喀斯特地區四種草本植物及其基質的優先選擇*
2020-08-25 03:37:50陳海霞黃宗勝趙明偉李祖國劉逸夫張麗欣楊春楊政蓉張元博
土壤學報 2020年4期
陳海霞黃宗勝?趙明偉李祖國劉逸夫張麗欣楊 春楊政蓉張元博
喀斯特地區四種草本植物及其基質的優先選擇*
陳海霞1,黃宗勝2?,趙明偉2,李祖國1,3,劉逸夫1,張麗欣2,楊 春1,楊政蓉1,張元博2,4
(1. 貴州大學林學院,貴陽 550025;2. 貴州大學建筑與城市規劃學院,貴陽 550025;3. 遵義市園林管理站,貴州遵義 563000;4. 貴州工商職業學院人文體育學院,貴陽 551400)
為弄清植物對喀斯特基質的優先選擇,用盆栽法研究了四種植物對喀斯特不同基質的適應性。結果表明:整體上隨植物生長,基質理化性質除鉀含量降低外,其他均增加;發芽率和存活率以石灰土、<1 mm砂、<1 mm砂+石灰土最高,碎石、碎石+枯落物最低,白三葉草發芽率最高、狗牙根存活率最高;株數、株高、蓋度以石灰土、<1 mm砂、<1 mm砂+枯落物高而碎石、碎石+枯落物低,且蓋度均呈升高趨勢;生物量和葉綠素含量狗牙根和白三葉草高而芒草和高羊茅較低,生物量以石灰土、<1 mm砂、<1 mm砂+枯落物高而碎石、碎石+枯落物低,葉綠素含量以<1 mm砂+石灰土、石灰土+1 mm~5 mm砂高而碎石低;δ13C值芒草和狗牙根高而白三葉草和高羊茅低,且δ13C值與植物及生長基質有關。綜上,四種植物對石灰土、<1 mm砂、<1 mm砂+枯落物適應性強,對碎石、碎石+枯落物適應性差;狗牙根和芒草適應性最強,白三葉草次之,高羊茅最差,喀斯特生態修復可優先選擇芒草、狗牙根。
喀斯特;草本植物;基質;適應性;生態恢復
當前喀斯特區石漠化問題已成為該區經濟和生態可持續發展的瓶頸,石漠化的生態恢復和重建亦成為研究熱點。喀斯特區生態環境惡劣,土層淺薄且不連續,成土速率慢[1],由于土層淺薄和巖石的高滲透性[2],喀斯特生境中不同基質普遍存在不同程度的水分虧缺,而基質的持水能力及其理化特性會影響植物生長,且植物對不同基質的適應性亦不同,因此開展不同植物對喀斯特生境中不同基質的適應性研究,能夠為喀斯特區植被恢復初期提供物種選擇及其生長基質優劣判定提供理論依據,對喀斯特石漠化生態系統恢復重建具有指導意義。
適應是生物在環境中沒經過生存競爭而形成的一種適合環境條件的特性,它是自然選擇的結果,適應性是指生物適應環境的程度[3]。近年來,國內學者從不同的角度研究了植物對喀斯特生境的適應性,包括干旱脅迫中的光合特性[4]、生理機制[5]、植物水分利用方式[6]及植物對高鈣的適應性[7]。此外,對喀斯特石漠化區的典型植物生理生態適應性的研究報道也比較多[8],但是現有的研究僅僅揭示了植物對石漠化逆境的適應方式與對策,其對各種極端生境類型低生態容量的適應性缺乏研究。土壤是地球陸地表面能生長綠色植物的疏松表層[9],基質是植物、微生物從中吸取養分借以生存的物質[10-11],土壤是植物生長的重要基質[12]。目前關于植物對喀斯特生境基質的適應性成果多體現在植物對土壤的適應性研究,植物對喀斯特生境其他生長基質的適應性研究卻鮮有報道。喀斯特生境中基質種類復雜多樣,土壤只是其中一種,目前喀斯特區對于喀斯特土壤質量[13]、土壤有機碳[14]、土壤養分的空間異質性及形成機制[15]、土壤保墑能力[16]、土壤養分含量變化規律及其影響因素[17]和碳匯[18]等方面進行了較為系統的研究,但是對于生長基質,如砂、石、枯落物等的研究卻鮮有報道。基質作為植物生長的重要載體,其理化特征對植物生長起到決定性作用,不應僅僅局限于研究土壤這單一基質,應該拓寬研究對象、研究思維和研究視角,重視所有能夠供植物生長的基質研究,這樣才能夠符合喀斯特生境多樣性的需要,才能為喀斯特區植被恢復以及生態系統的恢復重建提供理論依據。草本植物階段是石漠化區植被恢復的初期階段,也是最主要的階段[19]。鑒于此,本文選擇4種草本植物(包括2種C4植物和2種C3植物)為研究對象,從植物形態、發芽率和存活率、δ13C值、光合色素和生物量分配以及基質理化特性等方面,對喀斯特生境中生長基質組成,其理化性質以及草本植物對其的適應性進行研究,旨在揭示喀斯特生境中基質特征,得出植物適應性特征,為喀斯特石漠化區生態系統恢復與重建提供理論依據。
1 材料與方法
1.1 試驗區概況
本試驗于2018年7月23日在貴州省溪北社區養牛村(26°27′N,106°38′E)開始實施,該區森林覆蓋率為35%,海拔為1 178 m,氣候類型為亞熱帶季風濕潤氣候,試驗期間平均氣溫為19.5℃,降雨量為361.3 mm,無霜期為148 d,屬喀斯特區,其土壤為石灰土、黃壤。
1.2 基質主要類型及其選擇
通過實地調研與文獻查閱發現,喀斯特生境中供試基質類型多樣,主要有石灰土、砂、石、枯落物等,其中枯落物層厚度約2 cm。貴州喀斯特石漠化嚴重,砂石含量大,有些地區甚至全是砂石,而土壤極少,因此本文選擇石灰土、砂、石、枯落物及其混合物作為研究基質。喀斯特生境基質構成可能是一種或多種組分混合,因此,通過采集野外不同喀斯特生境基質,對其進行篩選分類,具體如下:將取自貴陽市花溪區喀斯特生境中的基質樣品帶回實驗室,通過野外調研發現喀斯特生境中有純的石質砂石,且有植物生長于其中,因此在進行基質組分分離時并未單獨按照土壤學中砂石粒徑進行劃分,而是按照土壤學與土木建筑相結合的標準進行砂、石、土粒徑劃分,簡述如下:將采回的樣品去除植物根系并過1 mm和5 mm篩,將其按照砂、石、土進行篩分,其中粒徑>5 mm的為碎石,粒徑<5 mm的為砂(即石質的砂)[20],并將砂分為<1 mm砂和1 mm~5 mm砂,碎石與1 mm~5 mm砂用干篩法將其與土壤、<1 mm砂分離(過篩前把肉眼可見的大于1 mm的土質顆粒捏碎),<1 mm砂和土按照吸管法分離[21];總體上將基質分為土、<1 mm砂(簡稱Ⅰ砂)、1 mm~5 mm砂(簡稱Ⅱ砂)、<5 mm的碎石(簡稱碎石)四種組分,篩分后稱重,由于有些基質樣品中存在分解或半分解的枯落物,這種分解或半分解的枯落物一方面其質量極小而無法與砂、石、土等的質量進行對比,另一方面篩分非常困難,故未將其分離出來,在分析基質組分質量比時亦未將其計入,但由于基質中有分解或半分解的枯落物,這對植物生長非常有利,是這些基質的組成部分,尤其是枯落物基質是由分解或半分解枯落物與未分解枯落物相混合而構成,故這些基質類型命名時仍有枯落物,所得基質類型及其構成見表1。表1顯示:混合基質中含有土、砂、石、枯落物四種組分,有的含有其中一種,有的是多種混合,通過組分構成分析,共有19種基質類型。
1.3 試驗材料
通過多年野外調查及查閱貴州植物志等相關文獻,發現狗牙根()、芒草()、高羊茅()和白三葉草()為貴州喀斯特區常見的草本植物,廣泛分布于各種類型的生境中,具有較強的代表性,因此本文選擇該4種草本植物為研究對象,其中狗牙根和芒草為C4植物,白三葉草和高羊茅為C3植物。試驗草種均購買于貴州貴陽,種子均為同一年采摘,花盆口徑為16.8 cm、高14 cm,所有試驗基質取自貴陽市花溪區(26°11′N,106°27′E)喀斯特山地。
1.4 試驗設計
本文栽培試驗基質按照表1中19種基質的四種組分質量構成進行試驗配制(取樣時,有分解或半分解枯落物的基質,由于分解或半分解枯落物質量極少,則按野外調查原位取樣,這樣確保基質中有分解或半分解枯落物),試驗所用砂石取于石質生境,所取土壤為石灰土,由于野外調研發現有分解或半分解枯落物的基質上均覆蓋有2 cm厚未分解的枯落物,為了保持原真性,試驗時在有分解或半分解枯落物的基質上鋪置2 cm厚未分解枯落物,所取基質組分均來自表1中野外調研時的原樣地。試驗于2018年7月23日—2018年12月15日進行,一共4種植物,19種基質類型,3個重復。播種前,篩選草種,去除有雜質、蟲蝕、裂口以及成熟度低的種子。將各基質按組分質量比混合配置完成,并將其裝進花盆中至同一高度,將挑選好的植物種子按照每盆100粒均勻撒在盆里的淺基質層(預試驗顯示喀斯特植物生長基質較土壤基質難以適合植物生長,試驗顯示以較密的100粒較為合適),在有枯落物的基質上鋪置2 cm厚未分解枯落物,所有花盆裝的基質最后高度一致。從播種開始,視生長基質干濕程度澆水,每次每盆澆透(約400 mL),生長過程中不添加任何營養液,也不噴灑農藥。播種后每天觀察種子的發芽數并記錄,以種植21d后的發芽數計算發芽率(通過觀察21d時的發芽數基本穩定)。

表1 喀斯特生境中基質類型及其構成

續表
注:為了便于稱呼,本文取基質簡稱,便于圖表的表達,下文同。Note:For the convenience of appellation,this paper uses abbreviations in referring to the matrices,for the convenience of chart expression,the same below.
1.5 基質理化性質的測定
基質pH采用pH計測定[22],全氮(Total nitrogen,TN)采用擴散法測定[23]、全磷(Total phosphorus,TP)采用硫酸—高氯酸消煮法測定[23]、全鉀(Total potassium,TK)采用火焰光度法測定[23],持水量(Water holding capacity,WHC)和氣體孔隙度(Gas porosity,AP)參照穴盤苗生產原理與技術測定[24]。
1.6 植物相關指標的測定
以播種21 d時的發芽數計算發芽率,8月26日起每月26日測定植物株高、株數、蓋度。在試驗的第4個月根據Lichtenthaler[25]的方法測定總葉綠素(Total chlorophyll,Chl a+b)和類胡蘿卜素(Carotenoid,Car)的含量,并計算類胡蘿卜素與葉綠素含量的比值(Car/Chl a+b)。在試驗的第5個月進行δ13C值測定,將采集的葉片經過清洗、烘干、冷卻、粉碎后,過20目篩密封保存。將處理好的植物樣品送自然資源部第三海洋研究所,用元素分析儀-穩定同位素質譜儀聯機(Vario ISOPOTE Cube-Isoprime,Elementar公司)測定樣品中δ13C值。并以PDB國際標準作為參考標準,δ13C值按通用標準公式計算[26]:
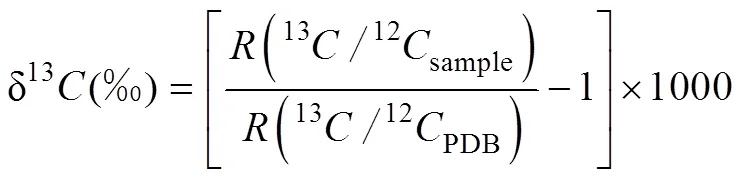
式中,(13C/12CPDB)為國際標準物PDB(Peedee Belemnite)的碳同位素豐度比值。
試驗結束時測定生物量。將每盆植物收割并將地上部分與地下部分分開,分別測量植物地上、地下鮮重,并在85℃下烘干至恒重并稱量其干重,計算地上部分與地下部分生物量比值與根重比(根干重/全株干重(Root weight ratio,RMR))。
1.7 數據處理
通過Microsoft Excel 2017、SPSS 17.0軟件對數據進行統計分析,采用單因素方差分析(one-way ANOVA)和LSD法檢驗各基質之間的差異顯著性,用雙因素方差分析(two-way ANOVA)檢驗基質類型和植物類型與各指標之間的差異,顯著性水平設定為α=0.05。
2 結 果
2.1 喀斯特生境的基質類型與特征
表1表明喀斯特生境基質組成多樣,按石、砂、土、枯落物的構成喀斯特生境基質類型可分為19種(為便于稱呼,本文取其簡稱,見表1,下文同),分別為石灰土、碎石、Ⅱ砂、Ⅰ砂、枯落物、砂石混、土枯、碎石枯、Ⅱ砂枯、Ⅰ砂枯、砂石混枯、土砂石混、土碎石、土Ⅱ砂、土Ⅰ砂、土砂石混枯、土碎石枯、土Ⅱ砂枯、土Ⅰ砂枯。在混合基質中,土的占比較小,碎石占比大。由于喀斯特生境的獨特性,有些基質上層有約2cm未分解枯落物。
2.2 植物種植前后基質理化性質的變化
由圖1可知整體上,植物種植后基質總孔隙度、氣體孔隙度(AP)、持水量(WHC)、水氣比、pH、全氮(TN)、全磷(TP)均增高,全鉀(TK)含量降低,基質呈堿性或強堿性;其中,白三葉草和狗牙根生長基質的理化性質變化較為明顯,高羊茅和芒草的變化不大;枯落物的總孔隙度、AP、水氣比增加,但是持水量降低,說明植物種植后,枯落物的透氣性增強,空氣容量大,但是保水能力下降;Ⅱ砂、碎石枯、土Ⅱ砂的總孔隙度、AP、WHC、水氣比與種植前相比下降,其余基質基本升高,說明植物對基質理化性質產生一定影響,種植植物后使得基質的透氣性增強,空氣容量增大,保水能力也有所增強;基質TK含量下降,說明植物生長過程中吸收TK,以促進生長發育。
2.3 四種草本植物在不同基質中的發芽率與存活率
整體上,四種植物的發芽率和存活率在石灰土、Ⅰ砂、Ⅱ砂、Ⅰ砂枯、土Ⅰ砂中高,其次為土枯、土砂石混、土Ⅱ砂枯、土Ⅰ砂枯、土砂石混枯,碎石枯、砂石混枯、土碎石和土碎石枯中較低,碎石中最低,可能是由于碎石粒徑大,總孔隙度大,持水量低導致保水性弱,不利于種子萌發與存活;白三葉草發芽率最高,其次為高羊茅、芒草,狗牙根發芽率最低,狗牙根的存活率最高,其次是芒草、白三葉草,高羊茅最低。
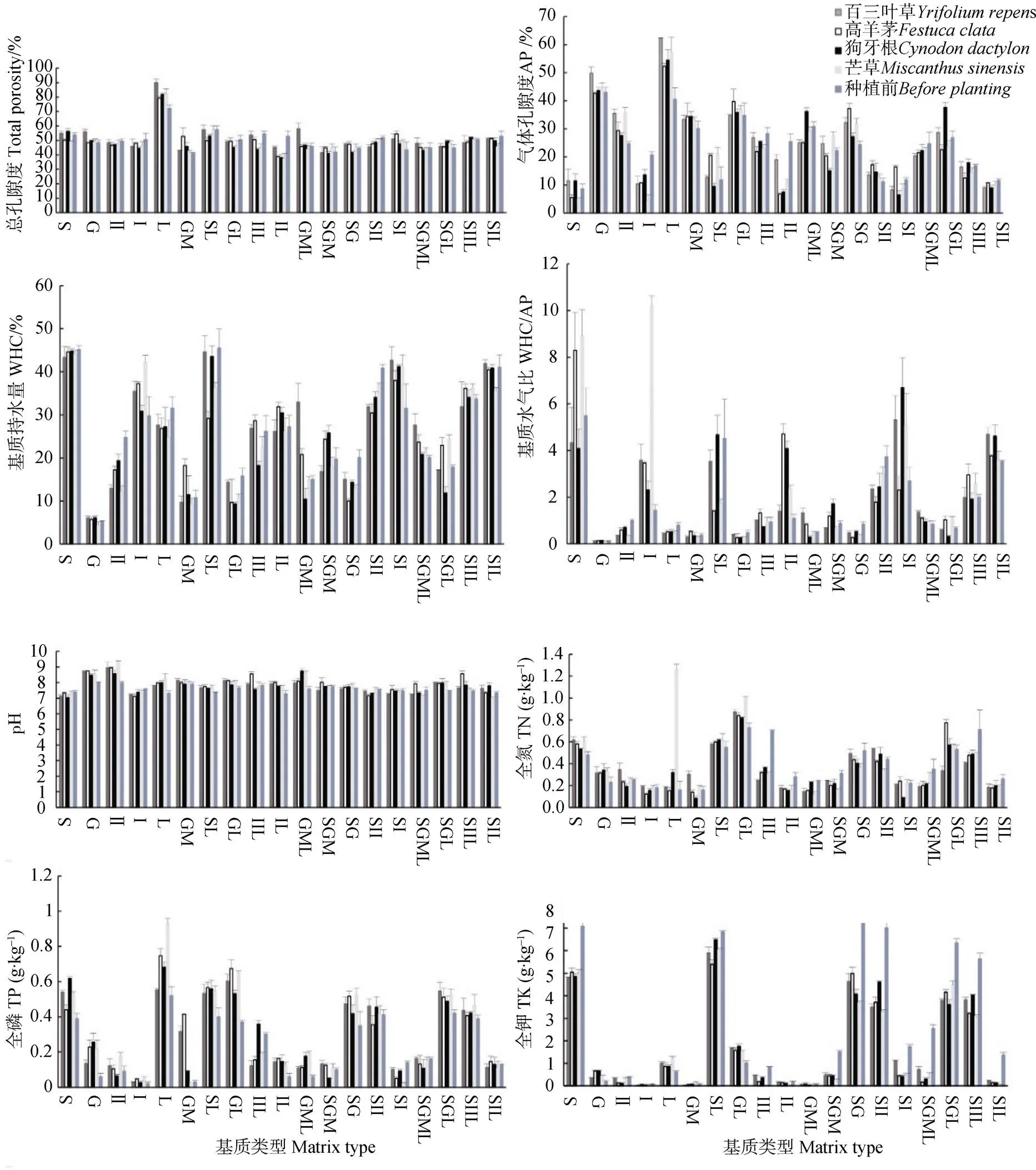
圖1 植物種植前后基質理化性質對比
2.4 植物形態對喀斯特生境基質的適應性
整體上,隨著種植時間的變化,白三葉草、高羊茅和芒草的株數逐漸減少,狗牙根株數逐漸增多最后基本保持不變,其中高羊茅株數變化最大,可能與高羊茅是冷季型草坪草,不適應高溫氣候有關,白三葉草、狗牙根和芒草的株數變化較小;白三葉草、狗牙根和芒草在石灰土、Ⅱ砂、Ⅰ砂、Ⅰ砂枯、土枯和土Ⅱ砂枯中株數變化較小,在枯落物、Ⅱ砂枯、砂石混枯中變化較大,在碎石和碎石枯中植物株數最少。
整體上,狗牙根株高呈上升趨勢,白三葉草和芒草株高呈先升高后下降,而高羊茅的株高則變化多樣,在多種基質中呈先增高后降低再升高,其中在石灰土和枯落物中呈降低再升高,可能是由于季節變化,植物枯黃甚至枯死,導致高度降低;四種植物在枯落物、Ⅰ砂枯、土Ⅰ砂和土Ⅰ砂枯中的株高最高,石灰土、Ⅰ砂、土枯、Ⅱ砂枯、砂石混枯和土砂石混枯中較高,Ⅱ砂、砂石混和土碎石中較低,碎石和碎石枯中最低,說明植物不適應于碎石和碎石枯。
由圖2可知整體上,隨時間變化,各基質中白三葉草、狗牙根和芒草的蓋度逐漸升高,高羊茅的變化多樣,在大多數基質中呈迅速下降再逐漸升高,在枯落物、土枯、砂石混中升高,這說明高羊茅對不同基質的適應性不同;白三葉草蓋度最高,其次為芒草、狗牙根,高羊茅最低;四種植物蓋度在Ⅰ砂、Ⅱ砂枯、Ⅰ砂枯和土Ⅰ砂中最高,石灰土、枯落物、土枯、土Ⅱ砂、土Ⅱ砂枯中較高,Ⅱ砂、砂石混、砂石混枯中較低,碎石和碎石枯中最低,說明四種植物不適合在碎石和碎枯中生長。
2.5 四種草本植物在不同基質中的生物量
圖3顯示整體上,四種植物在Ⅰ砂、枯落物、Ⅰ砂枯和土碎石枯中地上生物量與地下生物量之比最大,石灰土、Ⅱ砂、砂石混枯、Ⅱ砂枯中較大,砂石混、土Ⅰ砂、土Ⅱ砂枯和土Ⅰ砂枯中較小,碎石和土碎石中最小;其中狗牙根的最大,其次為三葉草、高羊茅,芒草最小,說明狗牙根主要采取地上部分生長的生長策略;四種植物的RMR在Ⅱ砂、砂石混、土枯、砂石混枯和土Ⅱ砂枯中最大,石灰土、枯落物、土砂石混、土Ⅱ砂、土Ⅰ砂和土砂石混枯中較大,Ⅱ砂枯、Ⅰ砂枯和土碎石枯中較小,碎石、Ⅰ砂中最小;芒草的RMR最大,其次為高羊茅、三葉草,狗牙根最低,說明芒草的根系發達,主要采取地下部分生長的生長策略。
2.6 植物光合色素對斯特生境中基質的適應性
圖4表明四種植物的Chl a+b在土砂石混、土Ⅱ砂、土Ⅰ砂、土砂石混枯和土Ⅰ砂枯中高,其次為石灰土、Ⅱ砂、Ⅰ砂、土枯和Ⅰ砂枯,碎石中最低;其中白三葉草的Chl a+b含量最高,其次為狗牙根和芒草,高羊茅最低,說明白三葉草和狗牙根吸收和轉化光能的作用強,更有利于光合作用;四種植物的Car/Chl(a+b)在Ⅰ砂、Ⅱ砂、枯落物、Ⅰ砂枯、土Ⅱ砂枯和土Ⅰ砂枯中大,土枯、砂石混枯、土砂石混枯和土Ⅰ砂中較大,土碎石枯和土碎石中較小,碎石中最小;狗牙根的Car/Chl(a+b)最大,其次為芒草、三葉草,高羊茅最小,說明狗牙根和芒草的光保護系統更容易增強,以能夠適應惡劣生境。
2.7 四種草本植物在不同基質中的水分利用特性
表2表明整體上,芒草的δ13C值最高,其次為狗牙根、白三葉草,高羊茅最低,說明芒草和狗牙根水分利用效率高,白三葉草和高羊茅低;白三葉草的δ13C值在Ⅰ砂、土砂石混、土砂石混枯和碎石枯中大,石灰土、Ⅱ砂和Ⅱ砂枯中較大,土碎石和土Ⅱ砂枯中較小,碎石、土Ⅱ砂和土碎石中最小;芒草的在砂石混中最大,土Ⅱ砂和土砂石混中較大,土碎石和土Ⅱ砂枯中較小,Ⅰ砂枯中最小;狗牙根的在Ⅰ砂中最大,土Ⅰ砂和土砂石混枯中較大,Ⅰ砂枯、土碎石和土砂石混中較小,Ⅱ砂中最小;高羊茅的在枯落物中最大,土Ⅰ砂和土砂石混中較大,砂石混枯和土碎石中較小,土枯中最小;以上表明植物葉片δ13C值跟植物種類與生長基質有關;說明不同植物、不同生長基質,植物水分利用效率不同,反映出喀斯特生境的復雜性及植被恢復的艱巨性。
3 討 論
四種草本植物對喀斯特不同生長基質的適應性表明:植物可以生長在非土壤的石質基質中,并且非傳統意義上如泥炭、草炭、蛭石等基質[27],而是取決于喀斯特區自然狀態下的生長基質。同時表明試驗基質中存在植物生長所必需的營養元素,如氮、磷、鉀等,圖1表明了19種基質中砂質基質的TN、TP、TK含量低于含土壤的基質。綜上所述,喀斯特生境中生長基質的概念可概括為喀斯特生境中能夠為種子萌發或者植物生長提供條件的一種或者多種物質混合的介質。其內涵特征是有的營養元素含量(氮磷鉀)較低,有的營養元素含量較高,類型豐富多樣(如本研究得出了19種類型),有土、少土、極少土或無土,且非土壤生長基質占比高,基質pH呈中性或偏堿性,極少數偏酸性。經植物物種、基質類型雙因素方差分析表明:不同植物物種對試驗結果中的發芽率、存活率、株數、株高、蓋度、根重比、葉綠素含量等指標數值大小影響均存在顯著差異(<0.05),不同基質類型對上述試驗指標亦存在顯著差異(<0.05),同時植物物種與基質類型還存在交換效應(<0.05),且其交互效應對上述指標亦存在顯著差異(<0.05)。這說明在喀斯特區植物物種的選擇及基質類型是喀斯特生態系統恢復的兩個重要因素,在將來的理論與實踐中應該同時引起足夠重視。
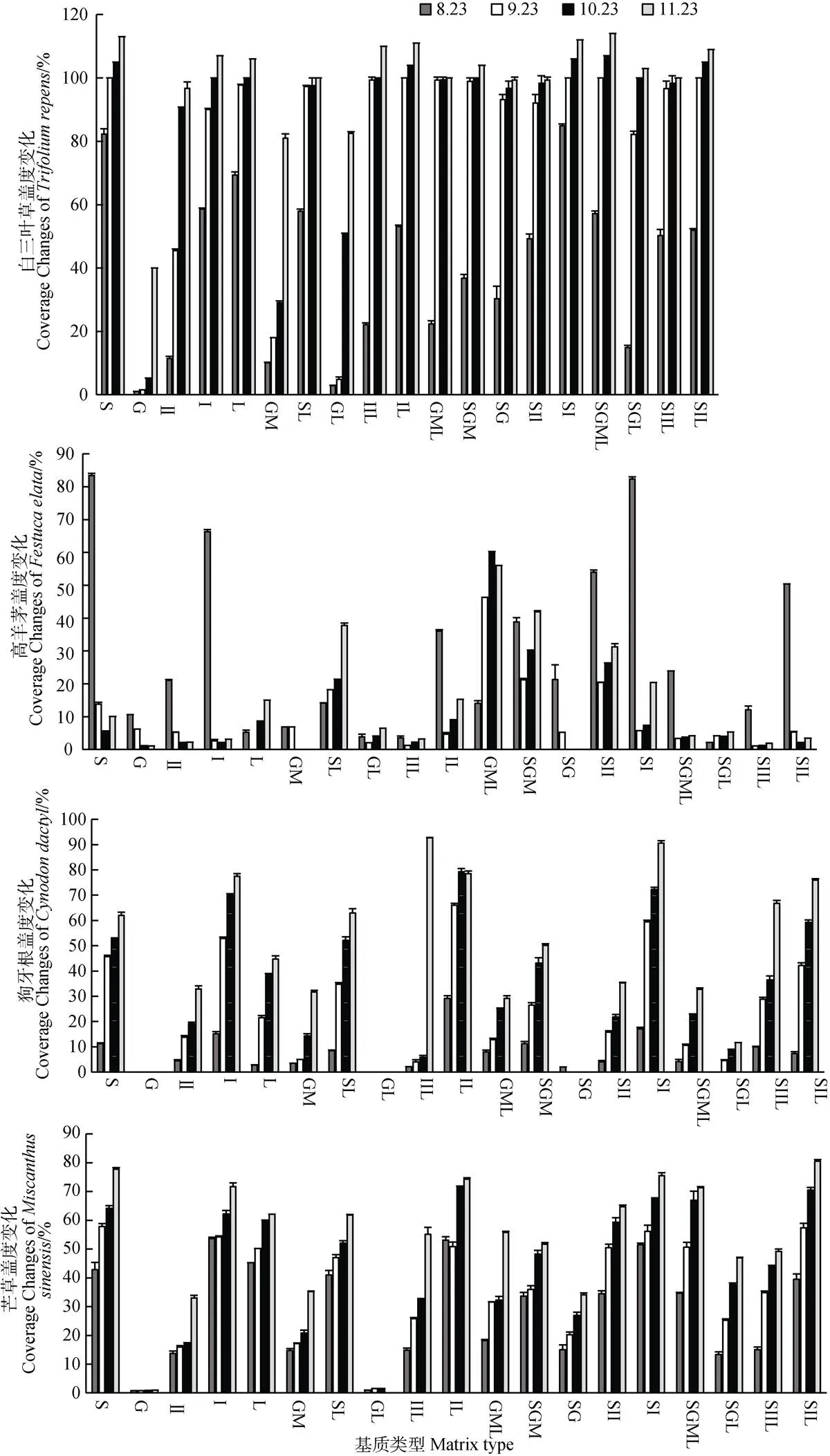
圖2 四種植物在不同基質中的蓋度變化
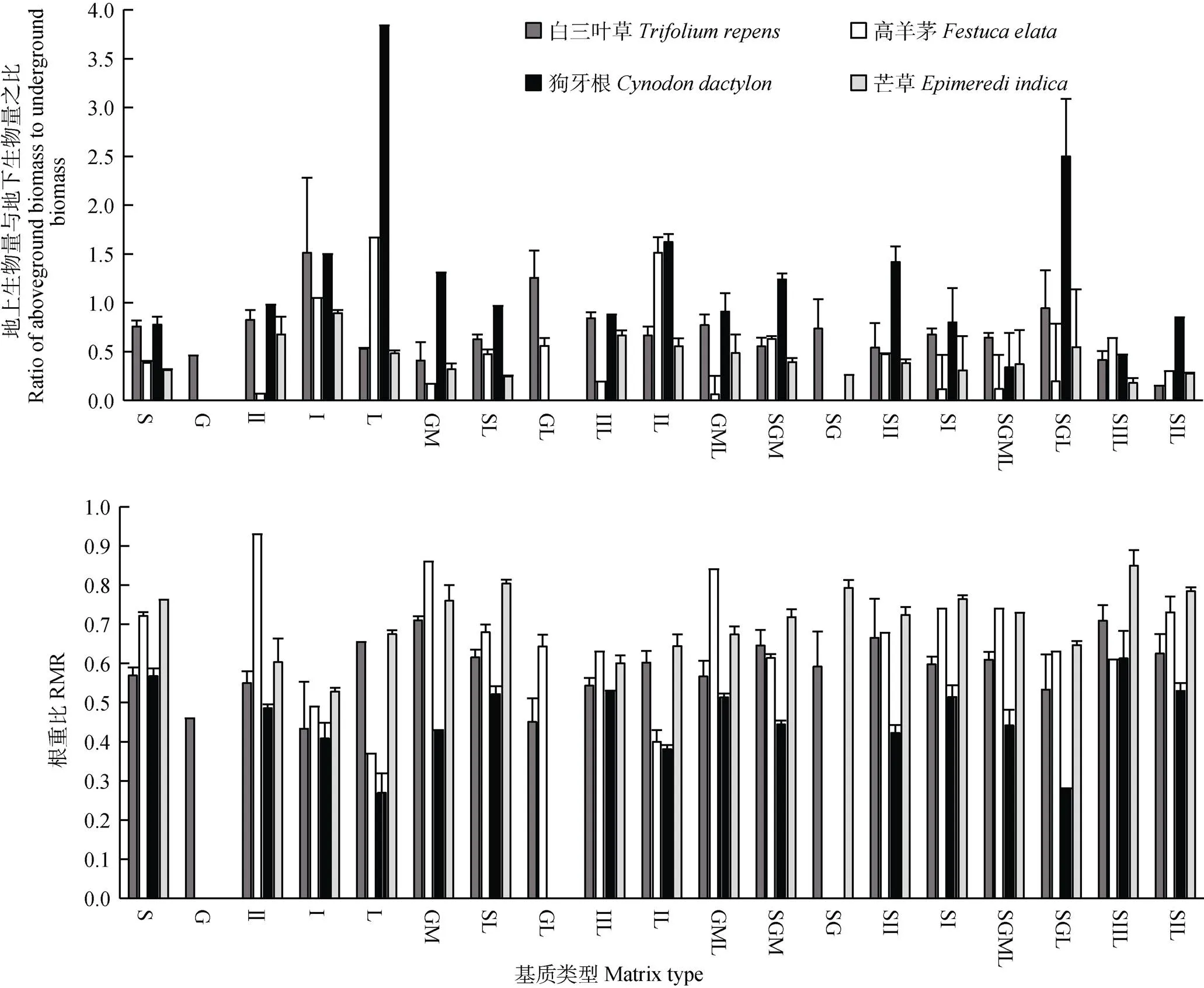
圖3 四種草本植物在不同基質中的生物量之比
基質作為植物生長的必要載體,它不僅是植物種子及幼苗萌發和生存的場所,也是為植物生長發育提供所需水分、養分的介質,因此基質類型對植物的生長發育有著重要作用。基質的物理和化學特性直接影響基質的蓄水能力、透水性及通氣性,間接影響著基質的肥力和植物生長狀況[28]。通過對基質理化特性的分析,得出種植植物后,整體上基質總孔隙度、AP、WHC、水氣比、pH、TN、TP增高,TK含量降低,基質呈堿性或強堿性,說明基質的透氣性增強,空氣容量增大,保水能力增強,在降雨或者澆水后能蓄存更多的水分,延長干旱季節水分的蒸發時間,從而為植物根系吸收水分創造有利條件,反映了植物根系對生長基質具有調節作用;氮和磷是限制植物生長的重要元素,對植物生長發育具有重要作用,而氮和磷在土壤中極易被固定,且有效性低,大氣中的氮元素被基質吸收,導致其中的TN含量增加,而19種基質中土壤占比低,砂石占比高,淋溶作用會產生較多的磷元素進入基質中,使基質中磷含量增加[29-30],因而大量的氮和磷在基質中積累,導致氮和磷含量增加,其中白三葉草根系具有根瘤菌,具有明顯的固氮作用,因此其基質中TN含量明顯增加;同時,基質中養分因子也受到植被及其他環境因子的影響,如土壤水分、降雨、土壤質地等,也會導致基質中出現TN、TP含量增加現象[29];本研究與王霖嬌等[29]的研究結果一致,但與Mulder和Elser[31]、Zhang等[32]的研究結果不一致,上述分析顯示喀斯特生境中植物生長基質特性變化的多樣性與特殊性;TK含量降低說明植物在生長過程中吸收鉀以促進其生長發育。綜上可知,植物在生長過程中會對其生長基質進行調節,以創造更有利于其生長的外界環境。因此在喀斯特石漠化區為了維持或提高植物生長基質的肥力或地力,應該按照科學原理,在合適的基質上種植適應的植物,從而形成良性循環的基質生態系統,確保植被生態系統得以恢復與重建。

圖4 四種植物在不同基質中的葉綠素含量(Chl a+b)、類胡蘿卜素與葉綠素含量的比值(Car/Chl(a+b))
植物的發芽率和存活率能夠直接反映植物的生長狀況。整體上,白三葉草的發芽率最高,狗牙根存活率最高,四種植物在石灰土、Ⅰ砂、土Ⅰ砂中發芽率和存活率高,在碎石枯中低,但在碎石中存活率均為0,說明植物在碎石中難以存活,而植物卻能在非土壤基質的其他生長基質中有較高的生存率和發芽率,反映了在喀斯特極端生境中植物仍然具有較高的適應性;分析原因是石灰土、Ⅰ砂、土Ⅰ砂的總孔隙度和持水量大,基質疏松、透氣性和保水性能好,其他與碎石、Ⅱ砂等混合的基質,因碎石與Ⅱ砂的粒徑較大,從而基質的孔隙度與持水量增大[33],使基質保持一個好的透氣性與保水能力,種子也能在其中萌發與存活,而碎石和碎石枯的粒徑過大、氣體孔隙度高,保水性差,易造成水分缺失,植物生長沒有足夠的水分,且草本植物根系較淺,很難吸收更深層次的水分和養分,因此植物在碎石和碎石枯中難以存活。生物量是植物對不同基質生長適應的綜合體現,也是對環境整體適應的結果[34-35],同時生物量的分配格局在一定程度上可以反映植物受到水分脅迫的生存策略,植物受到干旱脅迫時通常將較多的生物量分配到地下部分以促進根系生長獲取必要的水分來維持其生命活動的正常進行[36-37],因此生物量累積值可以為植物對不同基質生長適應提供可靠的標準。本試驗中狗牙根的地上生物量與地下生物量之比最大,其次為白三葉草、高羊茅,芒草最小(四種植物的RMR剛好相反),這反映了四種植物的生存策略不同,狗牙根和白三葉草地上部分生長更為發達,采取分配更多的水分及養分到地上部分,以便獲得更好的地上資源,而芒草和高羊茅根系更為發達,地上光合產物向根部運輸的比例高于地上部分,使根系能吸收更多的營養和水分來維持植物生存,體現植物對逆境的適應性。葉綠素是植物進行光合作用的重要物質,在植物對光能的吸收和轉換中起著重要作用。本研究中,四種植物的Chl a+b和Car/Chl(a+b)在土枯、土Ⅰ砂、土Ⅰ砂枯中高,碎石中最低,且白三葉草的Chl(a+b)最高,其次為狗牙根、芒草,高羊茅最低,說明四種植物在土枯、土Ⅰ砂、土Ⅰ砂枯中能夠吸收更多的光能,有利于光合作用,在碎石和碎石枯中則降低葉綠素含量以增強光保護能力,使其能夠在逆境中生長;植物的Car/Chl(a+b)比值反映植物的光保護能力[32-33],狗牙根的Car/Chl(a+b)最大,其次芒草、三葉草,高羊茅最小,說明狗牙根和芒草更有利于增強光保護能力,反映狗牙根和芒草更能適應惡劣環境,揭示植物光合色素的分配反映植物對于不同基質采用的光合機制不同。δ13C值既由植物本身的生物學特性決定,又受到外界環境因素的影響[38]。四種草本植物的δ13C值符合韓家懋等[39]的研究結果,芒草的δ13C值最大,其次為狗牙根,白三葉草和高羊茅的較小,說明C4植物水分利用效率遠遠高于C3植物,更適合在缺乏水分的逆境中生長;同一植物在不同生長基質中δ13C值也有所不同,白三葉草在Ⅰ砂、土砂石混、土砂石混枯和碎枯中大,狗牙根在Ⅰ砂中最大,說明植物葉片δ13C值跟植物種類與生長基質有關。
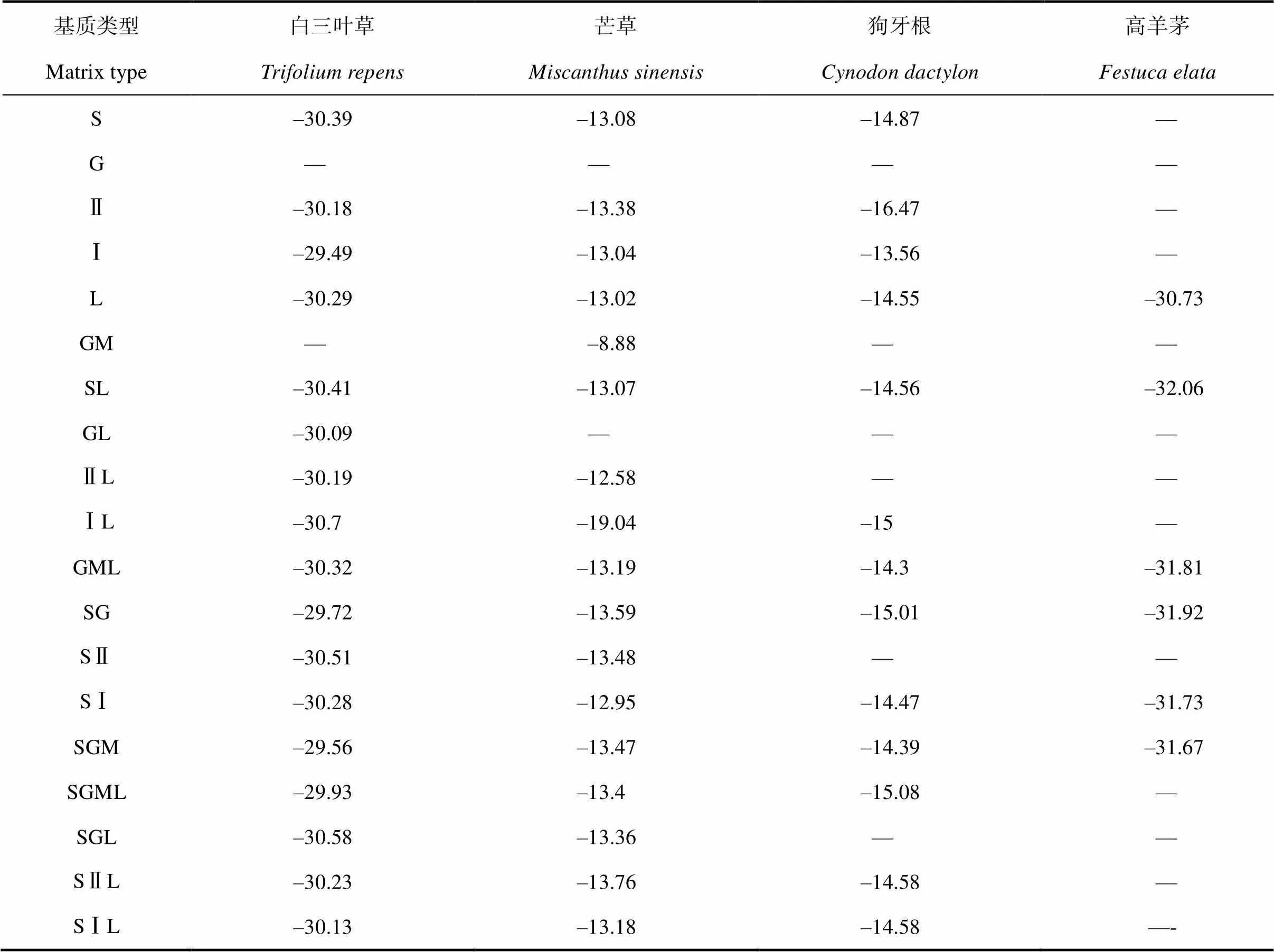
表2 四種植物葉片的δ13C值
注:“-”表示在相對應的基質中已經落葉而未測定的植物葉片δ13C值。Note:“-” refers to the undetermined δ13C values of plant leaves that have been defoliated in the corresponding matrix.
綜上所述,整體上,在不同基質中,四種植物通過調節自身的形態、生理特征,如株高、蓋度、Chl a+b含量、生物量分配及水分利用效率等來適應不同的生長基質,對于碎石或者砂石占比較高的基質采取降低Chl a+b含量,增強水分利用效率,擴大根系,增強光保護能力,吸收更多的水分和養分,以維持自身生長。植物通過調節基質理化特性創造更有利于其生長的基質環境。四種植物能在除土壤外的其他基質中生長良好,但在碎石和碎石枯中難以存活。整體上,四種植物對石灰土、Ⅰ砂、土枯、土Ⅰ砂和土Ⅱ砂的適應性強,對Ⅱ砂、枯落物、砂石混枯、土Ⅱ砂枯、土Ⅰ砂枯的較強,對砂石混、Ⅱ砂枯、土Ⅰ砂、土砂石混、土砂石混枯的較差,對碎石、碎石枯的最差。其中狗牙根和芒草對于19種基質的適應性強,白三葉草較強,高羊茅適應性差,因此,在喀斯特石漠化恢復初期,可以優先選擇種植狗牙根和芒草。
4 結 論
植物種植前后基質理化性質發生改變,基質呈堿性或強堿性;不同植物對于生長基質的適應性調節方式不同,通過調整生物量的分配格局來適應不同的生存環境,同時調節光合色素以適應不同基質的環境機制,C4植物狗牙根和芒草的水分利用效率高,更能適應惡劣的生長基質,植物葉片δ13C值不僅與植物葉片有關,還與生長基質有關,建議在喀斯特區植被恢復初期多選用C4草本植物;四種植物在石灰土、Ⅰ砂、土枯、Ⅰ砂枯和土Ⅱ砂適應性強,其次為砂石混、Ⅱ砂枯、土Ⅰ砂、土砂石混、土砂石混枯等,在碎石、碎石枯、土碎石、土碎石枯中差;整體上,狗牙根和芒草生長適應性強,白三葉草較強,高羊茅最差,因此在喀斯特石漠化區恢復進程中,可以選擇優先種植芒草和狗牙根;本研究基于單一土壤基質研究拓展到喀斯特生長基質研究,拓寬了研究理論和研究視角,對喀斯特區植被生態恢復具有重要意義。綜上所述,研究喀斯特區四種草本植物及其基質優先選擇,對喀斯特石漠化區生態恢復和重建有重要意義,同時也能為喀斯特石漠化區草本植被恢復提供物種選擇。
[1] Liu C Q. Biogeochemical processes and cycling of nutrients in the earth’s surface:Cycling of nutrients in soil-plant systems of karstic environments,Southwest China[M]. Beijing:Science Press,2009.
[2] Zhu S Q. Study on Karst forest ecology[M]. Guiyang:Guizhou Science and Technology Rress,1993. [朱守謙. 喀斯特森林生態研究[M]. 貴陽:貴州科學技術出版社,1993. ]
[3] Li J Q. Forest ecology[M]. Beijing:High Education Press,2010. [李俊清. 森林生態學[M]. 北京:高等教育出版社,2010. ]
[4] Ou Z Y,Cao Y Y,Tan Z Q,et al. Effects of drought on photosynthesis and resistance physiology ofseedlings in Karst habitat[J]. Chinese Journal of Ecology,2018,37(11):3270—3276. [歐芷陽,曹艷云,譚長強,等. 干旱脅迫對喀斯特生境蜆木幼苗光合特性及抗性生理的影響[J]. 生態學雜志,2018,37(11):3270—3276. ]
[5] Chen C. Ecophysiological mechanisms of forage shrubs resistant to drought&cold in Karst areas[D]. Beijing:China Agricultural University,2014. [陳超. 喀斯特地區飼用灌木抗旱抗寒性的生理生態學機制研究[D]. 北京:中國農業大學,2014. ]
[6] Zhang Z F,You Y M,Huang Y Q,et al. Effects of drought stress on the photosynthesis and growth of Cyclobalanopsis glauca seedlings:A study with simulated hierarchical Karst water supply[J]. Chinese Journal of Ecology,2012,31(9):2197—2202. [張中峰,尤業明,黃玉清,等. 模擬巖溶水分供應分層的干旱脅迫對青岡櫟光合特性和生長的影響[J]. 生態學雜志,2012,31(9):2197—2202. ]
[7] Ji F T,Li N,Deng X. Calcium contents and high calcium adaptation of plants in Karst areas of China[J]. Chinese Journal of Plant Ecology,2009,33(5):926—935. [姬飛騰,李楠,鄧馨. 喀斯特地區植物鈣含量特征與高鈣適應方式分析[J]. 植物生態學報,2009,33(5):926—935. ]
[8] Liu C C,Liu Y G,Guo K. Ecophysiological adaptations to drought stress of seedlings of four plant species with different growth forms in Karst habitats[J]. Chinese Journal of Plant Ecology,2011,35(10):1070—1082. [劉長成,劉玉國,郭柯. 四種不同生活型植物幼苗對喀斯特生境干旱的生理生態適應性[J]. 植物生態學報,2011,35(10):1070—1082. ]
[9] Huang C Y,Xu J M. Soil science[M]. Beijing:China Agriculture Press,2010. [黃昌勇,徐建明. 土壤學[M]. 北京:中國農業出版社,2010. ]
[10] Wang G F. Landscape floristics[M]. Hangzhou:Zhejiang University Press,2018. [王國夫. 園林花卉學[M]. 杭州:浙江大學出版社,2018. ]
[11] Wan C G. Floriculture[M]. Shanghai:Shanghai Jiaotong University Press,2008. [宛成剛. 花卉學[M]. 上海:上海交通大學出版社,2008. ]
[12] Pan B H. Landscape floristics[M]. Changsha:National Defense University of science and Technology Press,2007. [潘百紅. 園林花卉學[M]. 長沙:國防科技大學出版社,2007. ]
[13] Fu Y H,Huang Z S,Yu L F,et al. Analysis on the soil quality of different typical underground root habitat types in Karst areas[J]. Research of Soil and Water Conservation,2012,19(3):67—73. [符裕紅,黃宗勝,喻理飛,等. 巖溶區典型根系地下生境的土壤質量分析[J]. 水土保持研究,2012,19(3):67—73. ]
[14] Huang Z S,Fu Y H,Yu L F. Characteristic evolution of soil organic carbon pool with the process of natural restoration of Karst forest vegetation[J]. Acta Pedologica Sinica,2013,50(2):306—314. [黃宗勝,符裕紅,喻理飛. 喀斯特森林植被自然恢復過程中土壤有機碳庫特征演化[J]. 土壤學報,2013,50(2):306—314. ]
[15] Liu L,Zeng F P,Song T Q,et al. Spatial heterogeneity of soil nutrients in Karst area’s Mulun National Nature Reserve[J]. Chinese Journal of Applied Ecology,2010,21(7):1667—1673. [劉璐,曾馥平,宋同清,等. 喀斯特木論自然保護區土壤養分的空間變異特征[J]. 應用生態學報,2010,21(7):1667—1673. ]
[16] Li A D,Li W J,Yang R,et al. Capacity of soil moisture conservation of different microhabitats under different mulching techniques in Karst area[J]. Science of Soil and Water Conservation,2010,8(5):55—60,66. [李安定,李葦潔,楊瑞,等. 喀斯特地區不同覆蓋下小生境土壤保墑能力[J]. 中國水土保持科學,2010,8(5):55—60,66. ]
[17] Liang S C,Zhang S M,Yu F H,et al. Small-scale spatial cross-correlation between ramet population variables ofvar.and soil available phosphorus[J]. Journal of Plant Ecology,2007,31(4):613—618. [梁士楚,張淑敏,于飛海,等. 絹毛匍匐委陵菜與土壤有效磷的小尺度空間相關分析[J]. 植物生態學報,2007,31(4):613—618. ]
[18] Zhang W,Wang K L,Chen H S,et al. Use of satellite information and GIS to predict distribution of soil organic carbon in depressions amid clusters of Karst peaks[J]. Acta Pedologica Sinica,2012,49(3):601—606. [張偉,王克林,陳洪松,等. 典型喀斯特峰叢洼地土壤有機碳含量空間預測研究[J]. 土壤學報,201,49(3):601—606. ]
[19] Yu L F,Zhu S Q,Ye J Z,et al. Dynamics of a degraded Karst forest in the process of natural restoration[J]. Scientia Silvae Sinicae,2002,38(1):1—7. [喻理飛,朱守謙,葉鏡中,等. 退化喀斯特森林自然恢復過程中群落動態研究[J]. 林業科學,2002,38(1):1—7. ]
[20] Standard production group. Standard for quality and inspection method of sand and stone for ordinary concrete (JGJ 52-2006) [S]. Beijing:China Building Materials Industry Press,2008. 5. isbn978-7-80227- 398-6. [標準制作組. 普通混凝土用砂石質量及檢驗方法標準(JGJ 52-2006)[S]. 北京:中國建材工業出版社,2008. 5. ISBN978-7-80227-398-6. ]
[21] Liu G S. Soil physical and chemical analysis and profile description[M]. Beijing:China Standard Press,1996. [劉光崧. 土壤理化分析與剖面描述[M]. 北京:中國標準出版社,1996. ]
[22] Qiao S Y. Instructions for soil physical and chemical properties experiments[M]. Beijing:China University of Geosciences Press,2012:47—50. [喬勝英. 土壤理化性質實驗指導書[M]. 北京:中國地質大學出版社,2012:47—50. ]
[23] Bao S D. Soil agrochemical analysis[M]. Beijing:China Agriculture Press. 2002. [鮑士旦. 土壤農化分析[M]. 北京:中國農業出版社. 2002. ]
[24] Styer R C,Koranski D S. Principle and technology of point-plate seedling production[M]. Liu B,et al. trans. Beijing:Chemical Industry Press,2007:80—83. [Roger C. Styer,David S. Koranski. 穴盤苗生產原理與技術[M]. 劉濱,等譯. 北京:化工工業出版社,2007:80—83. ]
[25] Lichtenthaler H K. Chlorophylls and carotenoids:Pigments of photosynthetic biomembranes[M]//Methods in Enzymology. Amsterdam:Elsevier,1987:350—382
[26] Rong L,Wang S J,Du X L,et al. Relationship among leaf anatomical characters and foliar δ13C values of six woody species for Karst rocky desertification areas[J]. Scientia Silvae Sinicae,2008,44(10):29—34. [容麗,王世杰,杜雪蓮,等. 喀斯特峽谷石漠化區6種常見植物葉片解剖結構與δ13C值的相關性[J]. 林業科學,2008,44(10):29—34. ]
[27] Meng X M. The concept of growing media and its application in the protected agriculture in China[J]. Journal of Shanxi Agricultural University(Natural Science Edition),2016,36 (3):155—159,165. [孟憲民. 專業基質的概念及其在我國設施農業中的意義[J]. 山西農業大學學報(自然科學版),2016,36(3):155—159,165. ]
[28] Liu J. Study on characteristics of vegetation and soil under different ecological rehabilitation measures in Karst area[D]. Guiyang:Guizhou University,2017. [劉娟. 喀斯特不同生態修復措施下植被與土壤特征研究[D]. 貴陽:貴州大學,2017. ]
[29] Wang L J,Wang P,Sheng M Y. Stoichiometric characteristics of soil nutrient elements and its influencing factors in typical karst rocky desertification ecosystems,Southwest China[J]. Acta Ecologica Sinica,2018,38(18):6580—6593. [王霖嬌,汪攀,盛茂銀. 西南喀斯特典型石漠化生態系統土壤養分生態化學計量特征及其影響因素[J]. 生態學報,2018,38(18):6580—6593. ]
[30] Qin C X,Peng C,Guo Q,et al. Effects of sugarcane and peanut intercropping on available phosphorus and pH value in red soils[J]. Jiangsu Agricultural Sciences,2019,47(11):137—140. [秦昌鮮,彭崇,郭強,等. 甘蔗花生間作對紅壤有效磷、pH值的影響[J]. 江蘇農業科學,2019,47(11):137—140. ]
[31] Mulder C,Elser J J. Soil acidity,ecological stoichiometry and allometric scaling in grassland food webs[J]. Global Change Biology,2009,15(11):2730—2738.
[32] Zhang C,Tian H Q,Liu J Y,et al. Pools and distributions of soil phosphorus in China[J]. Global Biogeochemical Cycles,2005,19(1):doi:10. 1029/2004GB002296.
[33] Zhao M,Wang W L,Guo M M,et al. Erosion of slopes of lou soil stacks different in limestone debris content[J]. Acta Pedologica Sinica,2020,57. DOI:10. 11766/ trxb201905220129. [趙滿,王文龍,郭明明,等. 不同礫石含量塿土堆積體坡面侵蝕特征研究[J]. 土壤學報,2020,57. DOI:10. 11766/trxb201905220129. ]
[34] Zhang X X,Yin X L,Li H L,et al. Effect of salt stress on the biomass and photosynthetic characteristics ofL. strains[J]. Acta Ecologica Sinica,2017。37(21):7258—7265. [張曉曉,殷小琳,李紅麗,等. NaCl脅迫對不同白榆品系生物量及光合作用的影響[J]. 生態學報,2017,37(21):7258—7265. ]
[35] Zong N,Duan C,Geng S B,et al. Effects of warming and nitrogen addition on community production and biomass allocation in an alpine meadow[J]. Chinese Journal of Applied Ecology,2018,29(1):59—67. [宗寧,段呈,耿守保,等. 增溫施氮對高寒草甸生產力及生物量分配的影響[J]. 應用生態學報,2018,29(1):59—67. ]
[36] Baquedano F J,Castillo F J. Comparative ecophysiological effects of drought on seedlings of the Medit erranean water-saverand water-spendersand[J]. Trees,2006,20(6):689—700.
[37] Elsheery N I,Cao K F. Gas exchange,chlorophyll fluorescence,and osmotic adjustment in two mango cultivars under drought stress[J]. Acta Physiologiae Plantarum,2008,30(6):769—777.
[38] Dong X,Li Y H,Xin Z M,et al. Variation in leaf traits and leaf δ13C and δ15N content in Nitraria tangutorum along precipitation gradient[J]. Acta Ecologica Sinica,2019,39(10):3700—3709. [董雪,李永華,辛智鳴,等. 唐古特白刺葉性狀及葉片δ13C、δ15N沿降水梯度的變化特征[J]. 生態學報,2019,39(10):3700—3709. ]
[39] Han J M,Wang G A,Liu D S. Appearance of C4plants and global changes[J]. Earth Science Frontiers,2002,9(1):233—243. [韓家懋,王國安,劉東生. C4植物的出現與全球環境變化[J]. 地學前緣,2002,9(1):233—243. ]
Four Species of Herbs in Karst Areas and Their Adaptabilities to the Karst Matrices
CHEN Haixia1, HUANG Zongsheng2?, ZHAO Mingwei2, LI Zuguo1,3, LIU Yifu1, ZHANG Lixin2, YANG Chun1, YANG Zhengrong1, ZHANG Yuanbo2, 4
( 1. Foresty College of Guizhou University, Guiyang 550025, China; 2. City Plan and Architecture College of Guizhou University, Guiyang 550025, China; 3. Zunyi Garden Management Station, Zunyi, Guizhou 563000, China; 4. College of Humanities and Physical Education, Guizhou Technology and Business Institute, Guiyang 551400, China)
【】 This paper is aimed to find out priority of plants in adaptability to karst matrices.【】Four native species of plants,,,andin the karst areas were selected for test in a pot experiment with pots packed with different matrices, separately 【】Results show that the karst habitat diversifies in plant growing matrix. Physico-chemical properties of the matrices as a whole improved with plants growing on and on, except for decrease in total potassium content. The plants were the highest in germination rate and survival rate in the pots of limestone soil, < 1 mm sand and < 1 mm sand + limestone soil and the lowest in the pots of limestone debris and limestone debris + litter, while among the four species of plants,andwas the highest in germination rate and survival rate, respectively. With the plants growing on and on, the plants were higher in plant number, plant height and coverage in the pots of limestone soil, < 1 mm sand, < 1 mm sand + litter and lower in the pots of limestone debris and limestone debris + litter; and the plants in all the pots increased in coverage, and onlyincreased in number. The plants in the pots of limestone soil, < 1 mm sand, litter, and < 1 mm sand + litter were the highest, and the lowest in the pots of limestone debris, and limestone debris + litter in biomass, while among the plants,andwere high andandwere low. The plants were high in total chlorophyll in the pots of < 1 mm sand, limestone soil, limestone soil + 1 mm–5 mm sand and < 1 mm sand+ 1 mm–5 mm sand + limestone debris + limestone soil, and low in the pots of limestone debris, while among the plants,andwere higher andandlower, andandwere higher in δ13C thanand. The value of δ13C in the plant is related not only to species of the plant, but also to the matrix it grew in. 【】To sum up, the plants in karst areas can grow not only in limestone soil, but also in other matrices. The four species of plants in this experiment are highly adaptable to the matrices of limestone soil, < 1 mm sand, < 1 mm sand + litter and limestone soil + 1 mm–5 mm sand, fairly adaptable to 1 mm–5 mm sand, litter, <1 mm sand + 1 mm–5 mm sand + limestone debris + litter, limestone soil + 1 mm–5 mm sand + litter, <1 mm sand + limestone soil + litter and 1 mm–5 mm sand + litter, and barely adaptable to limestone debris and limestone debris + litter, andandare more adaptable than, and. Therefore,andcan be selected preferentially as pioneer plant for ecological restoration in karst areas. The extension from studies on the single soil matrix to studies on plant growing matrices of karst areas helps broadens the research theory and perspective, which is of great significance to ecological vegetation restoration in karst areas.
Karst; Herb plants; Matrix; Adaptability; Ecological restoration
Q14
A
10.11766/trxb201907170370
陳海霞,黃宗勝,趙明偉,李祖國,劉逸夫,張麗欣,楊春,楊政蓉,張元博. 喀斯特地區四種草本植物及其基質的優先選擇[J]. 土壤學報,2020,57(4):1017–1031.
CHEN Haixia,HUANG Zongsheng,ZHAO Mingwei,LI Zuguo,LIU Yifu,ZHANG Lixin,YANG Chun,YANG Zhengrong,ZHANG Yuanbo. Four Species of Herbs in Karst Areas and Their Adaptabilities to the Karst Matrices[J]. Acta Pedologica Sinica,2020,57(4):1017–1031.
* 國家自然科學基金項目(31560187,51868008,51987187)資助Supported by the National Natural Science Foundation of China(Nos.31560187,51868008,51987187)
,E-mail:hzsxjh@126.com
陳海霞(1993—),女,碩士研究生,主要從事退化生態系統恢復,景觀規劃設計研究。E-mail:1434473499@qq.com
2019–07–17;
2019–11–27;
2020–03–11
(責任編輯:檀滿枝)
猜你喜歡
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
