中國西南地區熱帶森林演替序列碳動態
2020-09-17 00:01:16步巧利譚正洪張一平
生態學報 2020年15期
步巧利,譚正洪,張一平
1 廣東省佛山市氣象局, 佛山 528000 2 中國科學院熱帶森林生態學重點實驗室, 昆明 650223
熱帶森林因其復雜的群落結構,多樣的物種和巨大的生物量而著稱。在熱帶森林研究享有盛譽的生態學家Kira Tatuo將熱帶森林生態學的研究喻為生物學研究的“臨界狀態”[1]。熱帶森林在全球碳平衡和碳循環中也扮演著十分重要的角色。雖然熱帶森林只占有約22%的潛在陸地植被分布面積[2],卻擁有著全世界陸地生產力的43%[3]和碳儲量的59%[4]。
在地球系統碳循環的研究中,熱帶地區和熱帶森林也是最不確定的[5]。無論是使用渦度相關技術,生物調查和生理生態尺度擴展,還是使用大氣CO2濃度反演,進行熱帶雨林碳平衡的評測都是一個挑戰[6]。因此,在回答全球碳循環“失匯”的問題上,熱帶森林也有著顯著的重要地位。
隨著人口的增長,社會的發展和經濟的膨脹,原生森林在不斷的減少,取而代之的是經濟價值較高的人工林,如橡膠林。同時,一部分破壞后遺棄的林地,次生林隨著時間生長起來。關于次生林演替過程中的碳儲量和碳平衡的變化卻鮮為人知,特別是在熱帶地區。因此針對中國西南地區熱帶森林演替碳動態的研究非常有必要,且可為全球碳循環的“失匯”問題尋找答案。
本文利用3個熱帶次生林的樣地的數據,探討:(1)處于不同演替狀態的熱帶次生林的碳儲量狀況如何;(2)森林的凈碳蓄積,死亡碳損失和更新碳增長等碳動態如何;(3)次生林在未來的固碳方面的潛力如何。
1 研究方法
1.1 研究地概況
研究樣地位于云南省西雙版納傣族自治州勐臘縣勐侖鎮(101°16′E, 21°55′N)。受大氣環流、地理位置及地方氣候的影響,地處熱帶北緣的西雙版納1年中有霧涼季(11—翌年2月)、干熱季(3—4月)和雨季(5—10月)之分。該地的年平均溫度為21.5℃,年平均降雨量為1492 mm。地帶性植被類型為熱帶季節雨林和季雨林,在海拔較高(800—1800 m)的山地分布有南亞熱帶季風常綠闊葉林, 在部分中山的局部地段分布有山地雨林。該地區是我國熱帶雨林集中分布的重要區域,同時也是東南亞熱帶雨林分布的最北緣。由于地處古熱帶植物區系向泛北極植物區系的過渡區及東亞植物區系向喜馬拉雅植物區系的過渡區,該區的生物區系成分十分復雜、物種多樣性高度富集。由于特殊的地形地貌,氣候的立體分異相當大,其東、北、西三面高,在一定程度上阻擋了西北方來的冷氣流,使得最冷月平均溫度并不低,彌補了積溫的不足。在冬季低山溝谷及低丘上的濃霧又彌補了降水的不足。因而在該地區的低山溝谷及低丘上,形成了熱帶濕潤氣候,具有熱帶雨林發育的條件。
1.2 調查樣地和樣地調查
本文選擇了西雙版納熱帶森林生態站調查樣地中關于次生林的三個樣地用于本研究。樣地的基本情況見表1。
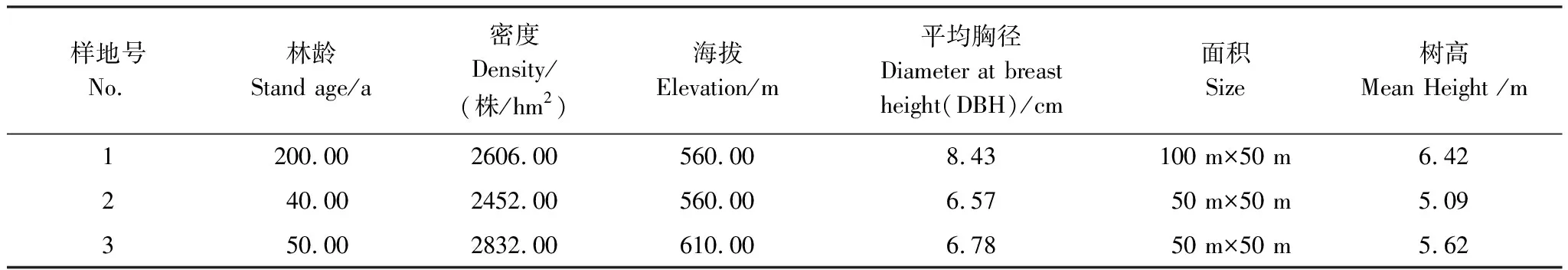
表1 三塊研究樣地的概況
樣地的每木調查按照CERN(中國生態系統研究網絡,Chinese Ecosystem Research Network)規定完成,利用每木調查法觀測胸徑、高度,1次/5 a(人工林或幼齡次生林1次/2 a或1次3 a,輪換觀測),按Ⅱ級樣方統計密度,同時在調查樣地內安置收集框來確定凋落物量,同時在調查樣地內安置收集框來確定凋落物量,具體調查細節請登陸CERN網站,參考西雙版納生態站鏈接(http://bnf.cern.ac.cn/)。樣地的具體情況如下:
樣地(1)是原生熱帶雨林樣地,位于西雙版納熱帶植物園的遷地保護區內,該樣地的植被受到一定程度的干擾(擇伐),以絨毛番龍眼(Pometiatomentosa)、箭毒木(Antiaristoxicaria)、蔥臭木(Dysoxylumexcelsum)、梭果玉蕊(Barringtoniafusicarpa)、窄序崖豆樹(Millettialeptobotrya)等占優勢的熱帶季節雨林。群落蓋度為90%,群落的垂直結構可分為喬木層、灌木層和草本層,其中喬木層又可分為2個亞層(喬木I、II亞層)。喬木I亞層(喬木上層)高約20—25 m,主要由絨毛番龍眼(Pometiatomentosa)、箭毒木(Antiaristoxicaria)、勐侖翅子樹(Pterospermummenglunense)等組成;喬木II亞層(喬木下層)高約5—20 m,主要由蔥臭木(Dysoxylumexcelsum)、窄序崖豆樹(Millettialeptobotrya)、小葉紅光樹(Knemaglobularia)、黑皮柿(Diospyrosnigrocortex)等組成;灌木層高約1—3.5 m,主要由大花哥納香(Goniothalamusgriffithii)、假海桐(PittosporopsisCraib)、山木患(Harpulliacupanioides)等組成;草本層主要由長葉實蕨(Bolbitesheteroclita)、小葉樓梯草(Elatostemaparvum)、假斜葉榕(Ficussubulata)等組成,層間附(寄)生植物豐富。
樣地(2)是熱帶季節雨林破壞后形成的以窄序崖豆樹為優勢樹種的次生林。群落蓋度較高,達到90%,群落高度為10—15 m,群落的垂直結構可分為喬木層、灌木層和草本層;喬木層主要由窄序崖豆樹(Millettialeptobotrya)、披針葉楠(Phoebelanceolata)、短藥蒲桃(Syzygiumbrachyantherum)、椴葉山麻桿(Alchorneatiliaefolia)等組成;灌木層主要由滇南九節(Psychotriahenryi)、彎管花(Chasaliscurviflora)等組成;草本層主要由南山花(Prismatomerisconnata)、馬唐(Digitariasanguinalis)等組成。
樣地(3)是熱帶季節雨林破壞后發育起來的次生林。群落蓋度達到80%,群落高度為10—12 m,群落的垂直結構可分為喬木層、灌木層和草本層,其中喬木層主要由清香木姜子(Litseaeuosma)、傘花木姜子(Litsea umbellata)、黃丹木姜子(Litseaelongata)、印度栲(Castanopsisindica)、鵝掌柴(Scheffleraoctophylla)、披針葉楠(PhoebeNees)、短藥蒲桃(Syzygiumbrachyantherum)、滇銀柴(Aporusayunnanensis)等組成;灌木層主要由大花哥納香(Goniothalamusgriffithii)、粗毛榕(Ficushirta)等組成;草本層主要由滇南素馨(Jasminumwangii)、銅錘玉帶草(Pratia nummularia)、南山花(Prismatomerisconnata)等組成。
1.3 數據處理
按照已經建立的異速生長方程[7-8]計算群落胸徑大于2 cm林木的生物量。已有研究表明,群落的生物量主要集中在胸徑大于2 cm的林木。因此本文以大于2 cm的林木的總生物量來代替整個群落的生物量。使用普遍的0.5為轉換系數,將生物量轉化為碳儲量[6]。
使用Sigmoidal函數擬合演替年份與群落的碳儲量。部分文獻針對群落碳動態中的幾個主要參量做了說明[9-10]。森林演替年齡的確定如下:其中2,3,4,5年4個林齡的數據引自馮至立等[8];5,10,14,22年4個林齡的數據引自唐建維等[7];本研究的樣地1,2和3的林齡根據查閱文獻和資料記載[6,11-13],并結合群落的物種組成和胸徑頻度分布,將樣地1、2和3的林齡分別定為200年,40年和50年。
2 結果與討論
在森林的演替過程中,森林的胸徑分布頻度從近正態分布逐漸向小徑級的偏態分布發展(圖1)。也就是說,隨著演替的進展,小徑級林木所占的比例越來越高。
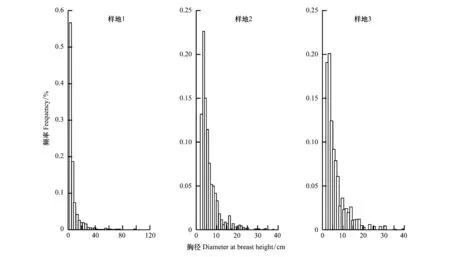
圖1 調查樣地內不同胸徑的林木的頻度分布(胸徑大于2 cm)
2.1 處于不同演替狀態的熱帶森林的碳儲量
不同演替階段西雙版納熱帶森林的碳儲量如圖2所示。整個演替過程可以被劃分為5個階段:干擾發生(I);干擾后的迅速生長期(II);較慢的生長期(III);儲量近穩定期(IV);下一次干擾發生(V)。
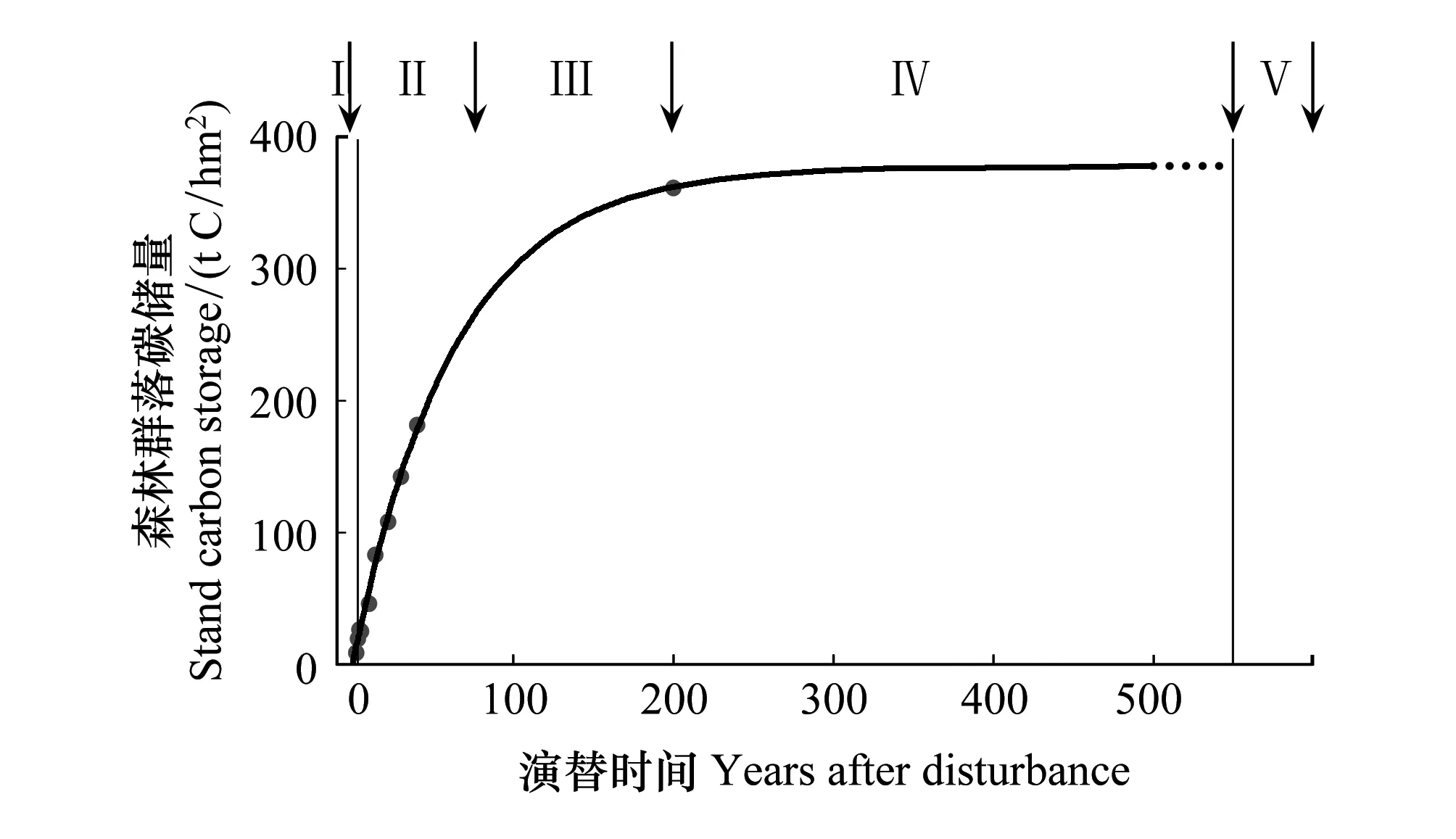
圖2 圖解演替過程中森林的碳動態
年輕的森林(Young forest)固碳能力最強是一個較為普遍的觀點。如圖2,西雙版納熱帶森林階段II的固碳能力明顯強于其他各階段的固碳能力。我們希望在此強調的是不論在森林演替的哪個階段,大樹(不同演替階段關于大樹沒有絕對的定義,對于原生熱帶雨林而言,大樹通常被定義為胸徑大于70 cm的林木)在森林固碳方面都占有非常重要的地位。
對于同一片森林,根據植物的生長規律,隨著胸徑的增長,胸徑的相對增量越小(圖3)。然而,從胸徑絕對增量來看,胸徑大的林木卻發揮著主導性的地位。生物量與胸徑之間存在較好的冪函數關系,方程表達式分別為y=0.6x1.14,R2=0.58;y=0.5x1.09,R2=0.525(圖3)。因此胸徑的絕對增量對生物量的貢獻按照冪次放大。因此,大樹在影響森林碳蓄積方面有著非常重要的地位。
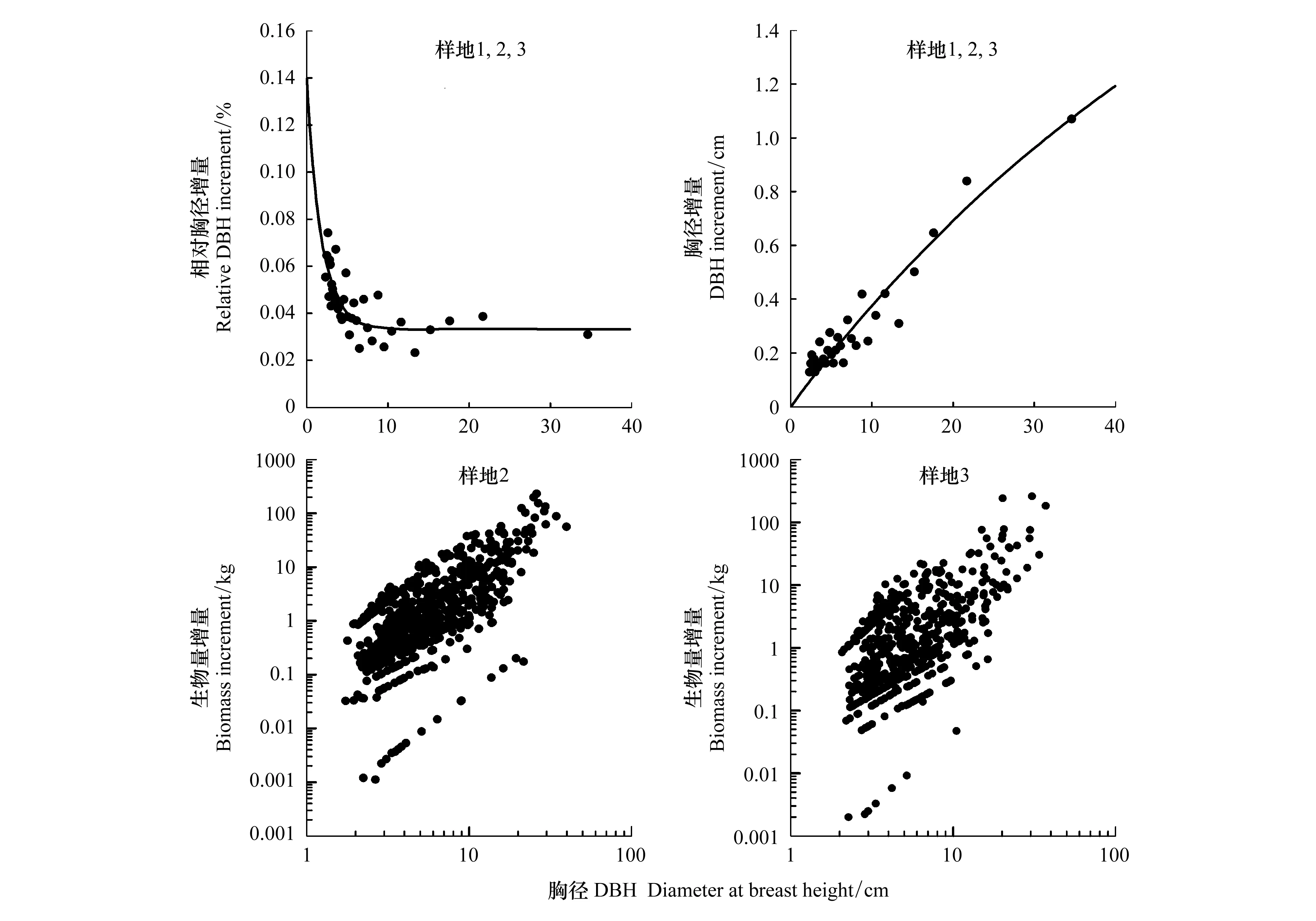
圖3 不同胸徑林木的相對胸徑增長,胸徑絕對增長,生物量增量
關于森林演替末期的碳平衡狀態是至今爭論較大的一個研究熱點。根據Gorden的預測[14],在森林自然演替的末期(稱老年林或過熟林(old-growth forest)),森林處于光合碳吸收和呼吸碳釋放的均等狀態,生態系統既不吸收碳亦不釋放碳。然而,實測數據卻并不支持該預測[15-16]。以西雙版納熱帶雨林為例,按照Odum的預測,當生態系統位于IV演替階段時,生態系統的碳儲量達到最高值,植被的碳蓄積過程幾近停止;然而,在對一塊未經明顯人為干擾的永久研究樣地的數據進行分析時,我們發現因為森林內林木的生長,森林每年以3.50 tC/m2的速度蓄積著碳[6]。根據全世界范圍內廣泛收集的數據建立的數據庫,Luysseart等[16]也得出世界上多數森林并沒有達到生物量的極限最大值。Luysseart等[16]認為,關于Odum的預測和實測數據之間的不吻合,最重要的原因在于老年林(old-growth)的定義。自然界的不同尺度的干擾是普遍存在的,森林并不能隔絕于干擾之外存在。小的干擾,如森林林窗的形成,將大大的改變森林內部的光環境,進而影響輻射,能量,水分平衡等生物物理過程,最終波及森林的碳動態。大的干擾,如火災和砍伐,將導致森林的次生演替,對森林的碳動態產生不可逆轉的改變。因而,我們認為的老年林并非Odum理想狀態下的生態系統。
2.2 森林凈碳蓄積,死亡碳損失和更新碳增長等碳動態
使用生物調查的數據來探討森林生態系統的凈生物量增量已經比較成熟:如單木跟蹤方法(track individual method)[17-18]。然而,多數的研究只提供一個樣地凈生物量增量的結果,很少有編制生物量動態表。不論使用生物量來計算生態系統的凈初級生產力還是凈生物量增量,編制森林碳動態表格是必要的。從西雙版納熱帶次生林的生物量動態表格(表2,3),我們可以看出研究樣地3中導致生物量損失的重要因素就是大徑級(>60 cm)的林木發生了死亡,樣地3中導致生物量損失的重要因素經查閱大量資料和咨詢相關研究人員后發現造成該死亡的主要原因則是干擾。
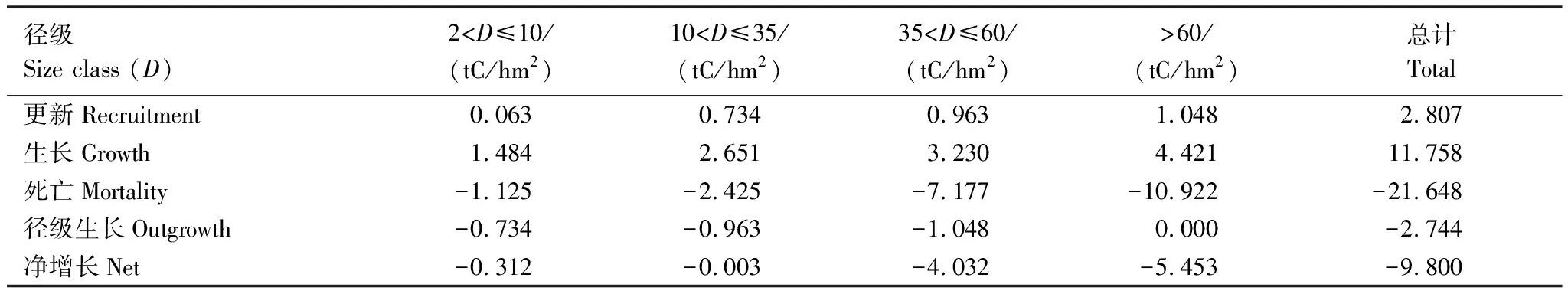
表2 樣點2喬木生物量在2003—2007年之間的變化
森林凋落物量是按照生物調查法計算生態系統凈初級生產力和探討森林碳動態的另一個重要方面。與蓄積在林木中周轉時間長的碳相比,凋落物可以在更小的一個時間尺度內影響碳動態。從2003—2007年樣地2和3的凋落物動態如圖4所示。與原生林相類似,在月尺度上,有兩個凋落峰,主峰出現在干熱季(3—4月),次峰在雨季末期(10月)。但是,次生林的凋落峰相對于原生林而言不太明顯。次生林的樹高較原生林低被推測為導致該現象的原因。特別需要指出的是,2003—2004年度西雙版納勐侖地區經歷了過去50年降雨最少的一年。由于2001和2002年為兩個豐水年,土壤含水量最低值并不是出現在2003年,而是在2004年。水分虧缺導致了2004—2005年度旱季凋落物的迅速增加,凋落物的增加,促進了森林的呼吸異化作用,增加了碳排放,這個結果也表現在渦度碳通量的觀測結果上[6,19-20]。同時,該結果也表明,造成西雙版納凋落物季節和年間動態上,干旱也起著非常重要的作用。
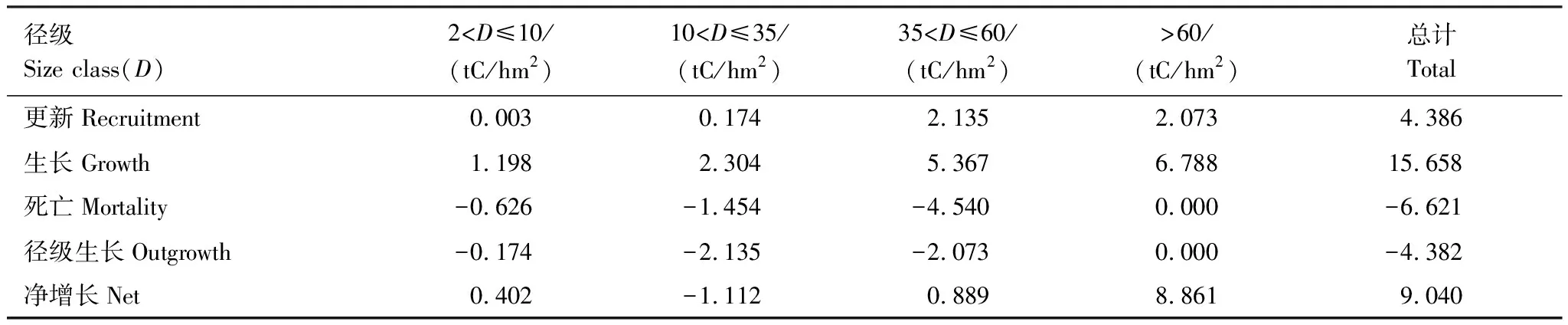
表3 樣點3喬木生物量在2003—2007年之間的變化
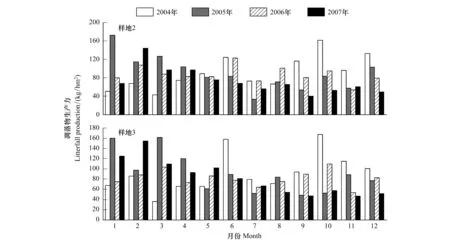
圖4 西雙版納熱帶次生林的凋落物月動態
2.3 對熱帶森林碳蓄積的啟示
雖然我們強調熱帶原始生林在固碳方面的作用,但是次生林在固碳方面的作用也不可小覷,特別是在快速生長期(圖2)。從FLUXNET(碳通量觀測網絡)的數據來看,干擾是導致生態系統成為碳源的主要原因[21],未遭受明顯干擾的森林基本上都表現出碳匯作用[22]。干擾可以導致原始林成為碳源[8],同時也使次生林的碳蓄積表現為負值(死亡的碳損失大于生物量增長的碳蓄積)(表2)。因此,不論是原生林還是次生林,加強森林的管理,竭力杜絕人為干擾,減弱自然干擾的影響,是促進森林碳蓄積的基礎。
3 結論
(1)熱帶森林在演替過程中,森林的碳儲量可以被劃分為五個階段:干擾發生(I);干擾后的迅速生長期(II);較慢的生長期(III);儲量近穩定期(IV);下一次干擾發生(V)。在熱帶森林的演替過程中,存在一個迅速生長期,該生長時段森林的固碳速度達到最高。因此熱帶次生林在森林固碳方面也發揮著不可忽略的作用。
(2)不論原生林還是次生林,干擾是導致森林成為碳源的主要原因,因此加強森林的管理,竭力杜絕人為干擾,減弱自然干擾的影響,是促進森林碳蓄積的基礎;推薦使用編制森林生物量動態表,以便詳細獲知造成森林碳蓄積或者損失的可能原因。
(3)熱帶次生林的凋落物動態與原生林的相類似,但是其季節趨勢較弱。干旱事件是影響凋落物的季節和年間動態的原因,也是短時間尺度上影響碳平衡的一個重要因子。
