鈾對蠶豆幼苗光合特性和呼吸代謝的毒害機理
2020-10-09 03:43:28劉澤偉賴金龍李俊柯丁峰張宇羅學剛
農業環境科學學報 2020年9期
劉澤偉,賴金龍,李俊柯,丁峰,張宇*,羅學剛
(1.西南科技大學生命科學與工程學院,四川 綿陽621010;2.西南科技大學環境與資源學院,四川 綿陽621010;3.西南科技大學生物質材料教育部工程研究中心,四川 綿陽621010)
核電及國防軍工對核燃料的需求推動鈾礦冶快速發展[1],鈾礦冶產生的大量鈾廢石、鈾尾礦中殘存的放射性核素及伴生重金屬被釋放進入環境,對環境造成嚴重的放射性及重金屬雙重污染。鈾是典型的重金屬污染元素,具有半衰期長、輻射強度大、衰變子體多等特點,其毒性效應包括輻射毒性和化學毒性雙重效應[2-3]。鈾在自然界中主要以化合物形式存在,其可溶態易被生物吸收,并沿著食物鏈進入人體,對人體造成內輻射,嚴重危害人體健康[4]。因此,鈾礦冶周邊環境鈾污染的修復與治理已成為亟需解決的問題。
近年來,利用植物根系對鈾的提取和固定來修復鈾污染的土壤或水體,已受到國內外學者的廣泛關注[5-6]。然而,鈾作為一種非必需、難轉移元素,極易附著、累積在植物根部[7-8],其在植物根部的過量累積,已對植物造成明顯的毒害作用,如植物生長緩慢、根系壞死[9],這也嚴重影響了植物對鈾污染的修復效率。因此,研究鈾對植物的毒害機理有助于了解植物對鈾的耐受性及修復植物的篩選,對利用植物修復鈾污染具有重要意義。
蠶豆等豆科植物除了根系能夠吸收重金屬外,還能改善土壤環境,具有修復土壤重金屬污染的潛力[10-11]。據研究報道蠶豆可作為研究重金屬對植物毒害作用的一種模式植物[12]。光合作用和呼吸代謝是植物生長發育過程中兩個重要的生理代謝過程,受到重金屬脅迫的影響[13-17]。已有研究表明,Cd、Pb等重金屬脅迫對植物的光合作用會造成一定的影響[13,18]。例如,趙魯雪等[8]研究發現鈾脅迫下四季豆的凈光合速率下降,而大葉菠菜、紅圓葉莧的凈光合速率上升,說明不同植物對鈾脅迫的適應能力存在差異。另外,有研究表明重金屬脅迫會影響植物根系的呼吸代謝過程[16,19-20]。例如,吳恒梅等[15]研究發現絲瓜種子的根系活力在低濃度鎘脅迫下有所增強,而在高濃度鎘脅迫下會被抑制。盡管已有報道顯示重金屬脅迫會影響植物的正常生理代謝過程,但關于鈾對植物光合作用及呼吸代謝影響的分子機理鮮有報道。
隨著轉錄組測序技術的發展,利用該技術從分子水平上分析植物逆境脅迫下的毒害機理或耐受機制方面的研究備受關注[21-22]。因此,本文采用轉錄組測序技術分析高濃度鈾脅迫下蠶豆幼苗光合作用和呼吸代謝相關基因的差異表達,并結合一些生理生化分析,以系統地闡釋鈾對蠶豆的毒害機理,對了解蠶豆對鈾的耐受性和修復植物的篩選具有參考意義。
1 材料與方法
1.1 植物培養與處理
本實驗以蠶豆為研究對象,種子購于綿陽豐益河種子有限公司。隨機挑選飽滿、種皮未破損的蠶豆種子用蒸餾水沖洗3次后,浸種24 h。將種子置于人工培養氣候箱(溫度25℃,濕度80%)中進行發芽。挑選長勢一致(胚根長度約2 cm)的發芽種子移植到定植籃中(每個定植籃放3粒種子),再將定植籃放置在含有改良霍格蘭營養液的植物組織培養瓶中。每3 d更換1次營養液,直至蠶豆長出兩片真葉。改良霍格蘭營養液組成如下:101.1 mg·L-1KNO3、236.15 mg·L-1Ca(NO3)2·4H2O、98.59 mg·L-1MgSO4·7H2O、16.01 mg·L-1NH4NO3、13.61 mg·L-1KH2PO4、鐵 鹽 溶 液(1.345 mg·L-1Na2-EDTA和1.112 mg·L-1FeSO4·7H2O),以及微量元素(包含:0.569 mg·L-1H3BO3、0.356 mg·L-1MnCl2·4H2O、0.043 mg·L-1ZnSO4·7H2O、0.01 mg·L-1CuSO4·5H2O和0.018 mg·L-1H2MoO4·H2O),pH為5.5。植物生長條件為:白天25℃/黑夜18℃,白天12 h/黑夜12 h,光照強度為5500 lx。
以硝酸鈾酰[238U,UO2(NO3)2·6H2O,分析純試劑]為鈾源,設計鈾污染濃度為0(對照)、5、10、15、20、25μmol·L-1,每組重復3次。本實驗采用不含磷的改良霍格蘭營養液制備不同濃度的鈾溶液,以避免營養液中的磷酸鹽與鈾酰離子形成不溶性沉淀。隨機選擇生長狀況基本一致的蠶豆幼苗(長至兩片真葉階段),用不同濃度的鈾溶液代替營養液,在與植物培養相同的條件下處理72 h。同時,每日在葉片上噴施1次磷溶液(0.1 mmol·L-1KH2PO4),以避免幼苗缺磷的癥狀。鈾處理72 h后,用蒸餾水沖洗幼苗根系3次,用濾紙吸干多余水分,液氮冷凍,-80℃保存,備用。
1.2 實驗方法
1.2.1 根系組織切片及透射電鏡觀察
觀察鈾處理后的蠶豆幼苗的生長狀況并進行拍照。選取對照組和高濃度鈾處理組(25μmol·L-1)的幼苗根系,根據Brooks等[23]的方法,制備主根和側根的石蠟切片(縱切,厚度3~4μm,番紅固綠染色法),用普通光學顯微鏡分析根系的微觀結構。選取對照組和高濃度鈾處理組的幼苗根系,將樣本用電鏡固定液處理后,經樹脂包埋-超薄切片-負染等步驟后,制備超薄切片(厚度50~60 nm),用日立120 kV HT7800透射電子顯微鏡[天美(中國)科學儀器有限公司]觀察鈾對根系超微結構的影響。
1.2.2各部分生物量及鈾含量的測定
樣品先105℃殺青15 min,然后85℃烘干至恒質量。測定各處理組(0、5、10、15、20、25μmol·L-1)蠶豆幼苗地上部和地下部的生物量。用微型植物粉碎機(上海科恒)將樣品粉碎,每個樣品取0.100 0 g置于消解罐內,加入8 mL濃硝酸和2 mL 30%過氧化氫,用微波消解儀(意大利Milestone公司)對植物樣品進行消解,采用電感耦合等離子體質譜儀Agilent 7700x ICP-MS(美國安捷倫公司)測定鈾含量。
1.2.3 葉綠素含量及光合參數的測定
葉綠素含量測定采用乙醇-丙酮(3∶1,V/V)提取法,稱取0.100 0 g植物葉片,加入5 mL色素提取液,快速研磨后浸提5 h,再用紫外分光光度計測定波長在645 nm和663 nm下的吸光值,根據相關公式計算葉綠素含量[24]。用LCpro-SD+全自動便攜式光合儀(北京澳作生態儀器有限公司)測定蠶豆幼苗的光合參數,包括:凈光合速率(A,μmol?m-2?s-1)、氣孔導度(Gs,mmol?m-2?s-1)、胞間CO2濃度(Ci,μmol·mol-1)和蒸騰速率(E,mmol?m-2?s-1),每個處理至少測定9次。
1.2.4 根系活力的測定
采用2,3,5-三苯基氯化四氮唑(TTC)比色法測定經不同濃度鈾(0、5、10、15、20、25μmol·L-1)處理后幼苗的根系活力。稱取0.500 0 g根,加入4 g·L-1TTC溶液和0.66 mol·L-1磷酸緩沖液各5 mL,在37℃下暗保溫2 h,加入1 mol·L-1硫酸2 mL終止反應,取出根系吸干表面水分后,加入乙酸乙酯充分研磨,定容至10 mL,分光光度計測定波長485 nm下的吸光值,再根據標準曲線,求得TTC還原量。
1.3 鈾脅迫下蠶豆的轉錄組分析
選取鈾處理(0μmol·L-1和25μmol·L-1)72 h后的根系樣品進行轉錄組測序,進行3次獨立的生物學重復。建庫測序及數據分析部分委托上海歐易生物醫學科技有限公司完成,將得到的Unigene.fa文件上傳到Mercator注釋網站(https://www.plabipd.de/portal/mercator-sequence-annotation)以獲得蠶豆的Mapping文件。采用MapMan軟件(Version 3.6.0RC1)對與光合作用和線粒體電子傳遞途徑相關的差異表達基因進行可視化分析[25]。差異表達基因(DEGs)的篩選條件是基因表達量差異倍數大于1.5,即 |log2FC|>1.5,且P小于0.05。
1.4 數據處理與分析
鈾的轉移系數按下列公式計算:
鈾的轉移系數(TF)=植物地上部鈾含量(mg?kg-1DW)/植物地下部鈾含量(mg?kg-1DW)式中:DW表示植物干質量。
采用SPSSv.20.0(SPSSInc.,Chicago,IL,USA)軟件對數據進行單因素方差分析和LSD多重比較。采用Origin 9.0(Origin Lab.Corporation,Northampton,MA,USA)作圖;采用Adobe Illustrator CS6進行圖像排版。
2 結果與分析
2.1 蠶豆幼苗各部分生物量、鈾含量及鈾轉移系數
如表1所示,鈾脅迫72 h后,隨著鈾處理濃度的增加,蠶豆幼苗地上部和地下部生物量呈降低的趨勢。當鈾濃度為25μmol·L-1時,與對照組比較,地上部和地下部生物量分別降低了11.63%和9.09%。當鈾濃度為5~25μmol·L-1時,地上部和地下部鈾累積量分別為0.06~0.24 mg·kg-1和766.28~5 309.82 mg·kg-1。在不同濃度的鈾脅迫下,鈾的轉移系數均小于0.01。結果表明,鈾的主要蓄積器官是根系,且難以易位至地上部。
2.2 蠶豆幼苗根系組織切片及透射電鏡觀察
如圖1A和圖1B所示,與對照組相比,U-25 μmol·L-1組的蠶豆幼苗根系出現了黑化現象。如圖1C至圖1F所示,鈾毒害明顯損傷了蠶豆根系的根冠層,表現為鈾處理組蠶豆根系的主根和側根的根冠層完全脫落。如圖1G和圖1H所示,鈾脅迫后的根系細胞壁區域出現明顯的鈾鹽沉積位點。這些結果表明,根系細胞壁是鈾的主要分布位點,過量蓄積鈾明顯損傷了根系結構。

表1鈾對蠶豆各部分生物量和鈾含量的影響Table 1 Effects of Uon the growth of Vicia faba L.and its accumulation content in plant
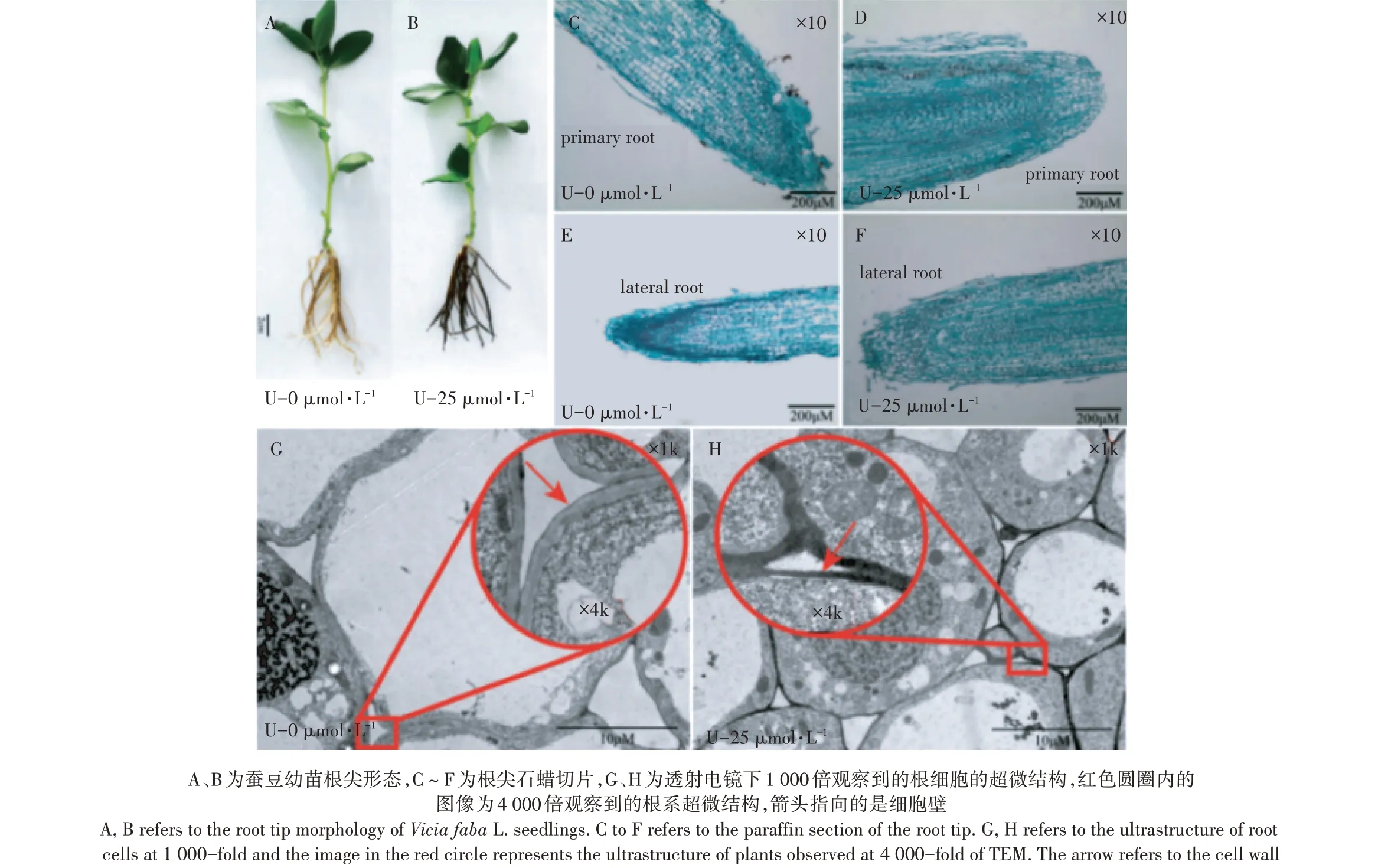
圖1鈾脅迫對蠶豆幼苗生長及微觀結構的影響Figure 1 Effects of Ustress on the growth and microstructure of Vicia faba L.seedlings
2.3 鈾脅迫下蠶豆幼苗的葉綠素含量及光合參數
如表2所示,鈾脅迫72 h后,5~25μmol·L-1組與對照組比較,葉綠素a、b和總葉綠素含量分別降低了4.32%~11.02%、3.9%~7.34%和4.21%~9.84%。隨著鈾處理濃度的增加,蠶豆幼苗光合作用參數(光合速率、氣孔導度和蒸騰速率)均顯著降低,但對葉片胞間CO2濃度的影響不明顯。當鈾濃度為25μmol·L-1時,與對照組比較,光合速率、氣孔導度和蒸騰速率分別降低了47.99%、38.66%和28.91%(圖2)。結果表明,高濃度鈾脅迫條件下,根系累積過量鈾,誘發植物光合作用異常。
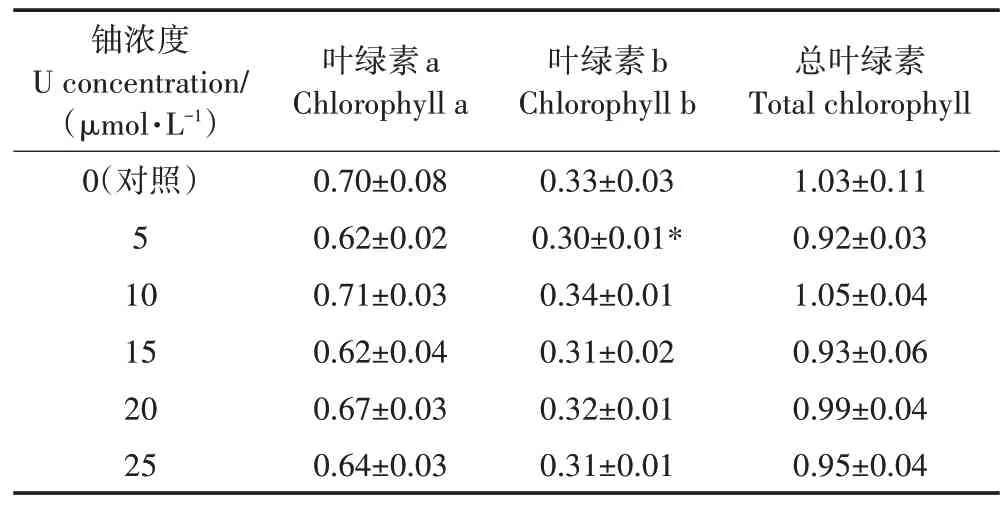
表2鈾脅迫對蠶豆幼苗葉綠素含量的影響(mg·g-1)Table 2 Effects of Ustress on chlorophyll content of Vicia faba L.(mg·g-1)
2.4 鈾脅迫下蠶豆幼苗的根系活力
如圖3所示,隨著鈾處理濃度的增加,蠶豆幼苗根系活力呈先升高后降低的趨勢。當鈾濃度為15 μmol·L-1時,根系活力達到最大值,比對照組提高了2.4倍。結果表明,鈾脅迫下蠶豆幼苗可通過誘導根系增強細胞的呼吸代謝,從而提高對鈾的耐受性。
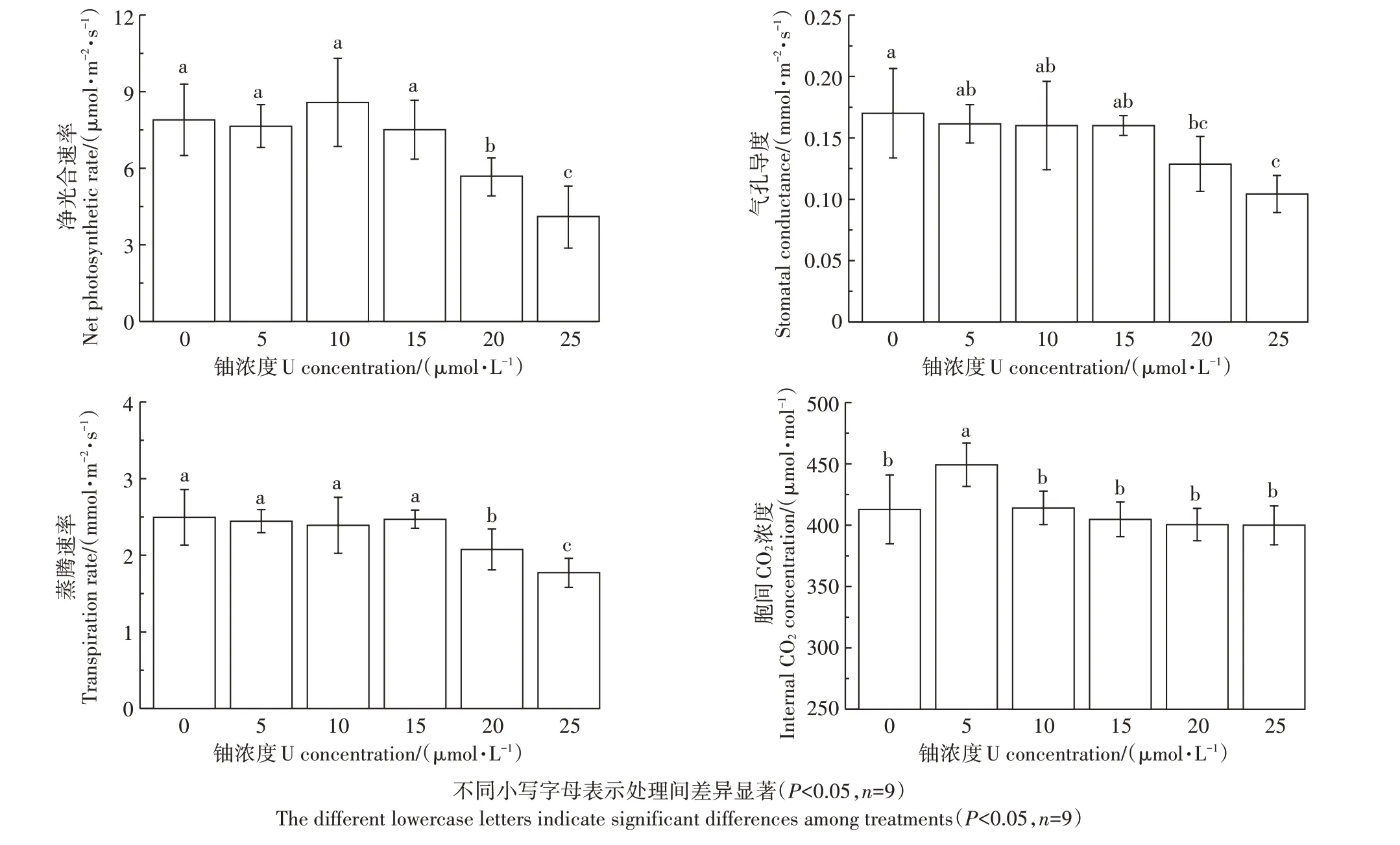
圖2鈾脅迫對蠶豆幼苗光合參數的影響Figure 2 Effects of Ustress on photosynthetic parameters of of Vicia faba L.seedlings
2.5 鈾脅迫下蠶豆幼苗光合作用相關基因的表達
RNA-seq結果顯示,鈾對光合作用的光反應途徑、卡爾文循環途徑及光呼吸途徑相關基因的表達具有明顯的抑制作用(90個DEGs;3個上調,87個下調)(圖4和表3)。例如,在光反應途徑中,高濃度鈾顯著地抑制了光反應電子傳遞鏈途徑中相關基因的表達(57個下調)(圖4A和表3)。在卡爾文循環途徑中,多種關鍵酶的基因也被顯著抑制,如transketolase gene(4.91)、aldolase gene(6.27)、triosephosphate isomerase gene(6.67)(圖4B和表3)。在光呼吸途徑中,高濃度鈾也顯著抑制了光呼吸代謝途徑相關基因的表達(圖4C和表3)。結果表明,鈾脅迫導致蠶豆幼苗光合作用效率降低,其機理是高濃度鈾損傷了光反應途徑中的電子傳遞鏈,抑制了卡爾文循環途徑和光呼吸途徑。
2.6 鈾脅迫下蠶豆幼苗線粒體電子傳遞途徑相關基因的表達
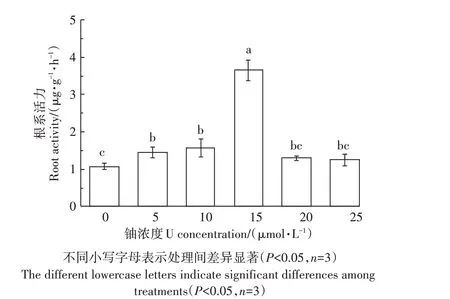
圖3鈾脅迫對蠶豆根系活力的影響Figure 3 Effects of Ustress on root activity of Vicia faba L.
如圖4D和表4所示,在高濃度鈾脅迫條件下,線粒體呼吸代謝途徑相關基因表達量明顯增加,共檢測到27個差異表達基因,包括23個顯著上調基因和4個顯著下調基因。例如,鈾誘導ATP synthesis.alternative oxidase gene、ATP synthesis.NADH-DH.complexⅠ和ATP synthesis.NADH-DH.typeⅡ.internal matrix gene的表達量分別增加了3.97、3.35倍和3.33倍(表4),表明植物根系通過加強自身的呼吸代謝,提高植物對鈾毒害作用的耐受能力,該結果與根系活力的結果一致。

表3鈾脅迫下與光合作用途徑相關的差異表達基因(前3位)Table 3 List of differentially expressed genes(TOP3)related to photosynthesispathway under Ustress
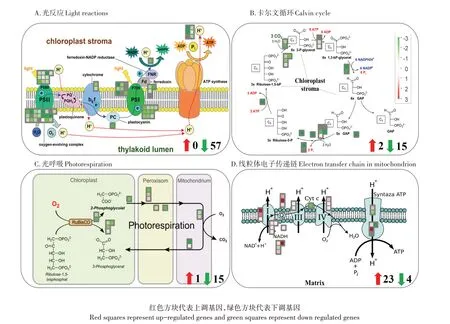
圖4鈾脅迫下與光合作用和線粒體電子傳遞鏈相關的差異表達基因Mapman可視化分析Figure 4 Mapman visualization analysis of differentially expression gene related to photosynthesis and mitochondrial electron transport under Ustress

表4鈾脅迫下與線粒體電子傳遞途徑相關的差異表達基因(前3)Table 4 List of differentially expressed genes(TOP3)related to mitochondrial electron transport pathway under Ustress
3 討論
3.1 鈾對蠶豆幼苗生長發育及根系結構的影響
植物的生物量是反映其生長狀況的重要指標[26]。在一定濃度范圍內的重金屬脅迫下,一般表現為低濃度促進植物生長和高濃度抑制植物生長[27-28]。例如,Misson等[29]的研究表明,在沒有磷的情況下,低濃度(2μmol·L-1)的鈾脅迫比對照組(0μmol·L-1)增加了植物生物量,而高濃度(50μmol·L-1和500μmol·L-1)的鈾脅迫導致植物生物量分別減少了40%和69%,其地上部分別下降25%和38.5%。本研究發現鈾脅迫下蠶豆的生物量略有下降,但差異不顯著,推測可能與鈾脅迫時間、濃度和植物種類不同有關。
已有研究表明,鈾主要在植物根系中積累,而向地上部轉移的鈾含量極低[7,30]。我們的研究結果與此類似,與對照組相比,鈾脅迫的蠶豆根系中鈾的累積量是地上部的數萬倍,且傳遞系數極低。有研究證實,由于根系細胞壁的阻滯作用[31-32],過量的重金屬累積在植物根系,以緩解重金屬對植物地上部的毒害,但會對根系造成損害。例如,鎘會誘導植物根部的邊緣細胞脫落;隨著鎘濃度的增加,根冠邊緣的細胞數量也會增加,由此產生的邊緣細胞形成黏液層,防止鎘進入根部[33]。植物根系對鈾酰離子有較高的吸附或積累作用,而鈾向莖葉的遷移受細胞壁沉降的限制[34]。與此結果類似的是,我們發現高濃度的鈾脅迫會導致植物根部的根冠脫落。與U-0μmol·L-1組相比,U-25μmol·L-1組的根尖細胞壁在透射電鏡下出現黑色沉淀物。這些結果表明,根系細胞壁阻礙了鈾向地上部的轉移,這可能是蠶豆耐受鈾脅迫的重要原因。付倩等[12]通過研究蠶豆對銫的吸收及其亞細胞分布,也得出了類似的結論。
3.2 鈾對蠶豆幼苗光合作用及其相關基因表達的影響
光合作用是在植物葉綠體內進行的儲能過程,其受葉綠素含量的影響[35]。另外,光合參數能夠直接反映光合作用的強弱。研究表明,在高濃度鈾(U-25 μmol·L-1)脅迫下,蠶豆幼苗的葉綠素含量有所下降但并不顯著,而光合參數(除胞間CO2濃度外)顯著降低。前人研究表明,重金屬脅迫下,葉綠素含量、凈光合速率和蒸騰速率降低,而胞間CO2濃度升高[36-37]。我們推測,高濃度鈾脅迫對蠶豆的光合效率有明顯的抑制作用,但在短時間(72 h)內葉綠素沒有被破壞。植物光合作用的變化受到相關基因的調控。Houot等[38]的研究表明,在鎘脅迫下,大量參與光合作用的基因(通常提供ATP和NADPH)表達下調,以適應鎘脅迫。我們的研究結果也表明,高濃度的鈾脅迫顯著抑制了植物光合作用中光反應途徑、卡爾文循環途徑和光呼吸途徑相關基因的表達。
3.3 鈾對蠶豆幼苗根系活力和呼吸代謝相關基因表達的影響
根系活力可以反映根系吸收水分和養分的能力,甚至與重金屬的遷移有關[28,39-40]。在本研究中,我們發現鈾濃度在0~25μmol·L-1范圍內,蠶豆幼苗的根系活力呈現先升高后降低的趨勢。我們認為,在低濃度鈾脅迫下植物能夠通過增強根系活力來維持自身正常的生理代謝,而當鈾濃度過高時會對植物結構造成損傷,從而導致根系活力下降。田小霞等[40]在研究鎘對馬藺根系活力的影響時也得到了類似的結果。逆境脅迫下,線粒體中與電子傳遞鏈相關基因的表達受到影響,其負責控制植物體內ATP的水解和能量釋放的過程,釋放的能量主要用于抵抗脅迫[41-42]。Irani等[43]的研究發現,在逆境脅迫下,擬南芥莖葉中與線粒體電子傳遞相關基因的表達有所增強,以提高植物的抗逆性。與此類似,我們的研究結果表明高濃度的鈾脅迫誘導了蠶豆幼苗根系的線粒體中呼吸電子傳遞途徑相關基因的高表達。
4 結論
(1)在鈾脅迫下,蠶豆根系能阻止大部分鈾向地上部轉移,從而維持植株地上部的正常生長。然而,蠶豆根系中鈾的過量累積造成了根系損傷,影響植物的生理代謝活動。
(2)高濃度的鈾脅迫降低了蠶豆幼苗的光合效率,抑制了光合作用過程中光反應途徑、卡爾文循環途徑和光呼吸途徑相關基因的表達。在一定的鈾濃度范圍內,蠶豆幼苗的根系活力呈現出先升高后降低的趨勢,高濃度的鈾脅迫誘導了根系線粒體中呼吸電子傳遞途徑相關基因的高表達。
(3)在鈾脅迫下,蠶豆幼苗的光合作用受到影響,但其根系的呼吸代謝有所增強,從而適應鈾脅迫環境。蠶豆對鈾具有一定的耐受性,但由于鈾主要累積在其根系,因此不能作為鈾的富集植物。
猜你喜歡
中學生數理化·七年級數學人教版(2019年10期)2019-11-25 07:33:58
中學生數理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
中國衛生(2016年3期)2016-11-12 13:23:26
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
