玉米矮桿突變體A5的表型鑒定及轉(zhuǎn)錄組分析
2020-09-14 06:52:02楊睿張正楊麗莉張彥琴董春林常建忠
山西大學學報(自然科學版) 2020年3期
關(guān)鍵詞:差異
楊睿,張正,楊麗莉,張彥琴,董春林,常建忠
(山西農(nóng)業(yè)大學 山西有機旱作農(nóng)業(yè)研究院,黃土高原東部旱作節(jié)水技術(shù)國家地方聯(lián)合工程實驗室,有機旱作山西省重點實驗室,山西 太原 030031)
0 引言
玉米(Zeamays)是我國第一大糧食作物,是重要飼料來源,也是食品加工、醫(yī)療衛(wèi)生和再生能源等行業(yè)不可或缺的原材料,有較高的經(jīng)濟價值。玉米具有較強的耐旱、耐寒、耐貧瘠等抗逆性,是確保我國糧食穩(wěn)產(chǎn)和增產(chǎn)的主力軍,也是保障我國糧食安全的關(guān)鍵農(nóng)作物之一[1]。
矮化玉米株型緊湊,通風透光、適合密植,選育矮化玉米雜交種可有效減少玉米倒伏,穩(wěn)定和提高玉米產(chǎn)量[2]。矮桿突變體是作物育種中重要的“矮源”,研究表明導(dǎo)入矮桿和半矮桿基因,可大幅度提高玉米抗倒伏能力和產(chǎn)量[3]。1960年代的第一次“綠色革命”,就是發(fā)現(xiàn)并利用了矮桿基因,從而使水稻和小麥產(chǎn)量在世界范圍內(nèi)大幅度提升[4-5]。因此,篩選玉米矮桿突變體,鑒定玉米株高基因,研究玉米株高調(diào)控機制,一直是現(xiàn)代玉米育種和理論研究的重要內(nèi)容[6]。目前已報道玉米株高基因大多為隱性基因,如brachytic1(br1)、brachytic2(br2)、dwarf-1(d1)、dwarf-2(d2)、dwarf-3(d3)和dwarf-5(d5)等[7-9],少數(shù)為顯性基因,如dwarf-8(d8)、dwarf-9(d9)和dwarf-11(d11)等[10-14]。br2基因可使莖稈變粗,節(jié)間縮短,抗倒性增強;br2矮生系與多基因矮生系組配,其F1植株高度可進一步矮化。
玉米株高發(fā)育與赤霉素(Gibberellins, GAs)、生長素(Indole-3-acetic acid,IAA)和油菜素內(nèi)酯(Brassinosteroids, BRs)等植物激素密切相關(guān)[15]。植物矮化現(xiàn)象常與赤霉素代謝能力下降或赤霉素信號通路受阻有關(guān)。赤霉素在植物生長發(fā)育等許多方面起著至關(guān)重要的作用,如種子萌發(fā)[16]、莖節(jié)伸長[17]、葉片伸展[18]和花發(fā)育[19]等。根據(jù)突變體對赤霉素的敏感程度,可將玉米矮桿突變體分為赤霉素敏感型和赤霉素鈍感型。赤霉素敏感型突變體通過阻斷玉米體內(nèi)赤霉素的合成通路,導(dǎo)致赤霉素含量降低和玉米植株矮化,噴施外源赤霉素后株高可恢復(fù)正常;赤霉素鈍感型突變體的赤霉素合成通路正常,但赤霉素信號轉(zhuǎn)導(dǎo)通路受阻,噴施外源赤霉素后株高無法恢復(fù)正常。
前期我們利用甲基磺酸乙酯(EMS)誘變玉米骨干自交系PH6WC獲得一個玉米矮化突變體,命名為突變體A5,并通過連續(xù)自交后純合。本文對其進行了表型鑒定、遺傳分析、赤霉素敏感性、轉(zhuǎn)錄組分析等。研究結(jié)果為進一步利用該矮桿材料選育抗倒伏玉米新品種提供理論依據(jù),也為克隆玉米矮桿基因奠定基礎(chǔ)。
1 材料與方法
1.1 試驗材料
玉米突變體A5為玉米骨干自交系PH6WC經(jīng)EMS誘變所得,經(jīng)連續(xù)自交,已純合一致、穩(wěn)定遺傳。
1.2 試驗方法
1.2.1 玉米突變體A5的表型分析
于2018年5月在山西省農(nóng)業(yè)科學院東陽基地種植矮桿突變體A5和自交系PH6WC。種植密度為83 000株/hm2。突變體種植行長6.7 m,行距0.6 m,株距0.27 m,每行種25穴,每穴雙粒播種,田間正常管理。連續(xù)取小區(qū)內(nèi)正常的植株8株,測量自交系PH6WC和突變體A5相關(guān)性狀。調(diào)查的性狀主要有:株高、穗位高、雄穗分枝數(shù)、雄穗長、果穗穗長、穗粗、穗行數(shù)和行粒數(shù)等,采用t檢驗差異顯著性。
1.2.2 玉米突變體A5遺傳分析
將突變體A5與自交系PH6WC(野生型)雜交獲得F1代,F1代雜交種自交得到F2代,統(tǒng)計F2代分離群體中野生型和矮桿突變體的植株數(shù)目,采用卡方檢驗驗證突變體的遺傳規(guī)律。
1.2.3 外源赤霉素處理玉米突變體A5
試驗在山西省農(nóng)業(yè)科學院東陽基地進行。將突變體A5單行區(qū)種植,行長2 m,株距0.27 m,每行7穴,雙粒播種,常規(guī)肥水管理措施。設(shè)置突變體A5水處理組和赤霉素(GA3)處理組(GA3濃度:20、50和100 mg/L),自交系PH6WC組(野生型組),隨機區(qū)組設(shè)計,3次重復(fù)。于2018年4月25日種植,出苗后10 d開始噴施,之后每隔7 d處理1次,整個過程使處理液充分且均勻分布于植株表面。取中間5株測量株高,從苗期到抽雄完全為止,每隔7 d測定1次株高。
1.2.4 自交系PH6WC突變體A5的轉(zhuǎn)錄組分析
自交系PH6WC和突變體A5種植于24℃人工氣候箱,相對濕度70%,光照強度5 000 Lux。待幼苗長至5~6葉期取植株莖部樣本,提取總RNA,送上海派森諾生物科技股份有限公司測序。測序原始數(shù)據(jù)去接頭序列和低質(zhì)量序列,獲得高質(zhì)量序列(Clean data)。利用Bowtie2將Clean data與玉米參考基因組(Zea_mays.B73_RefGen_v4.42)進行比對,獲取基因組位置信息;用RPKM值(Reads Per Kilobase per Million mapped reads)對基因表達量進行標準化;通過DESeq對自交系PH6WC和突變體A5樣本中的基因表達進行差異分析,篩選閾值為Fold change>2(P-value<0.05和FDR<0.05),對差異表達基因進行GO功能注釋和KEGG通路富集分析。
2 結(jié)果與分析
2.1 玉米突變體A5的表型鑒定
突變體A5植株矮小,平均株高125.14 cm,較野生型PH6WC株高顯著降低37.99%(P<0.01)(圖1C)。穗位高度無顯著性變化。株型緊湊,莖稈直徑增加,葉片密集(圖1A),葉寬厚,節(jié)間數(shù)目與野生型無差異,節(jié)間長度縮短(圖1D),雄穗分枝數(shù)為1~4個,雄穗長29.60 cm,穗長12.23 cm,穗粗3.57 cm,穗行數(shù)12.80行,行粒數(shù)17.90粒,穗粗、穗行數(shù)、行粒數(shù)較野生型PH6WC有顯著增高,其余性狀無明顯差別(表1)。
2.2 玉米突變體A5的遺傳分析
玉米矮桿突變體A5和自交系PH6WC雜交得到的F1表型正常。F2分離群體中一個單穗197個后代中,有42株矮化植株;另一單穗179個后代中,鑒定到40株矮化型。卡方適合性檢驗表明此分離比符合孟德爾遺傳分離比率3∶1(表2),該矮化突變體是由隱性單基因控制。
2.3 外源赤霉素對突變體A5株高的影響
用不同濃度赤霉素(GA3)噴施玉米突變體A5,于全生育期測量玉米的株高,見表3。結(jié)果表明在未噴施GA3前(出苗后10 d),突變體A5的株高顯著低于PH6WC(P<0.01)。出苗后17 d,50 mg/L GA3處理的A5株高顯著高于PH6WC株高(P<0.05),20 mg/L和100 mg/L GA3處理的A5株高與野生型株高差異不明顯。在出苗后25 d、32 d和38 d,100 mg/LGA3處理的A5均高于野生型(P<0.05),20 mg/L和50 mg/L GA3處理的A5高度與野生型高度差異不明顯。出苗后53和60 d,野生型株高均極顯著(P<0.01)高于各濃度GA3處理的A5株高。以上結(jié)果表明噴施不同濃度的GA3均可提高突變體A5的株高,總趨勢為100 mg/L GA3處理組>50 mg/L GA3處理組>20 mg/L GA3處理組,但最終都未能使突變體恢復(fù)至野生型株高。
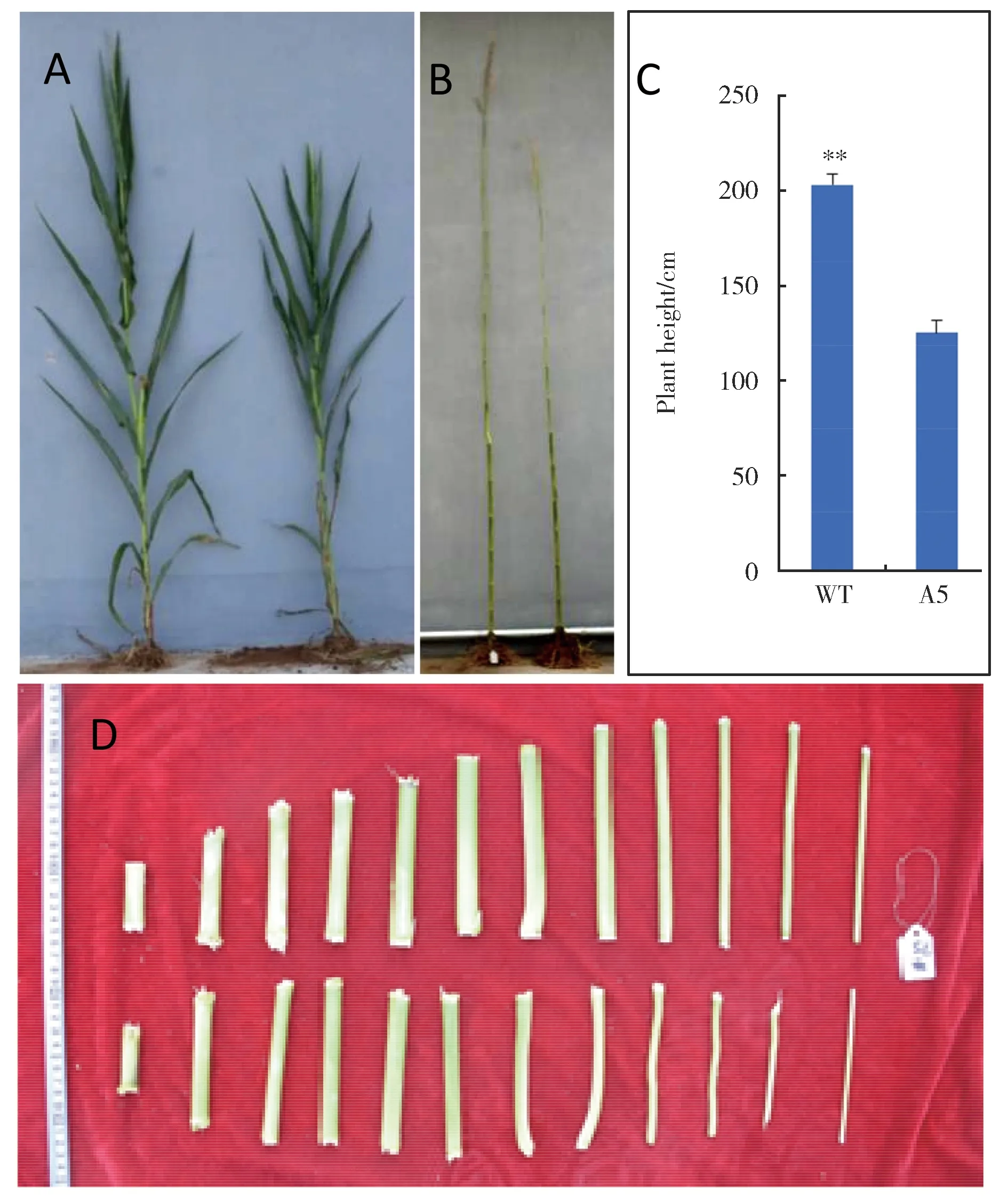
注:A:植株;B:莖稈;C:株高;D:節(jié)間;A-C (左:PH6WC;右:突變體A5);D (上:PH6WC;下:突變體A5)**P<0.01圖1 矮桿突變體A5和自交系PH6WC的表型比較Note:** Indicated extremely significant height differenceNote: A: Plant; B: Stems;C: Plant height; D: Internodes;A-C (Left: PH6WC; Right: mutant A5);D (Top: wild type; Bottom: mutant);**P<0.01Fig.1 Morphological comparison ofbetween mutant A5 and PH6WC
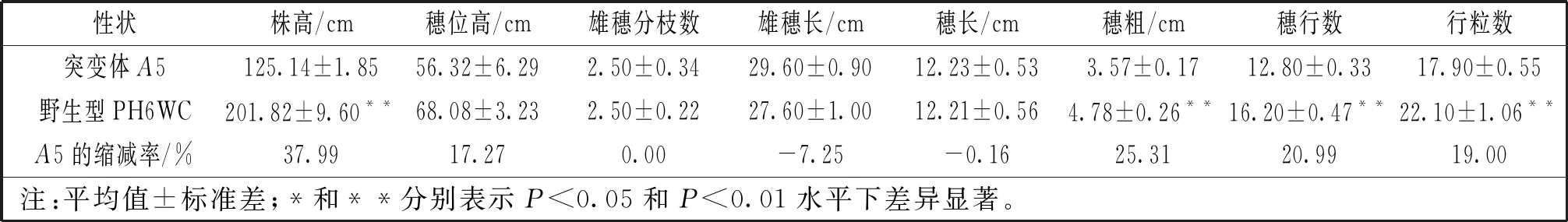
表1 矮桿突變體A5及其野生型PH6WC表型性狀比較

表2 F2群體中正常植株和矮桿植株的分離比
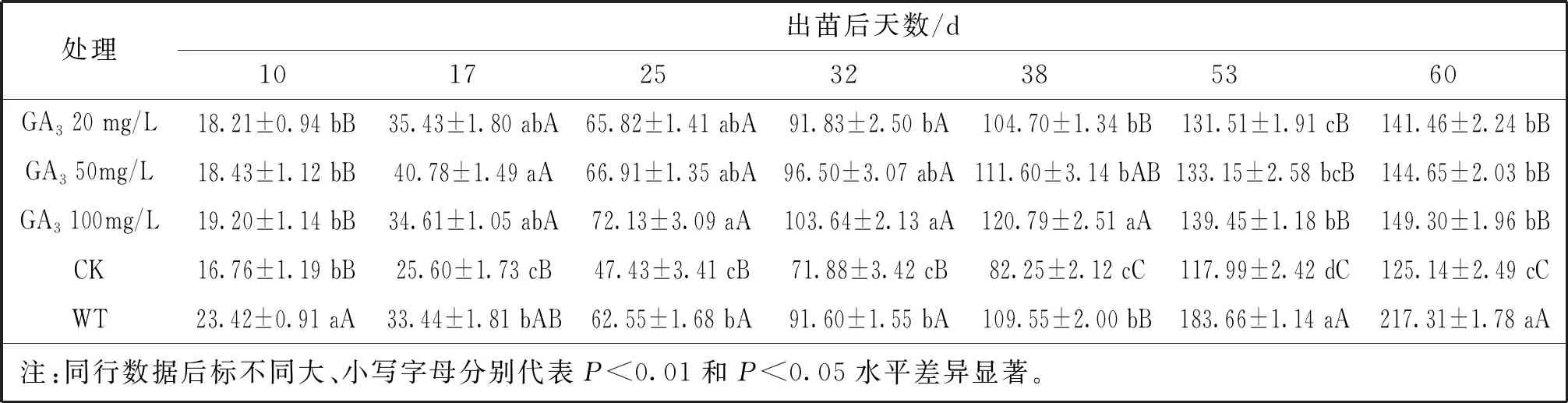
表3 GA3濃度梯度下突變體A5的株高變化
2.4 突變體A5的轉(zhuǎn)錄組分析
轉(zhuǎn)錄組分析共鑒定出2 040個差異表達基因,其中下調(diào)表達基因1 332個(4.39%),上調(diào)表達基因708個(2.33%)(圖2)。差異表達基因中發(fā)現(xiàn)一些與株高發(fā)育相關(guān)基因,如d1(Zm00001d039634)和ga2ox(Zm00001d037565)等,還有與細胞分裂有關(guān)的基因,如tan1(Zm00001d038060)等。對差異表達基因進行GO富集分析發(fā)現(xiàn)共有713個差異表達基因獲得功能注釋,其中463個基因參與生物學過程,包括的功能條目DNA復(fù)制(GO:0006260)、細胞過程(GO:0009987)、發(fā)育進程(GO:0032502)和細胞發(fā)育過程(GO:0048869);118個基因參與細胞組分包括細胞器(GO:0043226)、微管細胞骨架(GO:0015630)和細胞外周(GO:0071944);132個基因參與分子功能包括蛋白激酶調(diào)節(jié)活性(GO:0019887)、錯配DNA結(jié)合(GO:0030983)、錯配修復(fù)復(fù)合體結(jié)合(GO:0032404)(圖3)。
進一步采用KEGG分析(Kyoto Encyclopedia of Genes and Genomes)對玉米差異表達基因進行注釋和分類,參與DNA復(fù)制、錯配修復(fù)和同源重組等生理過程的23個、10個和12個關(guān)鍵基因獲得富集,且均為上調(diào)表達(圖4)。
3 討論
株高對作物種植密度、抗倒性和光合效能有著非常重要的影響。玉米矮化突變體植株矮小、株型緊湊、抗倒伏能力強,是培育耐密植玉米雜交種的重要種質(zhì)材料。然而目前生產(chǎn)上應(yīng)用的玉米矮桿材料(基因)比較單一,不利于玉米產(chǎn)量和品質(zhì)的進一步提升,發(fā)掘和創(chuàng)制新矮桿種質(zhì)資源是玉米遺傳育種的重要研究方向。本研究所用玉米矮桿突變體A5是經(jīng)EMS誘變玉米自交系PH6WC獲得玉米矮化突變體,株高較野生型顯著降低了37.99%,總體表現(xiàn)為株型緊湊、耐密植,具有重要的研究和實際應(yīng)用價值。遺傳分析表明A5表型受一對隱性單基因控制,目前已報道的矮化單基因突變體中,絕大多數(shù)為隱性單基因控制[20-22],如d1、d2、d3、d5和antherear1(an1),突變體A5與它們的等位關(guān)系,需進一步分析證明。
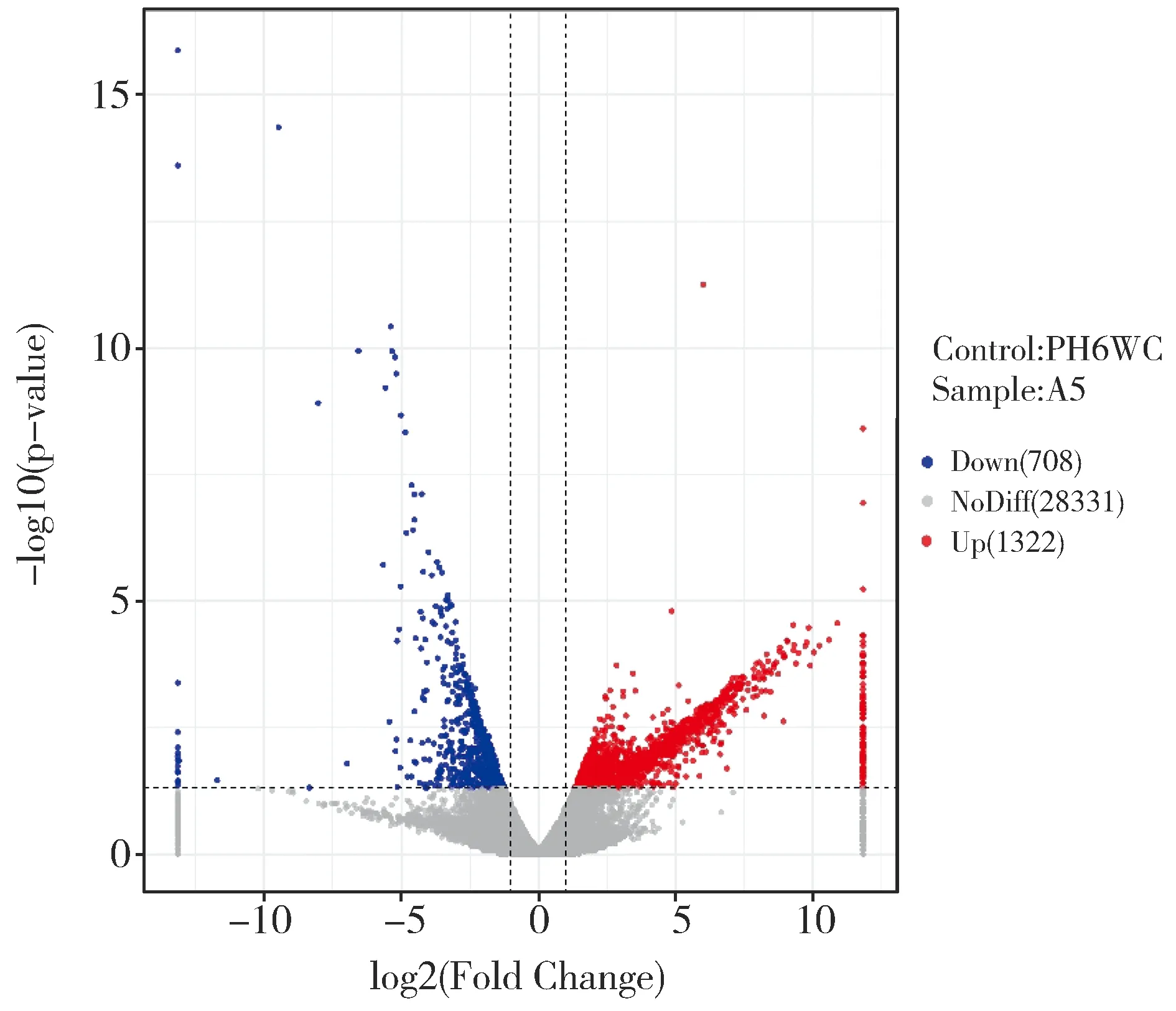
圖2 自交系PH6WC和突變體A5的基因差異表達分析火山圖Fig.2 Volcano plot of differential gene expression analysis between PH6WC and mutant A5
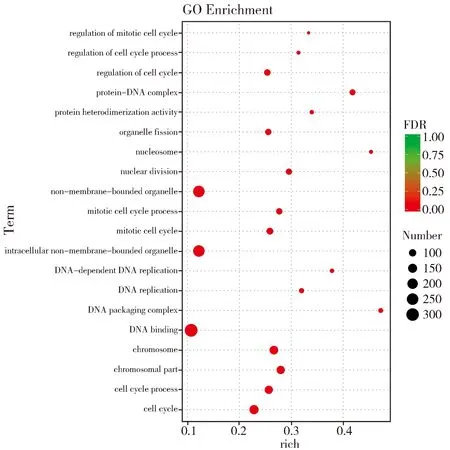
圖3 差異表達基因GO分析Fig.3 GO analysis of the differentially expressed genes
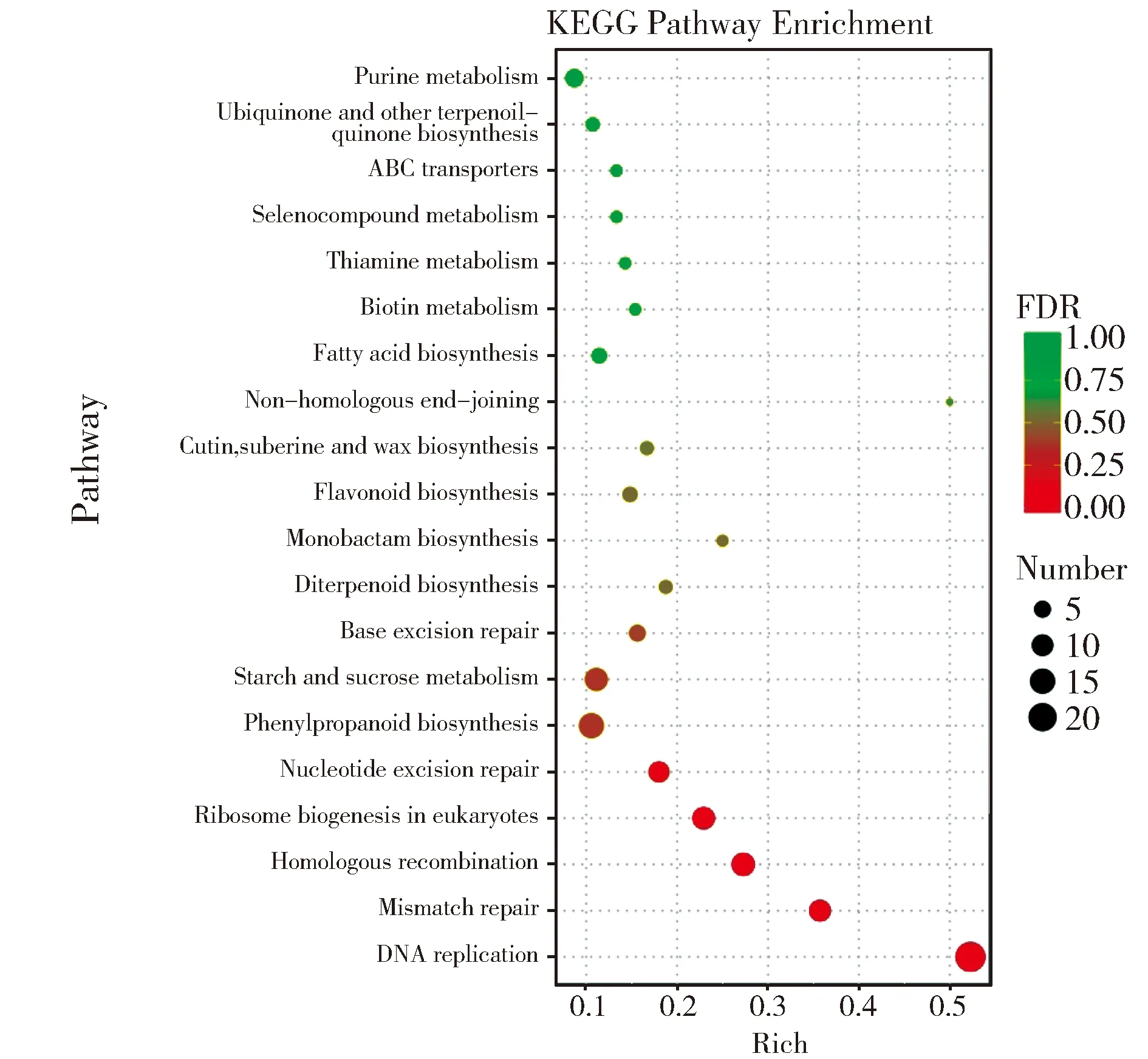
圖4 差異表達基因KEGG富集分析Fig.4 KEGG enrichment analysis of differentially expressed genes
GAs作為內(nèi)源激素之一,與IAA和BRs一起參與植物株高調(diào)控過程[23]。已發(fā)現(xiàn)玉米矮桿突變體中,D1[8]、Na1[21]、Dt[24]、D2003[25]、Dm1[26]、D8-1023[11]和D11[9]為赤霉素敏感型,外施激素后表型可恢復(fù)至野生型。本研究A5植株噴施不同濃度GA3后株高有所增加,總趨勢為100 mg/L GA3處理組>50 mg/L GA3處理組>20 mg/L GA3處理組,但均未達到野生型高度。這一結(jié)果表明,突變體A5只能對外源GA3做出有限響應(yīng),因此推測該突變體的矮化表型不僅僅是赤霉素合成缺陷造成的[27]。
高通量測序技術(shù)成本低、準確性高和耗時短,已廣泛用于各物種的組學分析[28]。對野生型和突變體進行轉(zhuǎn)錄組測序分析,有助于在全基因組水平上發(fā)掘?qū)τ衩字旮哂兄匾饔玫墓δ芑騕28],為矮桿候選基因的確定和克隆提供依據(jù)。本研究以突變體A5和野生型PH6WC于V5期(5葉期)玉米幼莖為材料,進行轉(zhuǎn)錄組分析。結(jié)果表明與野生型相比,A5中共2 040個差異表達基因(708個上調(diào)表達基因,1 332個下調(diào)表達基因),其中包含d1(Zm00001d039634)和ga2ox(Zm00001d037565)等赤霉素合成相關(guān)基因。矮桿基因d1編碼GA3-氧化酶,催化生物活性GA合成的最后一步[20],本研究中d1的表達量在A5中比野生型顯著上調(diào)2.21倍(P=0.007);ga2ox編碼赤霉素2-氧化酶(Gibberellin 2-oxidase),其在A5中較對照下調(diào)表達3.18倍(P=0.01),GA2ox作用于有生物活性的GA1和GA4,通過調(diào)控GAs的水平來調(diào)節(jié)植物生長,如使植物節(jié)間縮短、矮化[29]。此外,還有與植物的細胞生長、分裂和分化有關(guān)的基因,如tan1(Zm00001d038060),Tan1是玉米細胞質(zhì)分裂空間調(diào)控所必須的微管結(jié)合蛋白,是一種能直接與微管結(jié)合的高堿性蛋白質(zhì)[30],其參與細胞分裂中細胞骨架結(jié)構(gòu)的定向及骨架空間的排列[30- 31]。
GO富集分析表明與野生型PH6WC相比,突變體A5的差異表達基因富集在DNA復(fù)制、錯配修復(fù)等關(guān)鍵生理過程。DNA復(fù)制是細胞分裂過程中的重要步驟,而錯配修復(fù)是DNA復(fù)制過程中的重要“保真機制”,相關(guān)基因突變可影響植物株高發(fā)育。如水稻OsKu80基因在DNA雙鏈斷裂修復(fù)和端粒調(diào)控中發(fā)揮重要作用,該基因突變可導(dǎo)致植株矮化,花不育.[32]。對差異表達基因中的一些重要基因進行表達分析和功能鑒定將是我們未來工作的重點之一。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(shù)(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數(shù)英綜合(2019年8期)2019-08-27 02:23:00
中國特種設(shè)備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現(xiàn)代語文(2016年21期)2016-05-25 13:13:50
