT、ABA和MT+ABA處理進一步降低了葉片水勢,在MT+ABA處理下達到最低值-6.08 MPa。
干旱脅迫導致植株根系含水量顯著降低。控水處理18 d后,Dck、MT、ABA和MT+ABA處理組的根系含水量都顯著低于對照處理(CK);ABA和MT+ABA處理組的幼苗根系含水量顯著低于其他干旱處理組,分別達到68.26%和71.64%,說明ABA處理加速了葡萄根系中水分的散失。
2.3 不同處理對干旱脅迫下葡萄葉片膜脂過氧化程度的影響
干旱脅迫導致活性氧大量累積,造成細胞膜脂過氧化。由圖2A可知,干旱處理0 d時,MT和MT+ABA處理的葡萄葉片MDA的含量低于其他處理,但差異不顯著;正常供水對照處理(CK)的MDA含量在干旱脅迫18 d時顯著高于0 d。干旱處理18 d后的葡萄葉片MDA含量顯著高于正常供水處理;MT和MT+ABA處理的MDA含量卻顯著低于干旱對照處理,而ABA處理的MDA含量顯著高于其他處理,達到最高值,為20.49 μmol·g-1。
干旱處理0 d,ABA處理的‘陽光玫瑰’幼苗H2O2含量高于CK和MT+ABA處理,但CK、Dck、MT和MT+ABA之間差異不顯著。干旱處理18 d后,正常供水對照處理的H2O2含量顯著低于干旱處理0 d;Dck處理的H2O2含量顯著高于CK處理,達到66.10 mmol·L-1;MT、ABA和MT+ABA處理下H2O2含量顯著低于Dck處理,且ABA處理下的含量最低,為27.69 mmol·L-1(圖2B)。
植物在逆境脅迫下細胞膜透性增大,相對電導率升高。干旱處理0 d各處理的葉片相對電導率差異不顯著。干旱處理18 d后的CK處理與0 d無明顯差異;Dck、MT、ABA和MT+ABA處理的相對電導率高于CK對照處理,在ABA單一處理下的相對電導率最高,達到43.15%;MT處理和MT+ABA處理下相對電導率顯著低于Dck處理,緩解了干旱脅迫導致的電解質泄漏(圖2C)。
2.4 不同處理對干旱脅迫下葡萄光合色素的影響
干旱脅迫會導致植物葉片發黃失綠、萎蔫,甚至脫落。從圖3可以看出,干旱處理0 d不同處理葡萄幼苗葉片葉綠素a、葉綠素b、總葉綠素和類胡蘿卜素含量均無顯著差異。干旱處理18 d后,正常供水處理(CK)相比于0 d各光合色素含量顯著增加;干旱脅迫導致各光合色素含量分別下降了14.30%、25.04%、17.23%和20.67%,而在外源MT處理和MT+ABA處理下葉片各光合色素含量均顯著增加;ABA處理的各光合色素含量較低,但與MT和MT+ABA處理相比差異不顯著,說明外源施用MT有效抑制了干旱脅迫造成的葉片光合色素的降解。
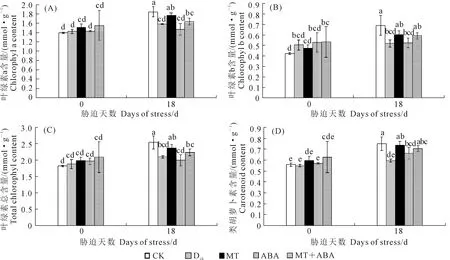
圖3 褪黑素和脫落酸對干旱脅迫下葡萄葉片葉綠素a(A)、葉綠素b(B)、總葉綠素(C)和類胡蘿卜素(D)含量的影響Fig.3 Effects of MT and ABA on chlorophyll a (A), Chlorophyll b (B), total chlorophyll (C), and carotenoid (D) contents in leaves of grape under drought stress
2.5 不同處理對干旱脅迫下葡萄滲透調節物質的影響
脯氨酸是植物體內非常重要的滲透調節物質,其含量的多少與植物抗旱性密切相關。由圖4A可以看出控水處理0 d時,經MT預處理的幼苗游離脯氨酸含量略高于其他處理,但差異不顯著。控水處理18 d,干旱處理導致葡萄幼苗游離脯氨酸含量升高了64.82%;MT處理和MT+ABA處理的游離脯氨酸含量顯著降低,在MT+ABA處理下達到最低值5.50 mg·g-1,而外源噴施ABA使其含量顯著上升,達到最大值15.41 mg·g-1。正常供水處理(CK)其脯氨酸含量與0 d相比差異不顯著。
干旱處理0d的葡萄幼苗葉片可溶性糖含量和可溶性蛋白含量均無明顯變化。干旱處理18 d時,正常供水(CK)處理的可溶性糖含量與0 d相比無顯著性差異,但可溶性蛋白含量降低了26.97%;干旱脅迫(Dck)導致幼苗可溶性糖含量和可溶性蛋白含量迅速增加,分別上升了25.69%和80.03%。在MT、ABA和MT+ABA處理下的可溶性糖含量顯著降低;MT+ABA處理的幼苗可溶性糖含量最低,為1.36 mg·g-1。經MT單獨預處理,其幼苗可溶性蛋白的含量最低,為251.03 mg·g-1(圖4B、4C)。

圖4 褪黑素和脫落酸對干旱脅迫下葡萄葉片游離脯氨酸(A)、可溶性糖(B)和可溶性蛋白(C)含量的影響Fig.4 Effects of MT and ABA on the contents of proline (A), soluble sugar (B), and soluble protein (C) in leaves of ‘Shine-Muscat’ under drought stress
2.6 不同處理對干旱脅迫下葡萄抗氧化酶活性的影響
在干旱脅迫下超氧化物歧化酶(SOD)、過氧化物酶(POD)、過氧化氫酶(CAT)作為保護細胞的重要酶類,能夠清除細胞內的活性氧。SOD參與植物體內的氧化代謝,能夠催化超氧陰離子發生歧化反應,生成O2和H2O2,保護細胞免遭逆境造成的過氧化傷害。從圖5可以看出,干旱處理18 d后,干旱對照處理(Dck)的SOD活性比正常供水處理CK降低了8.88%,而MT、ABA和MT+ABA處理的幼苗SOD活性顯著升高,且在MT+ABA處理下其活性顯著低于MT處理和ABA處理(圖5A)。
POD和CAT能夠通過分解H2O2從而對植物起到一定的保護作用。控水處理18 d,幼苗POD活性顯著高于正常供水(CK)處理,外源物質處理顯著增加了POD活性,且ABA處理下其活性最高,達到了17.91 U·min·mg-1;MT+ABA處理的POD活性顯著低于MT處理和ABA處理(圖5B)。由圖5C可以看出,干旱處理0 d,MT處理降低了CAT活性,干旱18d 時CAT活性相比于正常供水(CK)處理顯著升高了59.31%,MT、ABA和MT+ABA處理顯著降低了其活性,但3組處理之間差異不顯著。

圖5 褪黑素和脫落酸對干旱脅迫下葡萄葉片超氧化物歧化酶(SOD)(A)、過氧化 物酶(POD)(B)和過氧化氫酶(CAT)(C)活性的影響Fig.5 Effects of MT and ABA on the activities of SOD (A), POD (B) and CAT (C) activity in leaves of ‘Shine-Muscat’ under drought stress
3 討論與結論
干旱脅迫下不同外源物質處理會對植物缺水反應產生不同的影響,緩解或加重干旱的傷害,同時使用它們可能會產生協同或拮抗作用。因此,在本研究中,通過外源施用MT、ABA及其相互結合處理,評價與干旱適應相關的表型和生理指標的變化,對葡萄幼苗抗旱性及MT與ABA的相互作用關系進行了研究。
干旱脅迫下單獨或共同施用MT和ABA對葡萄幼苗的表型及生理特性有顯著的影響。如圖1表型所示,MT處理和MT+ABA處理減輕了干旱脅迫對葡萄幼苗葉片產生的黃化、脫落等毒性癥狀。研究表明光合色素很容易遭受逆境脅迫的傷害[19],本研究中MT處理增加了干旱脅迫下光合色素的含量(圖3),這與Kostopoulou等[20]對于MT的研究結果一致,說明MT削弱了植物體內的過氧化作用,緩解了干旱脅迫造成的損傷。50 μmol·L-1的ABA處理對光合色素的影響可能是由于對葡萄幼苗來說該濃度的ABA過高,對植物產生了毒害作用,降低了植物光合系統的穩定性和光合速率,從而阻礙了光合色素的合成或加速了光合色素的降解[12]。MT+ABA處理的作用與MT處理趨近,說明在MT+ABA處理下MT的促進作用緩解了ABA濃度過高所造成的傷害。
葉片水勢的高低和相對含水量的多少可以準確地反映干旱脅迫下植物葉片的水分狀況和受到土壤缺水的影響程度[21]。水分脅迫下,植物不僅要從低水勢的介質中吸收水分來維持植物體內的水分平衡,還要維持壓力勢基木不變以保證生理生化過程的正常運轉,所以通過降低滲透勢來進行調節是極其重要的一種方式,而滲透勢與細胞內多種滲透調節物質密切相關[22]。干旱脅迫下外源物質處理進一步降低了葉片的水勢(表1),可能是MT和ABA的施用雖然降低了脯氨酸、可溶性糖和可溶性蛋白等滲透調節物質的含量,卻大幅度提高了山梨醇、淀粉和蔗糖等其他滲透調節物質的水平[23],導致細胞滲透勢降低,從而降低了葉片水勢。葉君等[24]研究表明MT處理可增加干旱下小麥幼苗的根冠比,而根冠比增加有助于提高植物的吸水能力,維持植物組織含水量。適宜濃度ABA可以促進小麥幼苗的生長,而高濃度ABA則會產生抑制作用[25]。本研究中ABA處理降低了根系含水量,可能是ABA濃度過高對葡萄幼苗產生了抑制作用,加速了葡萄葉片水分的蒸騰,植物為了維持正常的生理代謝從而增強了地下根部水分向地上部的運輸。
脯氨酸、可溶性糖和可溶性蛋白作為重要的滲透調節物質,可以維持細胞狀態,最大限度地減少水分虧缺帶來的不利影響[26]。干旱脅迫導致葡萄幼苗滲透調節物質脯氨酸、可溶性蛋白和可溶性糖的含量顯著升高,這種變化是由于滲透調節物質的積累增強了葡萄幼苗自身的滲透調節水平以應對干旱脅迫[27]。外源MT處理降低了滲透調節物質的含量,維持了細胞內滲透調節水平的穩定。ABA處理顯著增加了游離脯氨酸的含量,其滲透調節物質整體水平高于MT處理,提高了滲透脅迫的整體水平,而MT+ABA處理的效果與MT處理相似(圖4)。由此推測,外源MT與ABA相互作用減輕了ABA濃度過高帶來的抑制作用,有效地調節了逆境下植物體內的滲透調節物質水平,維持了植物的正常生理代謝,從而提高了植物的抗旱性。
逆境會打破植物體細胞內自由基產生和清除的平衡,造成活性氧產生增加,膜透性增大,從而破壞細胞膜結構[28-29]。適宜濃度的MT可降低活性氧積累,減輕膜脂過氧化程度,提高植物氧自由基清除能力[30]。王娟[31]研究發現,適宜濃度的ABA能夠維持煙草幼苗葉片細胞內環境的穩定性,從而緩解干旱脅迫對煙草幼苗的損傷,而隨著干旱處理時間的延長,高濃度ABA噴施處理效果不明顯。本試驗中,雖然干旱脅迫增加了活性氧對植株造成明顯的損傷,但MT處理和MT+ABA處理能夠降低MDA和相對電導率水平,控制細胞膜的結構完整性,減輕干旱傷害。相反,ABA處理下MDA含量顯著增加,相對電導率升高(圖2),推測是由于ABA的濃度過高加重了幼苗的氧化損傷,這與趙春章等[32]的研究結果相似,其研究表明長期高濃度ABA(20 mg·L-1)處理不利于云杉幼苗生長,而MT清除活性氧的能力抵消或者部分減輕了ABA的效應。
植物葉片中抗氧化酶SOD、POD和CAT的協同作用可以有效清除植物體內多余的活性氧,共同抵抗干旱脅迫所造成的氧化損傷,在逆境脅迫中發揮著重要的作用[33]。已有研究表明,干旱脅迫下甘藍型油菜抗氧化酶活性提高,外源MT顯著提高了SOD和POD的活性[34]。適宜濃度ABA還可以誘導某些酶的重新合成從而提高植物的抗旱性[35]。在本研究中,干旱脅迫顯著降低了SOD活性,使POD和CAT活性升高,而外源MT處理通過提高SOD和POD活性,并降低CAT活性加強了酶促系統的抗旱能力,對清除植物體內多余活性氧的積累、減輕膜脂過氧化程度產生了一定的積極作用。ABA處理下抗氧化酶的變化與MT相似,這與本試驗中其他脅迫指標的變化相反,有可能是ABA濃度過高,長期處理對葡萄產生的消極影響超出了抗氧化系統防御能力的限度,致使SOD和POD的活性達到最高值。與其他脅迫相關指標的變化相似,MT與ABA結合處理下,兩者相互作用減弱了高濃度ABA造成的傷害,同時削弱了MT對于干旱脅迫的作用效果,但其具體的作用機理有待于進一步研究。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
