競賽表現的力量突破與系統訓練的“總功”約束(下)
2020-10-21 06:02:10李端英晟3王曉軍4
中國體育教練員 2020年3期
關鍵詞:生物
李 捷, 李端英, 裘 晟3, 王曉軍4, 李 稚
(1.廣東省體育局,廣東 廣州 510108; 2.廣州體育學院,廣東 廣州 510500;3.廣東省體育科學研究所,廣東 廣州 510663; 4.廣東省體育局黃村體育訓練中心,廣東 廣州 510663;5.廣東工業大學,廣東 廣州 510006)
前文我們以賽艇項目為例,結合部分“119”項目,對當前我國競速類項目訓練的常見問題進行了實證分析,運用系統訓練理論的總功約束原理,解析這些訓練現象的發生原因。本文繼續對此進行討論。
1 競速項目專項定義邏輯混亂
在體育運動項目中,無論依據何種分類方法,形式邏輯的排中律告訴我們:在同一個思維過程中,2個相互矛盾的思想不能同假,必有一真。即“要么A要么非A”,不能對不能同假的命題(矛盾關系、反對關系)同時加以否定。根據排中律,若競賽項目規則規定是競速,這些項目就是比速度的項目,速度項目的核心是限定的距離內時間越短越好,表達為距離與動作鏈時間的比值。專項發展的邏輯是縮短固定距離內的動作鏈時間,客觀上表現為專項距離內單位時間內動作鏈秩序的輸出功率越大越好。根據質能方程,其訓練核心是:在有限的時間內實現競賽距離條件下的動作鏈時間能量容量的最大化。
很多教材或文獻對競速類項目都存在定義模糊的問題,如競速項目是“周期”運動、“速度耐力”運動、“速度有氧耐力”運動等。進行屬性范疇的定義延伸,不僅違反“排中律”,也忽略“速度”的生物本質而誤導訓練。從系統訓練的角度看,速度是限定距離內時間限制的動作鏈能量秩序表達,形成目標化系統與能量鏈極限突破的訓練程序遞階過程始終是競速項目訓練的根本。
2 訓練突破核心問題不清晰
目前在我國,無論是訓練管理單位下達任務,還是教練員申報參賽任務,都是報名次而不報具體成績指標。由于名次只是競賽結果,其本身并不是目標成績標準,內在具有對競賽表現狀態的邏輯定態約束性。假若金牌設定與具體目標成績不掛鉤,沒有目標成績標準,運動員與競賽目標差距的焦點問題就不能具體化,訓練過程的系統標準定位結構就難以實現,訓練內容的指向性及其組織程序就缺乏目標標準的約束性。運動競賽的本質是物理量值(時間、分數)在相應系統的生物能量表達,確定的競賽成績就是確定動作鏈的生物能量秩序表現。
對訓練過程組織來說,沒有競賽目標標準的約束就沒有目標時間限制的物理總功約束,沒有目標時間限制的總功約束就沒有目標動作鏈總功率時序約束,沒有目標動作鏈總功率時序約束就沒有目標系統力量協同標準的要求與生物能量在動作鏈的時序分配依據。因為成績表現是運動員專項生物系統的節點狀態,所以,目標定態形成是競賽節點穩定狀態表現的生物基礎。訓練的核心在于按照目標成績(比如賽艇2 000 m,目標成績設計為5 min 41.9 s,訓練組織過程就應以5 min 41.9 s的定態節奏作為主結構)進行遞階發展倒計時訓練。訓練過程的主結構內容始終依據競技目標(成績)表現標準約束的能量生物結構時間表達系統構建,而非泛泛的“兩極化”,或者所謂“有氧訓練”、元素“加和”訓練、“沖酸”適應訓練等。用目標成績量值約束系統的所有相關訓練因素,建立目標動力鏈的能量秩序,提高目標時間限制條件下的生物能量轉換效率成為訓練的核心。實現目標動作鏈的能量定態,才能得到目標系統在競賽節點的穩定表現。具體訓練模式如跳水、體操、乒乓球的目標動作鏈重復訓練,賽艇的能量秩序訓練等,都由能量與功率輸出的生物規律決定。
賽艇、皮劃艇、自行車、游泳、田徑徑賽等競速項目,都遵循目標時間限制條件下的動作鏈生物能量秩序。能量總功率=S×F/TTB,其中,S指動作力量,F指動作頻率,TTB(Target time box)指目標時間限制,3個因素的互動關系決定專項負荷(能量總功率)。其中,動作頻率(F)并不是越快越好,而是具有項目約束與神經控制最優特點,目標時間限制(TTB)是一個定值,唯一具有無限發展邏輯空間的就是力量(S)。所以,對競速項目來說,力量與專項力量具有相對無限的發展要求與空間。以賽艇為例,在目標時間限制條件下,處理好目標劃頻與每槳力量功率在動力鏈的生物能量輸出分配最優秩序的關系是訓練突破的核心問題。這也適用于自行車、游泳等競速項目。
3 速度訓練的習慣化效應與目標動作鏈功率秩序不一致
訓練過程導致的肌肉習慣性活動(熵序)會形成基于細胞分子網絡效應的神經行為路徑,當形成習慣化記憶后,肌肉表達為對習慣速度條件下做功效率的提高與能量節省的狀態,并在意識的主導下與習慣性環境關聯成習慣性速度行為控制。但是,習慣性速度定位實際上是一把“雙刃劍”,速度習慣化就是能量秩序標準化,能量秩序標準化意味著速度能量秩序各種參數的定態控制關系。如果訓練中設定的習慣化速度是競賽目標速度,對于競賽來說沒有問題;但如果訓練經驗的習慣化速度低于競賽目標速度,就不能在競賽時達到目標速度。在競技比賽中,由于訓練導致的習慣性速度達不到競賽目標速度的情況非常常見。如果賽中強制改變習慣速度,會打破運動員能量代謝鏈的平衡,降低運動表現水平。競走的“兔子”戰術,就是通過干擾對手習慣化速度進而導致其代謝環境紊亂、降低運動表現的例子。所以,長距離項目(劃、游、騎行、跑)的任何訓練方式(距離、間歇、組數、時間限制)作為訓練手段本身都沒有問題,這些手段有沒有進行目標速度的限制才是關鍵。依據目標標準的速度距離分段多組訓練及間歇控制訓練程序,是與能量代謝量化相對應的目標速度習慣化控制的有效手段。在總功與時間約定的條件下,輸出功率的參數關系(頻率與力量)實際上是可調的。它的訓練學意義在于,在目標時間限制基礎上,將動作頻率取最佳值(不是最大值)后,加強每劃(踏)力量及優化連續力量最大化輸出的時間秩序,然后按照訓練過程的定態時間重復,在長時間的訓練過程中形成程序化目標速度的習慣化生物效應,確保競賽時達到目標速度。
4 目標能量時間秩序定態程序化組織與大周期分期訓練相矛盾
大周期訓練組織仍然是當前影響訓練效率的主要因素。確定了正確的訓練手段,并不代表訓練組織過程正確。現實中,大周期“兩極化”訓練階段內容劃分,導致目標訓練過程的多狀態結構與反向變化的能量秩序排列。從系統養成的角度看,穩定的系統狀態效應經過長年的經驗積累才能在競賽中得以表現。實踐中“兩極法”或者目的性板塊(力量訓練8~12周)訓練,都是通過提高能量代謝強度標準來發展相應的競賽狀態水平的。有些教練員在競賽節點之間(只有20天時間)也進行基本、賽前與競賽階段劃分,而不用競賽目標系統去設計穩定的生物節奏訓練負荷;無論年度競賽有幾個節點,在已經是多內容狀態訓練條件下,始終堅持貌似合理的強度、運動量反向安排;而且,訓練標準不高也要進行賽前減量等:這些都有悖于目標速度成績(動作鏈)對應的能量秩序必須經過穩定才能得以表現的系統生物學原理,人為擾動目標狀態與能量過程的自組織定位,最終在訓練組織過程中形成人為的不穩定能量生物狀態定位。因此,進行目標定態訓練或是進行大周期的“兩極法”訓練,是訓練組織原則正確與否的重要問題。在訓練組織的認識上不走出大周期分期訓練觀念,就不可能提高運動成績,多數“119”項目的奧運成績長期表現不佳就是明顯的例證。
對國際主流訓練原則最新發展認識不足。比如在訓練原則的認識上,國外訓練書籍或文獻并不存在通用的原理或手段,百花齊放的學術認識在互動碰撞中快速發展。美國的OPT訓練講究急性應激訓練;2019年12月,德國維爾茨堡大學的Billy Sperlich教授在北京體育大學的講座中提到“Principles to trigger adaptation”與“Principles to maintain adaptation”,即“觸發性適應原則”與“保持性適應原則”。在觸發性適應中,他強調特異性干涉與應激恢復比率的概念,而在保持性適應中則增加周期性訓練的強度分配比、板塊與長時程訓練設置等。這些訓練原則都基于一定的生物原理,不但豐富、發展了傳統訓練原則,也反映了德國、瑞典、美國、澳大利亞等國對訓練原則的思維與認識不斷創新。對系統訓練原理認識不足與對國際相關研究最新動態缺乏了解,是我國競技訓練理論研究人員、教練員及科研人員的“軟肋”。
5 能量輸出與氧耗生物關系認識倒置
前文依據愛因斯坦的質能方程,對競技訓練中能量與目標系統質量之間的關系進行多次討論,確定標準的質能關系實際上就是確定標準的質量與能量關系。根據化學反應的定比定律,能量確定標志著氧耗總量的確定。在人體運動中,能量消耗與氧耗是物質統一體的2個方面,相伴而生,不可分割。肌肉運動的物理功是細胞生物能量消耗的根源與鏡面反映,能量代謝導致氧呼吸鏈隨動以保持能量循環輸出,能量的代謝底物變化則是驅動機體內環境包括細胞內環境系統再平衡的根源。做功是主動過程,氧代謝鏈則是隨動過程,氧耗成為人體能量(熱量)輸出對應的生物標度,能量(功率)輸出秩序是決定氧耗與能量代謝體系動態關系的本源。
從生物原理上,安靜狀態下除正常生命代謝外,其他能量(熱量)/氧耗是不存在的,呼吸潮氣的基線就是正常能量的氧耗標準(即使是巴斯德效應也符合質能原理),所以,無論如何都是能量消耗引導氧呼吸鏈隨動以保持內環境穩態,而不是氧耗曲線引導能量輸出。時間限制與能量輸出容量的關系才是氧耗曲線幅度變化的根本原因。而且,限定時間的能量秩序的發生及與之相應的能量過程氧耗曲線涵蓋了所有與能量輸出相關系統活動的代謝底物氧化還原與內環境穩態的自組織再平衡。有氧氧化是保持系統能量反應平衡關系的動態自組織過程,涉及全身各功能網絡系統的協同活動,所以,時序能量效應經驗才是攝氧水平自組織動態調適的根源,所謂“能量連續統一體”[1]的原因也在于此。而用氧耗百分比評估訓練效果,很容易忽略與倒置競賽目標實現過程的能量與氧耗動態生物關系,不符合競賽目標是通過目標標準能量狀態定位進行訓練的實質。“以賽帶練或者系統訓練”與“有氧訓練”2種不同的思維導致訓練認識與手段不同。長期進行有氧訓練的“119”大部分項目的成績,可能和能量輸出與氧耗認識關系的倒置有直接關系。
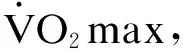
無論氧耗還是攝氧說法的根源都在于能量消耗,而氧耗導致的穩態調適使生命環境恢復正常是正常的機體自組織生命活動。因此,把最大攝氧作為主動過程是不正確的,誤導訓練設計,進而違反競技生物訓練的能量特點與競技目標表現系統形成的客觀規律。
大學生與專業運動員最大攝氧量水平差異的原因在于,產生能量的細胞數量及能量體系代謝效率不同。因為運動員的氧呼吸鏈體系、肌肉能量容積、細胞數量與能量代謝效率遠高于普通大學生,所以,影響最大攝氧量的本質是個體對應的生物能量容量效率。在生物機制上,只有通過動態發展的力量(功率)需求調適訓練,不斷增加產生能量的細胞數量、提升能量池氧呼吸鏈體系代謝效率,最大攝氧量才會發生變化。由于攝氧量決定于對應的能量消耗量,非目標標準約束的強度分級訓練與不加能量/時間條件約束的有氧訓練,均不能提高最大攝氧量。肌肉肥大訓練、力量訓練及與之相關的競賽目標力量功率秩序訓練之所以非常重要,是因為它們是能量發生的物質基礎與氧耗的限制源。在目標總功概念的前提下,擴大能量容量,通過目標能量程序化力量訓練增大肌細胞數量與體積,根據生物極限不斷提升氧呼吸效率與單位時間內能量輸出總量,提高能量利用效率,才是最大攝氧量提高的具體訓練路徑。
在徑賽項目中,一般的最大攝氧量負荷訓練其實是氧供應狀態下的能量消耗練習,與競速項目提高速度極限/能量容量的訓練生物邏輯相背離,偏離了實戰訓練的基本生物原理。在實際訓練中,形成目標速度標準限制的習慣性氧呼吸狀態才是競賽所需要的。國家體育總局近年進行的體能大比武、奧運選拔體能準入標準[5]等,對促進運動員的力量協同水平,間接促進氧呼吸效率改善,提高最大攝氧量等都具有積極意義。美國OPT優化訓練的6級遞增訓練模式,即基礎調適—增肌訓練—力量訓練—最大力量訓練—功率訓練—最大功率訓練及其優化,準確指出了通過增加力量功率促進專項提高的生物學路徑。所以,慣用的最大攝氧量概念只是運動能量輸出狀態節點的生理評估指標。最大攝氧量是一個與能量動員程度及范圍相關而有一定幅度變化的相對值。因此,對最大攝氧量概念的正確理解非常重要,不能倒置耗氧、攝氧的生物關系。
6 無氧訓練“線粒體萎縮”與競賽目標功率秩序系統訓練原理的邏輯沖突
無氧供能時間的延長,或者氧極限狀態下的能量循環保持,是競速項目的永遠追求。無氧能量代謝意味著能量輸出速率對細胞氧呼吸空間與時間的條件性調適,氧極限狀態下的穩定能量循環保持是長距離競速項目運動員保持高速的生物機制。前者為機體提供了時間限制條件下的最大功率秩序表達,后者則是時間容量內的氧能量最大化,能量調適的無氧/氧能量最大化及其細胞氧呼吸關聯網絡系統的進化調適訓練,是決定競速項目成績的核心內容。但是,在多年訓練實踐中,對這一生物原理的認識存在很多不同觀點。
“無氧訓練導致線粒體萎縮”,所以只能進行有氧訓練?曾有德國學者根據實驗結果,報道了無氧訓練環境會導致細胞內線粒體萎縮的現象,并建議賽艇運動員線粒體的改變只能通過低強度的長劃來實現。這是因為隨著強度的增加,對線粒體的影響會加大,許多線粒體會變性壞死。因此,賽艇絕大部分耐力訓練應在最大攝氧量的55%~75%。這是一個在國內船艇教練員中影響頗大的說法。以下對此進行分析。
首先,在“無氧”概念的定義上,需要從生理學角度與訓練實際情況進行區分。在生理學上,無氧是指細胞呼吸環境中沒有氧分子存在的情況;而在實際訓練狀態下,由于細胞自身的保護作用與能力最大化自組織適應網絡調適的緣故,具有傷害性的“無氧”“缺血”情況極難發生。在極限訓練中,經常遇到運動員“練不上去”的現象,盡管運動員已拼盡全力,實際輸出功率水平卻在下降。這是機體能量代謝過程自我反饋限制的自然現象,并非是運動員意志力不夠堅強。依據應激理論,傷害的發生要經過“警覺、抵抗、衰竭”3個階段,這種應激環境由于細胞效應的時域限制,通過課次訓練活動很難出現。比如國外訓練中常用的“應激恢復比率原則”,也不會出現絕對的細胞內無氧現象。
其次,線粒體萎縮的實驗多數是動物實驗,且多為急性實驗,萎縮的原因一般解釋為過量氧應激條件下活性氧(ROS)所致的細胞器損害[6]。除動物實驗外,未檢索到人體無氧訓練條件下長期跟蹤研究的文獻。當然,我們并不懷疑在極端缺血缺氧的實驗條件下,線粒體呼吸鏈及細胞會發生凋亡的情況,但在實際訓練中很難發生導致極端缺血缺氧的情況。即使有這種訓練手段,運動員對訓練負荷的生物自我保護機制也不允許導致細胞死亡的極度缺血缺氧現象的發生。
再次,在氧不足狀態下,細胞內化學反應過程的限制性負反饋抑制,也不會讓絕對的缺氧情況長時間存在。機體化學反應底物增加的同時,會減慢代謝的速度,這種情況為減少細胞傷害提供了最直接的保護限制。簡單來說,與電路過載自動跳閘的道理類似。大量實驗也支持人體在氧不足狀態下的自適應機制。缺氧是引起細胞損傷的重要原因,而缺氧誘導因子HIF-1是新發現的在氧濃度改變的一系列自適應反應中的重要調節因子。HIF-1對腫瘤細胞糖代謝的影響,主要通過低氧誘導的糖酵解代謝酶的增加而起作用。Lu等發現,HIF-1α不僅可以調節細胞糖酵解代謝酶的表達,糖酵解代謝的產物乳酸和丙酮酸也可以促進HIF-1α的穩定,促進HIF-1α誘導的基因轉錄,構成一個正反饋循環,但機制還不清楚[7]。眾多研究表明,除實驗室條件下的特殊無氧限制情況外,競技運動中的無氧與低氧分級訓練即使按照最原始的“應激”適應理論,所謂的無氧能量代謝在生物邏輯上只能造成線粒體系統的效率重調而不是萎縮。比如線粒體自噬的雙向調節、凋亡現象等,都是線粒體系統效率重調過程的正常表現。這才是生命系統與環境變化動態調適的生存原理。
另外,無氧的說法也值得斟酌,因為無論能量消耗的容積表達功率多大、時限多窄,能量穩態最終還是通過氧化還原過程來實現的,無氧或氧不足只是能耗與氧恢復的即時動態關系表達而已。所謂無氧訓練的正確理解應當是能量容量功率容積的極大化,是生命生存遺傳機制的能量時間表現,后天訓練中短暫的間歇性氧不足,可以逐步重構能量容量功率容積的極大化與線粒體系統的穩態調適關系。沒有無氧訓練,限制時間的能量容量功率容積的極大化,就不能滿足速度競賽的生物要求。
國內學者對線粒體的研究集中在運動性疲勞與線粒體的關系,包括能量耗竭學說、代謝產物堆積學說、離子代謝紊亂學說、氧自由基學說等。研究認為,急性運動中線粒體ROS調控能量代謝與抗氧化系統的平衡,ROS在運動中線粒體能量轉換和抗氧化平衡中發揮“分子開關”的作用。美籍華裔學者吉力立首次發現運動誘導線粒體ROS可通過激活NF-κB上調MnSOD表達。
研究指出,線粒體是高度動態的細胞器,可以從4點進行考慮:線粒體融合(fusion)提高ATP轉換效率,減少損傷,利于修復;線粒體分裂(fission)和移動(movement)利于細胞內不同區域線粒體執行各自功能;線粒體自噬(mitophagy)清除完全損傷的線粒體——線粒體質量控制體系;線粒體生物合成(biogenesis)維持細胞健康線粒體的閾值。
急性運動中骨骼肌線粒體趨向分裂,有助于提高ATP的輸出效率;運動后恢復期線粒體趨向于網絡化的融合過程,維持能量代謝的效率,有利于能量的節省化——耐力訓練的深遠效應。線粒體營養素——羥基酪醇可刺激PGC-1α和p53通路,促進線粒體融合,抑制分裂和自噬,上調線粒體復合體及抗氧化酶系表達,從而抑制劇烈運動對骨骼肌線粒體的損傷。線粒體穩態受細胞核DNA與線粒體DNA共同調控,線粒體ROS是諸多細胞生理事件的必要條件;ROS是線粒體與細胞核cross-talk的信使,其水平決定其生物學效應。運動訓練通過干預ROS調控細胞核和線粒體 cross-talk,不同水平的外源性H2O2雙向調控成肌細胞線粒體的動態重構,雷帕霉素靶蛋白mTOR參與不同水平H2O2對PGC-1α和線粒體重構的雙向調控。
從前面的分析可知,氧耗對應的是能量消耗,能量消耗對應的是輸出功的大小,動作鏈輸出功的增加才是競技訓練的發展方向。僅以部分條件標準并不統一的動物線粒體實驗結果,就推論出無氧訓練導致線粒體萎縮的指導性訓練意見,提倡進行所謂最大攝氧量55%~75% 水平的低功率有氧訓練極不嚴謹。實際上,這不僅是一個有爭議的生物科學問題,也是一個嚴重違反競賽目標訓練生物適應邏輯導向的問題。我國多數長距離項目的教練員與運動員,訓練艱苦異常而成績依然不佳,原因可能很多,但基于上述錯誤認識,很難達到競賽成績的提高與突破。
氧適應狀態分子網絡調適機制。在對人體缺氧機制系統性的認識方面,目前只有2019年諾貝爾醫學生理學獎研究內容進行了有針對性的表述。諾貝爾醫學生理學獎獲得者經過長時間研究,發現細胞都存在的缺氧誘導因子(HIF),是由結構同源的α亞基和相同的β亞基組成的異二聚體轉錄因子,目前已知包括HIF-1、HIF-2、HIF-3這3個成員。HIF-1α和ARNT(HIF-β)在缺氧的情況下引起缺氧調適,而在正常氧水平條件下又通過泛素復合蛋白VHL與HIF-1α作用引起其降解,恢復氧活動正常水平。缺氧與氧恢復因子轉錄水平的研究成果,對氧調適機制有更進一步的闡釋。從生物原理上分析,總功功率訓練與氧適應狀態調適的關系,才是競技訓練中應關注與深入研究的問題。何云凌利用對低氧敏感的PC12 細胞系,通過實驗比較不同氧濃度、不同低氧處理時間對不同接種密度下PC12細胞存活能力的影響。結果表明:經10%低氧處理24 h和48 h后,PC12細胞存活能力都顯著增加;而0.3%低氧條件處理24 h或48 h,均明顯抑制PC12細胞的存活。這說明在適度低氧條件下,細胞的抗氧化能力穩定,ROS 含量沒有明顯改變;而在嚴重低氧條件下,細胞的抗氧化能力降低,ROS 含量增加。研究表明,細胞在低氧條件下通過激活低氧誘導因子和BNIP3介導的PI3K通路誘導線粒體自噬,減少ROS的產生,促進細胞的存活和低氧適應[8]。申去非等[9]研究平原短期高強度間歇運動訓練(HIT)對急進高原青年男性淋巴細胞線粒體DNA氧化損傷的影響,發現平原階段短期高強度間歇訓練可提高機體高原低氧環境下淋巴細胞線粒體的抗氧化能力和能量代謝水平。研究[8-9]表明,無氧發生之前的低氧分級訓練一般不會造成線粒體萎縮,而是促進其進行積極的調適活動。比如線粒體自噬的雙向調節,是生命系統與環境變化動態調適的生存原理。機體細胞在自己生命周期的每一刻,都在為生命的存在而進行最大限度的生命活動,比如應激的3個階段,這是細胞極限生存的最大化體現。腦的主動控制與復雜的細胞信息控制網絡自動反饋調節,保障我們能夠在各種極端環境條件下生存,發生順應于環境要求的從基因到細胞生命迭代的演化進化,這就是最簡單的生命活動原理。
在競技訓練的生物原則上,應意識到:堅持生命與能量代謝極限環境動態自組織調適的訓練生物原理,倒計時設計與組織目標化訓練程式,才是最重要的訓練問題。以徑賽項目為例,隨著距離的延長,ATP發生的狀態在改變,但這并不代表ATP只能在有氧狀態才能產生,時間約束是ATP發生與狀態變化的根本原因。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
小學科學(學生版)(2018年3期)2018-04-18 12:34:19
知識經濟·中國直銷(2017年10期)2017-11-07 02:39:52
知識經濟·中國直銷(2017年3期)2017-04-16 03:08:01
文學少年(有聲彩繪)(2017年2期)2017-03-06 08:10:54
