β-DG過表達對結腸癌細胞系SW620細胞凋亡的影響
2020-10-13 14:32:36潘林鑫
安徽醫科大學學報 2020年10期
錢 成,徐 濤,潘林鑫
結腸癌是常見的發生于結腸部位的消化道惡性腫瘤,可向肝、肺、腦、骨等臟器轉移,晚期可出現貧血、消瘦、惡病質等表現,嚴重者危及生命。除了對結腸癌的臨床研究外,目前對其細胞生物學研究也越來越深刻[1-2]。HCT116、SW620等細胞系是從結腸癌患者中分離建立的惡性細胞,廣泛應用于結腸癌的細胞學研究。人類的肌營養不良蛋白 (dystroglycan,DG)是由位于染色體3p21上的DAG1基因編碼的含有895個氨基酸殘基的跨膜蛋白,在體內翻譯修飾后發生多肽鍵斷裂,最終水解成α-DG和β-DG 2個具有獨立功能的蛋白。α-DG是位于細胞膜上的高度糖基化蛋白,并通過與多種細胞外基質(extracellular matrix,ECM)蛋白結合進行信號轉導。而β-DG作為跨膜蛋白,其胞內區直接與抗肌萎縮蛋白(dystrophin,Dys)結合,從結構上分析,β-DG主要由2個結構域組成,即N端(654~750 aa)和C端(775~895 aa),它們通過跨膜結構域(transmembrane domain,TM)相連[3]。β-DG的N端與α-DG的C端通過非共價鍵的方式交聯,其C端的酪氨酸序列能夠結合胞內的信號分子,并通過酪氨酸的磷酸化參與激活多條信號轉導途徑,發揮著調節細胞的分化、增殖、凋亡以及改變細胞的極性等生物功能,在癌癥、遺傳性疾病等疾病的發生發展中扮演著重要角色[4-5]。該研究首先檢測四株常見的人結腸癌細胞系中β-DG的內源表達情況,以確定β-DG低表達細胞系SW620,然后構建pcDGFP-β-DG的真核表達質粒,并過表達于SW620細胞中,研究β-DG對其細胞凋亡及相關蛋白表達的影響。
1 材料與方法
1.1 質粒、菌株與細胞pcDNA3.1-FLAG-DAG1和pcDGFP真核表達載體質粒由安徽醫科大學生物系范禮斌教授實驗室惠贈,DH5α感受態菌株HEK 293T和SW620等結腸癌細胞株由校實驗中心實驗室保存。
1.2 主要試劑與儀器DNA聚合酶購自日本TaKaRa公司;BamH I、Not I內切酶以及T4連接酶購自上海生工生物公司;膠回收、質粒小量抽提試劑盒購自美國Axygen公司;細胞裂解液、PMSF蛋白酶抑制劑、一抗稀釋液購自上海碧云天公司;GFP抗體、β-DG抗體購自美國Santa Cruz公司;DMEM培養基購自澳洲HyClone公司;胎牛血清購自美國CLARK公司;脂質體轉染試劑盒、Opti-MEM低血清培養基購自美國Invitrogen公司。AL600RGB凝膠成像系統購自美國GE公司;Axio Observer 3倒置熒光顯微鏡購自德國卡爾蔡司公司;MoFlo XDP超速流式細胞分選儀、CytoFlex流式細胞分析儀購自美國Beckman公司。
1.3 方法
1.3.1質粒構建[6]設計并合成引物序列:F:CC GGAATTCCGGGCCACCATGTCCATCGTGGTGGAA;R:AAGGAAAAAAGCGGCCGCAAAAGGAAAACAGG TGGGACCATAGGGA,以pcDNA3.1-FLAG-DAG1真核表達質粒為模板,PCR擴增出β-DG的CDS序列,將產物進行瓊脂糖凝膠電泳,并利用膠回收試劑盒進行回收,得到其PCR純化產物,利用限制性內切酶EcoR I和Not I對β-DG的PCR純化產物和pcDGFP空載體分別進行雙酶切,用T4 DNA連接酶將二者的酶切產物進行連接,并將連接產物轉化于DH5α感受態細胞中,37 ℃培養至單克隆生成(約12 h),分別挑取數個單克隆進行擴大培養(約12 h),并抽提菌液中的質粒進行雙酶切鑒定(EcoR I和Not I),選擇鑒定正確的重組質粒送公司進行測序。
1.3.2細胞培養 37 ℃水浴復蘇凍存于液氮中的各細胞系,并用完全培養基(90% DMEM高糖+10% FBS+青鏈霉素)進行培養,培養環境為37 ℃、5% CO2的恒溫培養箱。定時觀察細胞,及時更換完全培養基,待細胞生長至90%以上密度時即可進行細胞傳代,將適量細胞重新接種至新的培養皿或培養瓶中,用于后續實驗。
1.3.3質粒轉染 本研究采用的轉染方法為脂質體轉染法,步驟如下(以六孔板為例):分別將適量pcDGFP-β-DG(2~3 μg)或pcDGFP和lip2000(4~6 μl)加到適量Opti-MEM轉染液中(約200 μl),并輕輕充分混勻,室溫靜置5 min后將兩者輕輕充分混勻,室溫靜置20 min后,將溶液加入待轉染的細胞中,培養4~6 h后更換為完全培養基,繼續培養48 h,用于后續蛋白表達實驗。
1.3.4Western blot檢測 轉染pcDGFP-β-DG約48 h后,分選并收集陽性細胞進行裂解(冰上或4 ℃),30 min后收集細胞裂解液進行高速冷凍離心(14 000 r/min,10 min)并收集上清液,蛋白定量后取出適量上清液,加入等量2×SDS上樣緩沖液充分混合,沸水浴5 min,冰浴5 min后取適量樣本進行 SDS-PAGE電泳(80 V約0.5 h,100 V約1.5 h),200 mA恒流電轉1 h,取出PVDF膜,用含5%脫脂奶粉的TBST溶液封閉1~2 h,TBST溶液洗膜后加入GFP一抗 (1 ∶500)進行孵育(4 ℃過夜或室溫2 h),TBST洗膜后加入HRP標記的山羊抗兔IgG(1 ∶5 000)進行孵育(室溫1 h),TBST溶液洗膜后用ECL試劑盒進行顯色,在蛋白成像儀上進行拍攝。
1.3.5免疫熒光觀察 轉染pcDGFP-β-DG約24 h后,取出SW620細胞爬片,用預冷的PBS溶液清洗3次后進行固定(-20 ℃預冷甲醇2 min+70%乙醇5 min),由于GFP能夠自發綠色熒光,因此無需進行一抗和二抗孵育,固定后用預冷的PBS溶液清洗3次后進行細胞核染色(0.15 g/L DAPI 2 min),棄DAPI溶液后用預冷的PBS溶液清洗3次后,用熒光封片膠將蓋玻片封于載玻片上。4 ℃保存過夜后用熒光顯微鏡進行觀察。
1.3.6細胞凋亡 將轉染了pcDGFP-β-DG(實驗組)和pcDGFP空載體(對照組),以及未轉染(空白組)的SW620,用不含EDTA的胰酶消化后低速收集(1 000 r/min, 3 min),用預冷的PBS洗滌細胞2次后,加入400 μl×Binding Buffer重懸細胞(濃度大約為1×106/ml),加入5 μl Annexin V-FITC,輕輕混勻后于2~8 ℃避光條件下反應15 min,然后加入10 μl PI輕輕混勻后于2~8 ℃避光條件下反應5 min,混勻后放置于冰上,樣品在1 h內用流式細胞儀檢測凋亡情況。凋亡圖分為4個象限,左下象限為細胞存活率(Q1-LL)、左上象限為細胞死亡率(Q1-UL)、右上象限為細胞晚期凋亡率(Q1-UR)、右下象限為細胞早期凋亡率(Q1-LR),本次實驗凋亡率為Q1-UL、Q1-UR和Q1-LR 3個象限數值之和。
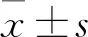
2 結果
2.1 結腸癌細胞系中β-DG的表達情況培養HT29、HCT116、SW480、SW620等結腸癌細胞系以及HEK293T腎上皮細胞系,并收集適量以上5種細胞進行裂解,提取總蛋白,定量分析后進行Western blot實驗,當用β-DG抗體進行免疫印跡時,結果發現5種細胞蛋白樣品在43 ku處均出現蛋白條帶,大小基本符合β-DG的分子量。但與正常上皮細胞相比,各結腸癌細胞系中β-DG的表達量都有不同程度的減少,其中SW620細胞的β-DG表達量最少,適合用于后續β-DG的過表達實驗,見圖1。

圖1 結腸癌細胞系中β-DG的表達情況

圖2 pcDGFP-β-DG重組質粒的酶切鑒定與測序結果
2.2 β-DG真核表達載體的構建通過PCR擴增、雙酶切、酶連、質粒轉化與抽提后擬得到pcDGFP-β-DG重組質粒,利用EcoR I和Not I進行雙酶切鑒定,瓊脂糖電泳結果如圖2A所示,重組質粒被切出兩條亮帶,通過DNA Marker、PCR產物和空載體的橫向比較,發現一條亮帶大小約為6 100 bp,與pcDGFP空載體的大小基本一致,另一條亮帶大小約為720 bp, 與β-DG的片段大小基本一致。將此重組質粒送生物公司進行測序,并將測序結果與β-DG的CDS序列(729 bp)進行比對,堿基序列完全一致,表明pcDGFP-β-DG重組質粒構建成功,見圖2B。
2.3 β-DG在SW620細胞中的過表達和熒光定位情況將pcDGFP-β-DG瞬時轉染至SW620細胞中,收集、裂解細胞后進行Western blot檢測,當用GFP抗體進行免疫印跡時,結果顯示空載體對照組、β-DG過表達組分別在25 ku和70 ku左右的位置出現條帶,即為GFP和GFP-β-DG(27+43 ku)融合蛋白的蛋白表達,而空白對照組未見任何條帶,表明pcDGFP-β-DG重組質粒能夠在SW620細胞中過表達,見圖3。另外,制作SW620細胞爬片,過表達GFP-β-DG后進行免疫熒光制片,并在熒光顯微鏡下進行觀察,結果顯示GFP-β-DG自發綠色熒光,主要定位在細胞質中,細胞核內未見顯著表達,且在細胞質中靠近核膜一側的定位更加明顯,見圖3。
2.4 β-DG過表達對SW620細胞凋亡的影響分別將pcDGFP-β-DG和pcDGFP(作為對照)瞬時轉染至SW620細胞中,約24 h后收集細胞,分別進行Annexin V-FITC/PI雙染,然后利用流式細胞術檢測凋亡情況,共進行3次重復實驗,結果如表1所示,統計分析后發現,空白組(未轉染)和對照組(轉染pcDGFP空載體)細胞凋亡率的差異無統計學意義(P>0.05),與對照組相比,實驗組(轉染pcDGFP-β-DG)細胞的凋亡率升高(P<0.05),差異有統計學意義,見圖4,表明過表達β-DG能夠有效促進SW620結腸癌細胞凋亡。

圖3 β-DG在SW620細胞中的過表達和熒光定位圖 ×630

圖4 β-DG過表達后SW620細胞的凋亡圖
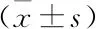
表1 β-DG過表達對SW620細胞凋亡的影響
2.5 β-DG過表達對細胞凋亡相關蛋白的影響分別將pcDGFP-β-DG和pcDGFP(作為對照)瞬時轉染至SW620細胞中,約24 h后收集細胞,提取總蛋白后進行Western blot檢測,共進行3次重復實驗,當用PARP抗體進行免疫印跡時,結果顯示空白組、空載體對照組、β-DG過表達組均在113 ku左右的位置出現條帶,即為完整的PARP蛋白,相對而言,β-DG過表達組的PARP蛋白含量有所降低,且在約90 ku的位置還出現了另一條帶,即為PARP的裂解片斷,見圖5。表明過表達β-DG能夠有效促進細胞凋亡蛋白PARP裂解,即促進SW620結腸癌細胞凋亡。

圖5 β-DG過表達對細胞凋亡蛋白PARP的影響
3 討論
DG是一種跨膜的糖蛋白,β-DG作為其水解衍生物,是組成細胞表面的重要成分,廣泛分布于脊椎動物的骨骼肌和其他組織中[7]。β-DG的N端存在著潛在的N-連接糖基化位點,在其C末端存在一個PPXY序列,該序列又與一個含有兩個高度保守色氨酸殘基的WW結構域相結合,構成了蛋白質相互作用的一種模式,此結合的穩定性取決于抗肌萎縮蛋白的EF-手形結構域。此外,β-DG的胞質尾區能夠與包括細胞外調節蛋白激酶(extracellular regulated protein kinases,ERK)、絲裂原激活的蛋白激酶激酶(mitogen-activated protein kinase kinase,MEK)、生長因子受體結合蛋白2(growth factor receptor-bound protein 2,GRB2)等在內的眾多信號分子充分結合,進而起到穩定神經肌肉接頭,傳遞細胞信號轉導的功能。DG的異常表達是癌癥發生的一個重要特征,與腫瘤的發展和轉移密切相關[8],這種異常除了與α-DG糖基化水平改變有關外,還與β-DG的異常表達有關。在人類的乳腺癌、前列腺癌、宮頸癌、舌鱗狀細胞癌中都發現了β-DG的異常[9-11],這種異常主要表現在其非正常裂解導致的表達量下降。本研究首先利用免疫印跡技術檢測了HT29、HCT116、SW480、SW620等常見的結腸癌細胞系中內源性β-DG的表達情況,結果發現各癌細胞系中正常分子量β-DG(43 ku)的表達量均有不同程度的下降,其中SW620下降的最為明顯,另外,這些結腸癌細胞系在約31 ku的位置均出現了蛋白條帶,這可能是正常β-DG蛋白水解后的胞外結構域,它的產生是由于膜相關基質金屬蛋白酶的活化,破壞了DG復合物的結構,最終導致癌癥的發生。
本研究首先確定了β-DG在各結腸癌細胞系中呈現低表達狀態,接下來選擇了低表達程度最高的SW620細胞進行β-DG的過表達研究。利用基因重組技術構建了pcDGFP-β-DG真核表達質粒,并將其轉染至SW620細胞中,利用Western blot技術檢測了其表達情況,結果發現β-DG能夠成功過表達。同時還利用了免疫熒光技術對β-DG在SW620中的細胞定位進行了檢測,結果發現GFP-β-DG蛋白主要定位在細胞質中,而且在細胞質中靠近核膜的一周表達量更多,此結果與之前研究的FLAG-β-DG在COS7細胞中的定位結果基本一致[12]。而且根據位置推測,β-DG蛋白定位可能與高爾基體或內質網有著密切的聯系,所以在接下來的研究中,可以選擇高爾基體或內質網的標志染料麥胚凝集素或刀豆球蛋白A與β-DG進行共定位研究,進一步確定β-DG蛋白的具體亞定位。
本研究旨在探索β-DG對結腸癌細胞系細胞凋亡的影響,細胞凋亡是細胞的一種基本生物學現象,是由基因控制的細胞自主有序的死亡,在生物體的進化、內環境的穩定以及組織系統的發育中起著重要的作用。而細胞凋亡的抑制或減弱與癌癥的發生發展有著密切的聯系,許多化療藥物發揮作用的機制就是誘導癌細胞的凋亡[13]。因此本研究在確定結腸癌細胞系和β-DG的過表達效果后,便利用流式細胞術檢測過表達β-DG對SW620細胞凋亡的影響,結果發現β-DG過表達后能夠提高SW620細胞的凋亡率,表明β-DG具有促凋亡功能。為了進一步確定這個結果,又檢測了過表達β-DG后SW620細胞中二磷酸腺苷核糖多聚酶(PARP)蛋白水平的變化,PARP為一種依賴鋅離子(Zn2+)的真核DNA結合蛋白,可特異性地識別DNA斷裂末端并結合,是一種重要的DNA修復酶,在細胞凋亡發生的早期, PARP是含半胱氨酸的天冬氨酸蛋白水解酶家族(如caspase 3和caspase 7)的作用底物,后者可使PARP的DNA 修復功能喪失,并將113 ku的PARP水解為89 ku的片斷。因此PARP可以作為細胞凋亡早期的標志物,許多研究便是通過Western blot檢測PARP的完整蛋白(113 ku)及其裂解片斷(89 ku)的表達量來觀察細胞凋亡效果[14]。本研究檢測發現β-DG過表達后PARP的完整蛋白(113 ku)含量下降,且裂解片斷(89 ku)的表達量升高,同樣表明過表達β-DG具有促凋亡功能。
綜上所述,β-DG在結腸癌細胞系中異常表達,主要表現在其異常的水解斷裂。過表達β-DG能夠顯著促進SW620細胞凋亡及相關凋亡蛋白的表達,為進一步了解β-DG功能,及其與結腸癌之間的關系提供了細胞學基礎。
猜你喜歡
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
西南國防醫藥(2016年7期)2016-12-01 06:01:15
海峽科技與產業(2016年3期)2016-05-17 04:32:12
中國衛生標準管理(2015年1期)2016-01-14 03:41:26
河南醫學研究(2014年3期)2014-02-27 14:51:48
沈陽醫學院學報(2014年1期)2014-02-16 06:19:24
