羊口瘡病毒ORFV129錨蛋白在山羊睪丸細胞中的亞細胞定位
2020-08-29 13:29:46朱江江張煥容
華北農(nóng)學報 2020年4期
向 華,黃 忍,朱江江,岳 華,湯 承,張煥容
(1.西南民族大學 生命科學與技術學院,四川 成都 610041;2.青藏高原動物遺傳資源保護與利用,四川省重點實驗室,四川 成都 610041)
羊傳染性膿皰皮炎俗稱“羊口瘡”,是由羊口瘡病毒(Orfvirus,ORFV)引起的一種急性、接觸性、嗜上皮性的人獸共患傳染病[1]。該病毒主要感染綿羊及山羊,特別是羔羊,人類接觸也可感染[2]。臨床上主要以感染動物的唇、鼻孔、口腔黏膜、乳房、陰部等部位皮膚形成紅斑、水皰、膿皰、丘疹和疣狀痂皮為特征[3-5]。初生羔羊和幼齡羊感染ORFV后發(fā)病率高達100%,死亡率達93%;雖然成年羊感染ORFV后的死亡率較低,但由此引起的采食困難,會導致機體消瘦、生長緩慢,以及繼發(fā)感染,帶來巨大的經(jīng)濟損失。近年來,隨著我國養(yǎng)羊數(shù)量增加和跨境流通,ORF作為人畜共患的傳染病,對人類的健康也產(chǎn)生了威脅,受到社會的廣泛關注。然而,由于ORFV特有的免疫機制和免疫逃逸機制,目前的疫苗并不能為羊提供完全的免疫保護。同時,現(xiàn)有的針對羊口瘡的治療藥物大多為“對癥治療”的緩解性藥物,其治療效果不甚理想,且費時費力。羊口瘡難以根治的特點和反復發(fā)作的現(xiàn)狀已成為制約養(yǎng)羊產(chǎn)業(yè)發(fā)展的瓶頸。
ORFV屬于痘病毒科副痘病毒屬,為雙股DNA病毒,共包含130多個基因[6]。由末端基因區(qū)(ORFVs001-008和ORFVs112-134)和中間核心基因區(qū)(ORFVs009-111)組成,中間核心區(qū)包含眾多保守基因,多與病毒的復制和形態(tài)相關,末端基因區(qū)多為非保守基因,與病毒毒力和致病性相關[7-9]。其中末端基因中包括5種編碼錨蛋白的基因,分別為ORFV008、ORFV123、ORFV126、ORFV128和ORFV129。ORFV129蛋白與其他ORFV錨蛋白具有相似的結構,包含多個錨蛋白重復基序(ANKs)和1個F-Box樣結構域[10-11]。ORFV129蛋白的功能目前不清楚,而蛋白質生物學功能的發(fā)揮往往與該蛋白的亞細胞定位密切相關[12]。本研究旨在克隆羊口瘡病毒ORFV129基因全長,并對其進行生物信息學分析。進一步構建真核表達載體pEGFP-ORFV129,轉染新生山羊睪丸原代細胞研究該蛋白在細胞中的定位特征,為進一步研究ORFV129在誘導機體免疫應答中的作用奠定基礎。
1 材料和方法
1.1 毒株、菌株、真核質粒和細胞
羊口瘡病毒JYY-ORFV株、新生山羊睪丸原代細胞和pEGFP-N1載體及大腸桿菌DH5α購自寶生物工程(大連)有限公司(TaKaRa)。
1.2 主要試劑及儀器
PremixTaq、DL2000 DNA Marker、T4DNA連接酶均購自寶生物工程(大連)有限公司(TaKaRa);Hind Ⅲ、PstⅠ購自上海玉博生物科技有限公司;膠回收試劑盒、質粒提取試劑盒均購自美國Omega公司;DMEM培養(yǎng)基、胎牛血清和OPTI-MEM 1(1X)均購自美國Gibco公司;RAPI裂解液、廣譜蛋白酶抑制劑、廣譜磷酸酶抑制劑、組織固定液、一抗鼠源GFP、一抗兔源GADPH均購自武漢博士德生物工程有限公司;HRP標記的山羊抗鼠IgG購自武漢亞科因生物技術有限公司;HRP標記的山羊抗兔IgG購自北京博奧龍免疫技術有限公司;DAPI核酸染料、LipoGeneTM2000均購自US Everbright?Inc.(簡稱UE)公司;Triton X-100、DNase Ⅰ均購自廣州賽國生物科技有限公司;熒光倒置顯微鏡購自日本奧林巴斯(Olympus Corporation);激光共聚焦顯微鏡購自德國卡爾·蔡司公司股份公司(Carl Zeiss AG)。
1.3 ORFV129基因全長的克隆鑒定及生物信息學分析
參照GenBank中登錄的羊口瘡病毒SY 17株ORFV129(登錄號:MG 712417.1),運用Primier Primer 5設計1對特異性引物。上游引物:5′-AACGCCACGCCATGGACGC-3′;下游引物:5′-GATGGGCAGTCACTCGGGCG-3′,引物由上海生工生物工程有限公司合成。選取接種了ORFV且具有明顯細胞病變的新生山羊睪丸原代細胞培養(yǎng)物反復凍融3次,12 000 r/min離心2 min,留上清,使用DNA提取試劑盒提取病毒DNA。以提取的病毒DNA為模板,使用上下游引物進行PCR反應,特異性擴增全長ORFV129基因。反應體系:Pre mix Taq 10 μL,上、下游引物各1 μL,模板1 μL,其余用ddH2O補足20 μL。反應條件:94 ℃預變性4 min;94 ℃變性30 s,65 ℃退火30 s,72 ℃延伸1 min,共35個循環(huán);72 ℃再延伸7 min。PCR產(chǎn)物經(jīng)1.5%瓊脂糖凝膠電泳進行檢測,切膠回收產(chǎn)物連接pMD19-T載體,并轉化DH5α感受態(tài)細胞,提取質粒后陽性的重組質粒送上海生工生物工程有限公司測序后保存?zhèn)溆茫瑢㈣b定正確的質粒命名為pMD19-ORFV129。對測序鑒定正確的ORFV129全基因序列進行相關結構和功能的預測:使用SMART(http://smart.emblheidelberg.de/smart/set_mode.cgi?NORMAL=1)軟件進行一級結構分析;使用TMHMM Server v. 2.0(http://www.cbs.dtu.dk/services/TMHMM-2.0/)軟件進行跨膜區(qū)域分析;使用在線軟件SignalP 5.0(http://www.cbs.dtu.dk/services/SignalP/)進行信號肽分析以及在線軟件WOLF(https://wolfpsort.hgc.jp/)進行蛋白亞細胞定位的預測。
1.4 融合表達ORFV129基因真核重組表達載體的構建及鑒定
參照ORFV SY 17株ORFV129(登錄號:MG 712417.1),運用Primier Primer 5設計引物。上游引物5′-AAGCTTATGGACGCCGCCGAGATGGAGGAGCTCGACATC-3′(劃線部分為Hind Ⅲ酶切位點);下游引物5′-CTGCAGCTCGGGCGCGCGGCGGTCGCACGAGCGGCGC-3′(劃線部分為PstⅠ酶切位點),引物由上海生工生物工程有限公司合成。以1.3中質粒pMD19-ORFV129為模板,PCR擴增帶酶切位點的129基因,反應體系及條件同1.3,1.5%瓊脂糖凝膠電泳進行檢測,切膠回收產(chǎn)物。將膠回收產(chǎn)物和載體pEGFP-N1分別使用Hind Ⅲ、PstⅠ進行雙酶切,酶切產(chǎn)物經(jīng)PCR產(chǎn)物純化試劑盒純化后,T4DNA連接酶16 ℃金屬儀連接過夜,轉化DH 5α感受態(tài)細胞,提取質粒后經(jīng)PCR、雙酶切鑒定為陽性的重組質粒送上海生工生物工程有限公司測序后保存?zhèn)溆茫瑢㈣b定正確的質粒命名為pEGFP-ORFV129。
1.5 pEGFP-ORFV129重組質粒轉染新生山羊睪丸原代細胞
將生長良好的新生山羊睪丸原代細胞接種于6孔細胞培養(yǎng)板,待細胞匯合度達70%~90%時,按照LipoGeneTM2000 轉染試劑說明書,通過脂質體介導法將重組表達質粒pEGFP-ORFV129轉染至生長良好的新生山羊睪丸原代細胞,以空載體pEGFP-N1為轉染對照,并以正常生長的細胞為非轉染空白對照。
1.6 檢測pEGFP-ORFV129重組質粒在睪丸細胞的mRNA表達
在轉染12~24 h后,將轉染空載pEGFP-N1和融合載體pEGFP-ORFV129的細胞培養(yǎng)板置于熒光倒置顯微鏡下,觀察pEGFP-ORFV129融合蛋白在睪丸細胞中的瞬時表達情況。對于檢測轉錄動力學,在轉染48 h后,用TRIzol法提取細胞總RNA,并用DNase Ⅰ消化殘留于總RNA中的DNA,總RNA逆轉錄為cDNA,以cDNA為模板進行PCR擴增,擴增引物和條件同1.4,1.5%瓊脂糖凝膠電泳進行檢測。通過逆轉錄PCR(RT-PCR)檢測ORFV129基因在細胞中ORFV復制期間的mRNA表達。
1.7 檢測ORFV129蛋白在羊睪丸細胞中的蛋白表達
將生長良好的睪丸細胞接種于6孔細胞培養(yǎng)板上,設3組,每組2個重復,分別為空白對照組、pEGFP-N1轉染組(4.0 μg)、pEGFP-ORFV129轉染組(4.0 μg)。轉染36 h后,將貼壁細胞刮取到1.5 mL的離心管,將廣譜蛋白酶抑制劑:RAPI細胞裂解液按照1∶100的比例混勻的細胞裂解液加入裝有細胞的離心管,在超聲波破碎儀作用下破碎,離心,收集蛋白。隨后,將提取的細胞總蛋白經(jīng)SDS-PAGE電泳后,轉入聚偏二氟乙烯(PVDF)膜上,經(jīng)5%(m/V)脫脂奶中,4 ℃封閉過夜。隨后分別加入鼠源GFP(1∶400)和兔源GADPH(1∶1 000),室溫孵育2 h,10 min/次TBST(3次)的洗膜。利用HRP標記山羊抗鼠IgG(1∶400)和HRP標記的山羊抗兔IgG(1∶400),室溫孵育2 h。TBST洗膜3次,隨后使用ECL試劑顯色,成像。
1.8 融合蛋白ORFV129的亞細胞定位
將睪丸細胞培養(yǎng)于含有蓋玻片的6孔細胞培養(yǎng)板,待細胞匯合度達70%~90%時,轉染重組表達質粒pEGFP-ORFV129,以空載體pEGFP-N1為對照,重組表達質粒組和空載體組每組設2個平行。轉染48 h后吸盡培養(yǎng)液,預冷PBS洗3次,4%的多聚甲醛室溫孵育15 min,預冷PBS洗3次;0.1%的Triton X-100室溫孵育10 min,預冷PBS洗3次;加入10~50 μmol/L DAPI核酸染料,室溫孵育15 min,預冷PBS洗2次;取出蓋玻片,將其貼于含有封片劑的載玻片上,于激光共聚焦顯微鏡下觀察ORFV129在細胞內的分布情況,分析其亞細胞定位。
2 結果與分析
2.1 ORFV129基因的PCR擴增、克隆鑒定及生物信息學分析
成功擴增ORFV129基因片段,經(jīng)瓊脂糖凝膠電泳檢測可見與預期大小(1 551 bp)相符的目的片段(圖1-A)。膠回收克隆轉化后,進行菌落PCR,經(jīng)瓊脂糖凝膠電泳檢測,與預期ORFV129基因的大小相符(圖1-B)。測序結果顯示,與NCBI上收錄的同一病毒的不同毒株基因序列相似性均在99%以上。利用DNAStar軟件分析發(fā)現(xiàn),ORFV129基因共編碼516個氨基酸。SMART分析顯示,ORFV129基因共含有8個ANK基序,分別位于49-80 aa,85-117 aa,123-157 aa,161-192 aa,196-227 aa,231-262 aa,266-297 aa,302-331 aa(圖2-A)。跨膜區(qū)域分析顯示,ORFV129蛋白不存在跨膜區(qū)域,推測其可能作為胞內蛋白而起作用(圖2-B)。通過WoLFPSORT在線比對軟件預測ORFV129蛋白的亞細胞定位,結果顯示蛋白質主要定位于細胞質(圖2-C)。

M. 相對分子質量標準(2 000 bp DNA ladder);P. 陽性對照;N. 陰性對照;1-5. ORFV129基因;A. ORFV129基因的PCR擴增;B. 重組質粒pMD-19T-ORFV129 的PCR擴增。
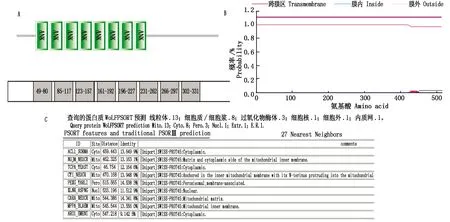
A. ORFV129蛋白的一級結構分析;B. ORFV129蛋白的跨膜區(qū)域分析;C. ORFV129蛋白的亞細胞定位分析。A. Predicted primary-structure of ORFV129 protein;B. Predicted transmembrane region of ORFV129 protein;C. Predicted subcellular localization of ORFV129 protein.
2.2 融合表達ORFV129基因真核重組表達載體的構建及鑒定
使用帶Hind Ⅲ、PstⅠ酶切位點的引物進行PCR擴增,結果顯示片段大小約為1 600 bp,與預期的片段大小(1 560 bp)相符合。隨后,膠回收后定向克隆至pEGFP-N1構建融合重組質粒pEGFP-ORFV129,經(jīng)雙酶切鑒定表明構建的質粒正確(圖3)。
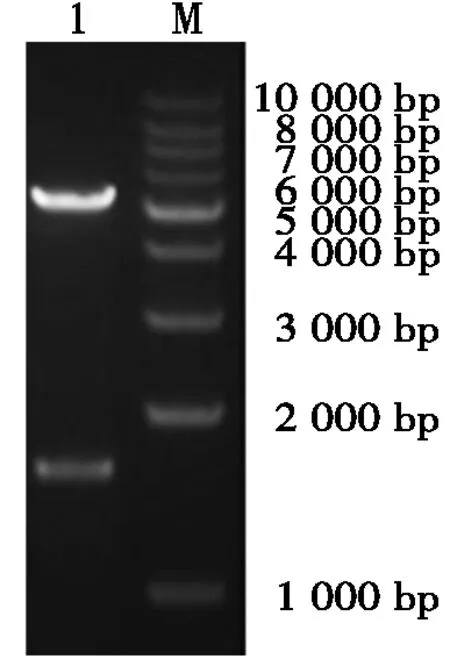
M. 相對分子質量標準(1 kb DNA ladder);1. pEGFP-ORFV129的Hind Ⅲ、Pst Ⅰ雙酶切鑒定。M. Relative molecular mass (1 kb DNA ladder);1. pEGFP-ORFV129 digested by Hind Ⅲ and Pst Ⅰ.
2.3 pEGFP-ORFV129重組質粒在睪丸細胞ORFV復制期間的mRNA表達分析
轉染12~24 h后,將融合載體pEGFP-ORFV129和空載體pEGFP-N1置于熒光倒置顯微鏡下,均能觀察到綠色熒光(圖4)。mRNA表達檢測發(fā)現(xiàn),細胞總RNA經(jīng)RT-PCR擴增后,可見與預期ORFV129基因大小相似的目的條帶(圖5),表明ORFV129基因在細胞中ORFV復制期間發(fā)生了轉錄。
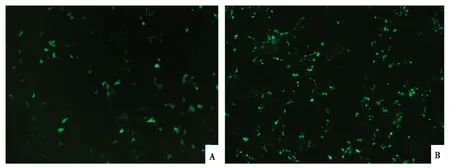
A. 轉染pEGFP-N1空載的新生山羊睪丸原代細胞;B. 轉染pEGFP-ORFV129的新生山羊睪丸原代細胞。A. Testis cells of newborn goat transfected with pEGFP-N1;B. Testis cells of newborn goat transfected with pEGFP-ORFV129.
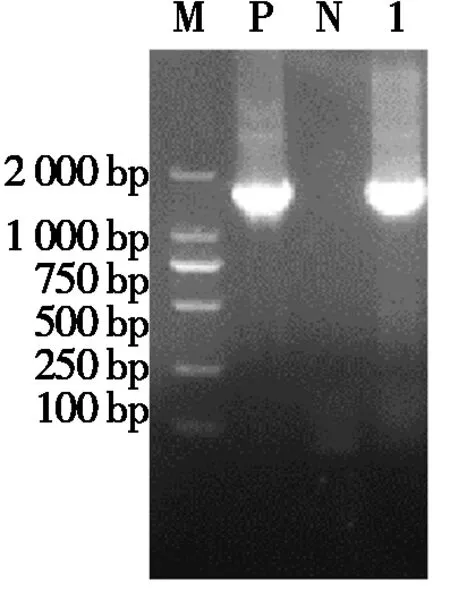
M.相對分子質量標準(2 000 bp DNA ladder);P.陽性對照;N.無模板對照;1.ORFV129基因。M.Relative molecular mass (2 000 bp DNA ladder);P.Positive product;N.No template control;1.ORFV129 gene.
2.4 融合蛋白ORFV129在羊睪丸細胞中的表達
將 pEGFP-N1和pEGFP-ORFV129重組質粒轉染羊睪丸原代細胞36 h后,Western Blot 結果顯示,空白對照組中沒有GFP蛋白的表達,而轉染了pEGFP-ORFV129重組質粒的山羊睪丸原代細胞中檢測到了ORFV129-GFP蛋白(75 ku 圖6 ORF129蛋白表達的Western Blot鑒定Fig.6 Western Blot identification of ORF129 protein 將 pEGFP-N1和pEGFP-ORFV129重組質粒轉染羊睪丸原代細胞24 h后,經(jīng)染料DAPI對細胞核進行藍染,激光共聚焦顯微鏡下分析ORFV129蛋白在細胞內的定位情況。結果顯示,在表達pEGFP-N1的細胞內,綠色熒光彌散性分布于整個細胞;而在表達pEGFP-ORFV129的睪丸細胞中,綠色熒光呈點狀地分布于細胞質(圖7)。 A. 轉染pEGFP-N1的睪丸細胞明場觀察;B.轉染pEGFP-N1的睪丸細胞綠色熒光觀察;C. DAPI 染色后細胞核;D. 圖B和C的合并;E. 轉染pEGFP-ORFV129的睪丸細胞明場觀察;F. 轉染pEGFP-ORFV129的睪丸細胞綠色熒光觀察;G. DAPI 染色后細胞核;H. 圖F和G的合并。 錨蛋白重復序列 (Ankyrinrepeat,ANKs )是普遍存在于生物體中的一種由33個氨基酸殘基基序組成的蛋白質序列模體,由2個α螺旋和1個Loop區(qū)構成,在細胞信號轉導、細胞骨架完整性、細胞周期調控、免疫應答、蛋白轉運等過程中均具有重要作用[13-14]。ANKs不僅存在生物體,也存在于痘病毒編碼的蛋白,而其他病毒幾乎不編碼錨蛋白。錨蛋白作為痘病毒編碼的蛋白中最大的家族,其特征是具有ANKs的N末端和1個F-Box樣的C末端結構域[15-17]。已有研究報道,痘病毒編碼的錨蛋白主要功能是可通過多層次的方式來影響NF-κB信號通路。例如,天花病毒(Variola,VARV)編碼的蛋白G1R,痘苗病毒(Vacciniavirus,VACV)編碼的蛋白CP77,兩者均可通過其N端的ANK重復基序,分別與NF-κB1/p105、NF-κB p65蛋白直接或間接結合抑制宿主NF-κB的活化[18-20]。2004年,Shisler等[21-22]發(fā)現(xiàn)VACV編碼的蛋白K1L可通過競爭與轉錄因子NF-κB的結合,阻止IκBα的降解,從而抑制NF-κB的活化。同時,該團隊也證實K1L蛋白能通過阻礙IKK的磷酸化從而抑制NF-κB的活化。因此,KIL蛋白準確的抑制機制仍然需進一步的研究。VACA A52和K7蛋白均能與腫瘤壞死因子受體相關分子6(TNF receptor-associated factor 6,TRAF6)相互作用,間接抑制NF-κB的激活[23]。羊口瘡病毒編碼的蛋白ORFV119,能通過與視網(wǎng)膜瘤蛋白相互作用抑制IKK的活化,從而抑制NF-κB信號通路的活化[24]。2014年,Lamb等[25]發(fā)現(xiàn),黏液瘤病毒(Myxomavirus,MYXV)編碼的錨蛋白M-T 5、M148R、M149R和M150R均能抑制NF-κB p65蛋白從細胞質遷移到細胞核,減少細胞核內NF-κB p65的聚集,抑制或者降低NF-κB信號通路的活性。同樣VACV編碼的錨蛋白K1L可通過對NF-κB信號途徑不同位點的作用而調控宿主的天然抗病毒免疫應答[26]。 對于ORFV編碼的5種錨蛋白,ORFV008蛋白可通過阻止IκBα蛋白磷酸化降解過程而抑制細胞NF-κB p65蛋白核移位,從而減弱細胞NF-κB信號通路的活化[27]。ORFV128蛋白在病毒感染早期表達且定位于宿主的細胞核,通過抑制IκBα蛋白磷酸化降解過程,阻止NF-κB p65蛋白核移位,進而抑制宿主NF-κB信號通路的活化[28]。可見,痘病毒內具有ANKs基序的錨蛋白,能通過對NF-κB信號途徑的不同位點的作用而調控宿主的天然抗病毒免疫應答。羊口瘡病毒ORFV129基因編碼的錨蛋白具有典型的N末端多個ANK重復序列和1個F-Box的C末端結構域。2013年,白剛等[10]利用真核表達系統(tǒng)對ORFV129基因編碼的蛋白進行表達并篩選出穩(wěn)定表達ORFV129基因的細胞株,為研究ORFV129蛋白調控宿主天然免疫應答的作用機制奠定了基礎。2016年,葛士坤等[29]對ORFV129基因序列及其編碼蛋白進行生物信息學分析,該結果為深入研究ANK在ORFV致病過程中的作用提供了基礎數(shù)據(jù)。對于ORFV編碼的129錨蛋白功能目前不清楚,這也正是本研究選擇ORFV129蛋白作為研究切入點的重要原因。本研究克隆了羊口瘡病毒ORFV129基因全序列,對其進行生物信息學分析;進一步構建真核表達載體pEGFP-ORFV129,并將其轉染新生山羊睪丸原代細胞,以空載體pEGFP-N1作為陰性對照,借助激光共聚焦顯微鏡觀察了ORFV129在細胞中的定位;亞細胞定位發(fā)現(xiàn)ORFV129分布于細胞質中。ORFV編碼的其他錨蛋白,如ORFV126蛋白表達后靶向定位于線粒體內,且ORFV126編碼的AR8和AR9對錨蛋白定位于線粒體內起著關鍵性的作用[30]。ORFV128蛋白表達后靶向定位于細胞核中[17]。ORFV129蛋白是否類似于ORFV126,定位于細胞質內的線粒體,或者其蛋白功能是否與ORFV128的不一致,目前仍不清楚,需要進一步的研究。 本研究成功克隆得到羊口瘡病毒ORFV129基因的全長,定位在新生山羊睪丸原代細胞細胞質中,這些結果為進一步研究ORFV129在誘導機體免疫應答中的作用奠定了基礎。
2.5 融合蛋白ORFV129-GFP的亞細胞定位

3 討論與結論
