廣西不同氣候條件下馬尾松中齡林木本植物葉性狀
2020-11-03 06:38:56潘小梅馬姜明楊章旗盧忠耐零天旺
廣西林業科學 2020年3期
關鍵詞:植物
菅 瑞,潘小梅,馬姜明,楊章旗,郭 飛,盧忠耐,零天旺
(1.廣西師范大學可持續發展創新研究院,廣西桂林 541001;2.珍稀瀕危動植物生態與環境保護教育部重點實驗室,廣西桂林 541001;3.廣西壯族自治區林業科學研究院,廣西南寧 530002;4.廣西國有派陽山林場,廣西寧明 532500;5.環江毛南族自治縣華山林場,廣西環江 547100;6.橫縣鎮龍林場,廣西橫縣 530327)
植物功能性狀是由Díaz 和Cabido 在總結前人對植物性狀研究的基礎上首次提出的,是指一切對植物的定居、生存和適應有著潛在重要影響的,或與資源獲取、利用和分配等相關的屬性[1],這些屬性是反映跨區域和跨尺度比較植物特征的指標,在植物與環境條件關系中發揮著重要作用[2],能單獨或聯合反映植物資源利用策略[3]。葉性狀一直是生態學家研究的熱點之一,近年來關于環境梯度的葉性狀研究主要涉及緯度梯度[4]、降水梯度[5]和海拔梯度[6]等環境因子,從不同氣候帶的角度研究的較少。
馬尾松(Pinus massoniana)廣泛分布于我國17個省(自治區、直轄市)[7],因具有適應能力強、材質好、生長快、耐干旱、耐瘠薄和抗蟲性強等特點,成為我國南方主要造林滅荒樹種之一[8]。隨著馬尾松種植的規模化和集約化,馬尾松得以速生,但同時地力衰退現象已成為馬尾松可持續經營的主要障礙。本研究以不同氣候條件下馬尾松中齡林中的木本植物為研究對象,從葉性狀特征入手,探究馬尾松中齡林葉性狀隨環境梯度的變化規律及反映馬尾松中齡林木本植物的關鍵葉性狀,揭示馬尾松中齡林木本植物沿環境梯度的生態策略,為預測環境變化對植物生存以及生態系統結構和功能的影響提供依據。
1 材料與方法
1.1 試驗地概況
試驗地位于廣西寧明縣國有派陽山林場(106°30'~107°15'E,21°15'~22°30'N),海拔200 ~800 m;屬北熱帶季風氣候,年均氣溫21.8 ℃,年均降水量1 475 mm;土壤以赤紅壤為主。廣西橫縣鎮龍林場(109°08' ~109°19'E,23°02' ~23°08'N),海拔400 ~700 m;屬南亞熱帶季風氣候,年均氣溫21.5 ℃,年均降水量1 477.8 mm;土壤多為赤紅壤,呈酸性或微酸性。廣西環江毛南族自治縣華山林場(108°06' ~108°38'E,25°6' ~25°31'N),海拔300 ~600 m;屬中亞熱帶季風氣候,年均氣溫19.8 ℃,年均降水量1 402 mm;土壤多為黃壤和紅壤。
1.2 樣地設置
在3 個試驗地中分別選取1 個典型馬尾松中齡林作為樣地(表1)。選擇無干擾、立地條件基本一致的地段設置樣方,每個樣地設置3個樣方,樣方面積為20 m×20 m,每個樣方內設置4 個10 m×10 m的小樣方。對各樣方內的木本植物種類、樹高、胸徑和冠幅等進行群落學調查,并記錄經緯度、海拔、坡向和坡度等情況。
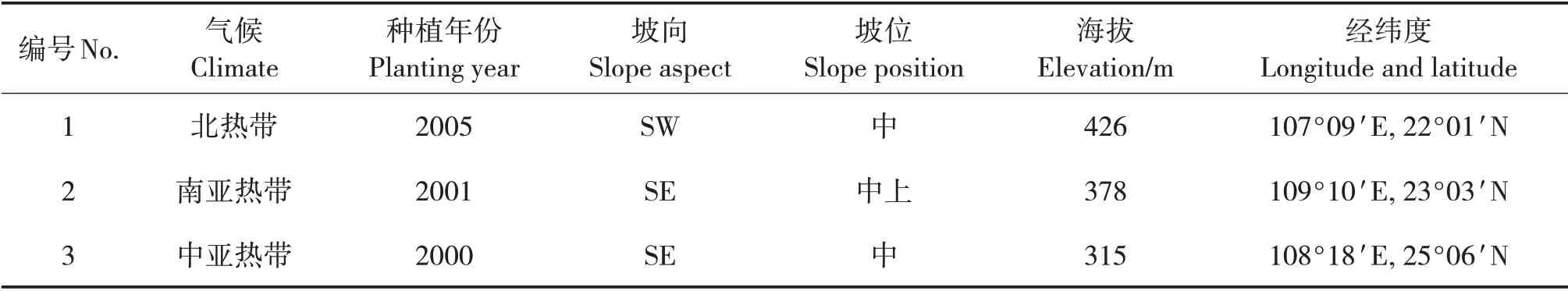
表1 樣地基本情況Tab.1 Basic conditions of sample plots
1.3 試驗方法
選取3 株生理形態健康成熟的植株,從不同方向采集10 片健康完整、大小相似的葉片,共30 片。去葉柄后置于濕潤的濾紙中,裝入自封袋后,放入有冰袋的保溫包中儲存。采集當天帶回實驗室后用水濕潤,放在5 ℃的冰箱中儲藏12 h,取出后迅速用吸水紙吸干葉片表面的水分,用電子天平稱重(精確至0.000 1 g)并編號,得到葉飽和鮮重(SFW)。用游標卡尺測定葉厚度(LT,精確至0.1 mm),用葉面積儀測量葉面積(LA,cm2)。葉片樣品置于60 ℃的烘箱烘至恒重后用電子天平測定葉干重(DW,g)。計算得到比葉面積(SLA,cm2/g)、葉干物質含量(LDMC,g/g)和葉組織密度(LTD,g/cm3),計算公式如下:

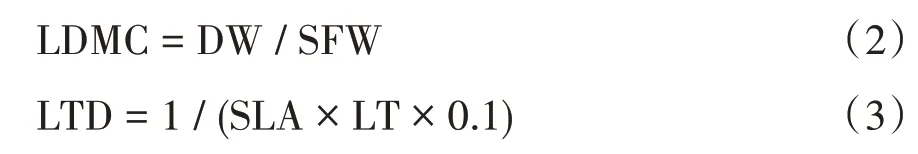
1.4 數據處理
對不同氣候條件下馬尾松中齡林喬木層和灌木層木本植物的6 個葉性狀(LT、DW、LA、LDMC、SLA和LTD)進行Pearson相關性分析。
2 結果與分析
2.1 不同氣候條件下馬尾松中齡林木本植物葉性狀的變化
LT、LDMC、DW 和LTD 隨緯度的升高呈上升趨勢;LA 和SLA 隨緯度的升高呈下降趨勢(圖1)。
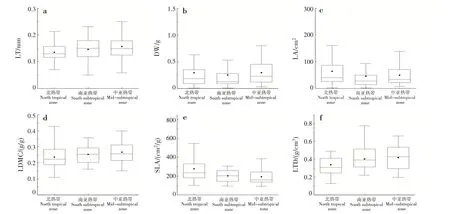
圖1 葉性狀在不同氣候條件下的變化Fig.1 Changes of leaf traits in different climatic conditions
為進一步探討葉片功能性狀的變化規律,對喬木層和灌木層植物葉性狀隨緯度的變化趨勢進行分析(圖2)。喬木層LT、LDMC 和LTD 隨緯度的升高呈上升趨勢,DW、LA 和SLA 呈下降趨勢;灌木層LT、LDMC 和LTD 隨緯度的升高呈上升趨勢,SLA 呈下降趨勢,DW 和LA 變化趨勢不明顯。喬木層植物葉片LDMC和LTD在南亞熱帶和中亞熱帶均大于灌木層,SLA和LT則小于灌木層。
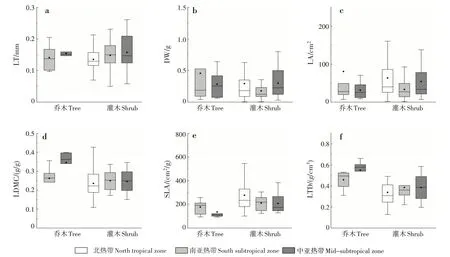
圖2 不同氣候條件下喬木層和灌木層植物葉性狀的變化Fig.2 Changes of leaf traits in tree layer and shrub layer under different climatic conditions
2.2 相關性分析
北熱帶灌木層DW與LDMC、LA、LT和LTD,LDMC 與LT和LTD 以及LA 與LT均呈顯著正相關;DW與SLA、LDMC 與LA 和SLA、LA 與LTD、LT與SLA 和LTD以及SLA與LTD均呈顯著負相關(表2)。
南亞熱帶喬木層DW 與LA 和LT、LDMC 與LT和LTD、LA與LT和SLA均呈顯著正相關;DW與LDMC 和LTD、LDMC 與LA 和SLA、LA 與LTD、LT 與SLA、SLA 與LTD 均呈顯著負相關(表3)。灌木層DW 與LA 和LT、LDMC 與LTD、LA 與LT 均呈顯著正相關;DW 與SLA,LDMC與LA、LT和SLA,LA與SLA和LTD,LT 與SLA 和LTD 以及SLA 與LTD 均呈顯著負相關。
中亞熱帶喬木層DW 與LDMC、LA、LT 和LTD,LDMC 與LA、LT 和LTD 以及LA 與LT 和LTD 均呈顯著正相關;SLA 與DW、LDMC、LA 和LT 均呈顯著負相關(表4)。灌木層DW 與LDMC、LA 和LT,LDMC與LTD以及LA與LT和SLA均呈顯著正相關;DW與SLA 和LTD、LDMC 與LA 和SLA、LA 與LTD、LT 與SLA和LTD以及SLA與LTD均呈顯著負相關。
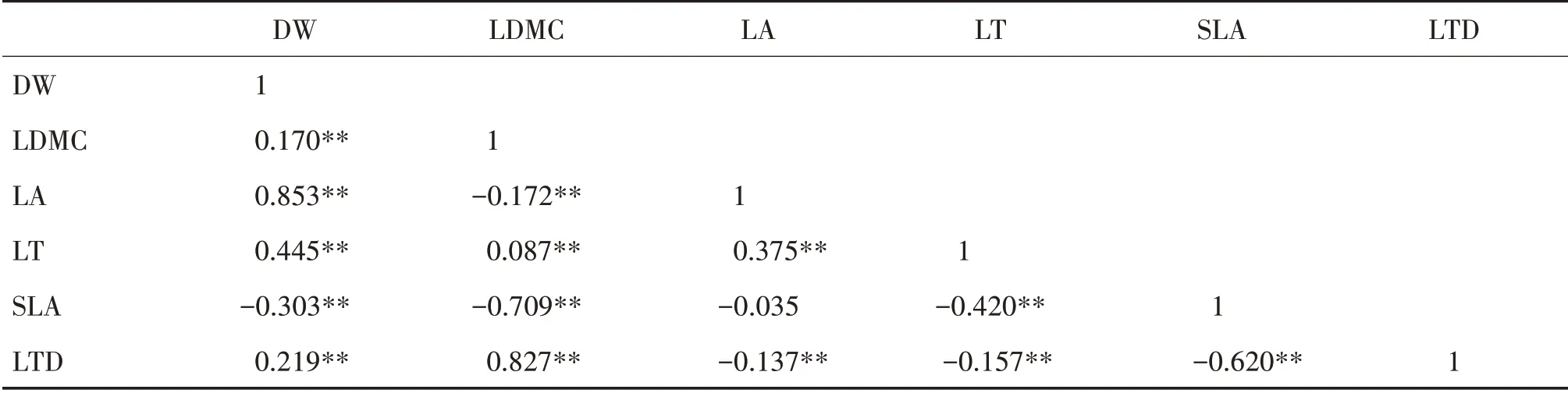
表2 北熱帶灌木層葉性狀的Pearson相關性分析Tab.2 Pearson correlation analysis of leaf traits of shrub layer in north tropical zone
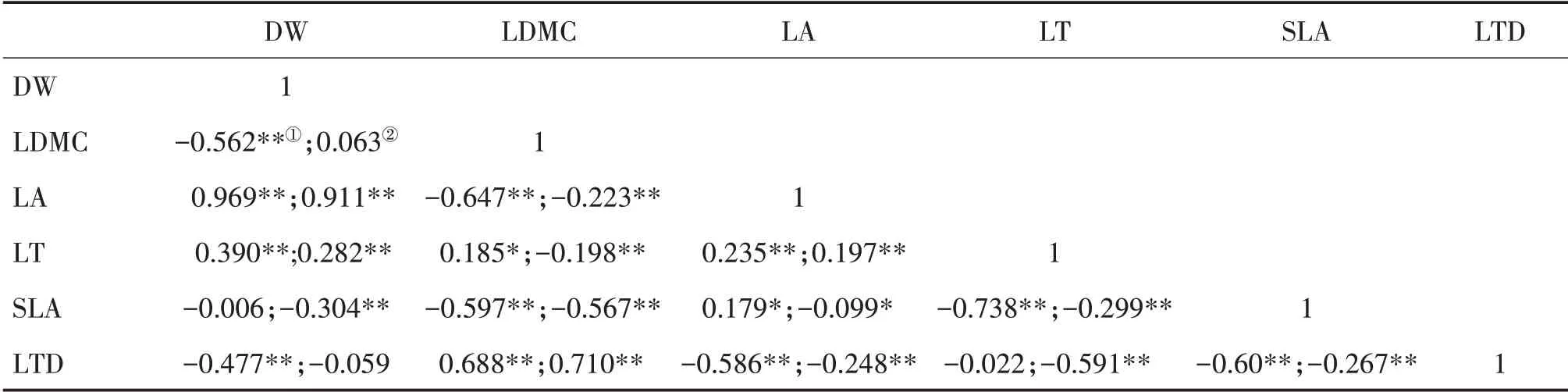
表3 南亞熱帶喬木層①和灌木層②葉性狀的Pearson相關性分析Tab.3 Pearson correlation analysis of leaf traits of tree layer①and shrub layer②in south subtropical zone
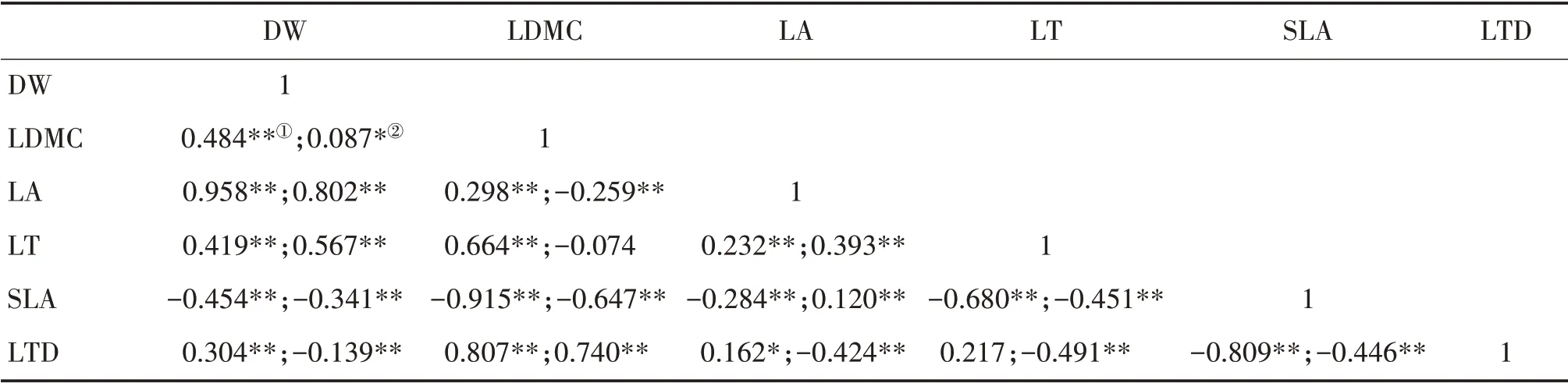
表4 中亞熱帶喬木層①和灌木層②葉性狀的Pearson相關性分析Tab.4 Pearson correlation analysis of leaf traits of tree layer①and shrub layer②in mid-subtropical zone
2.3 主成分分析
南亞熱帶喬木層葉性狀前2個主成分可以解釋葉性狀總方差的95%,其中PC1 可以解釋總方差的60%,PC2可以解釋總方差的35%;LA、LDMC和LTD對PC1 的貢獻率較大;SLA 和LT 對PC2 的貢獻率較大(圖3a)。第一主成分突出反映植物對光的吸收和對碳的獲取能力,第二主成分突出反映植物對光照和水分等異質性環境因子的資源利用策略。中亞熱帶喬木層葉性狀前2個主成分可以解釋葉性狀總方差的90%,其中PC1 可以解釋總方差的65%,PC2可以解釋總方差的25%;SLA 和LDMC 對PC1的貢獻率較大;LA對PC2的貢獻率較大(圖3b)。第一主成分突出反映植物抵御外界干擾的能力,第二主成分突出反映植物對光的吸收和對碳的獲取能力。
北熱帶灌木層葉性狀前2 個主成分可以解釋葉性狀總方差的82%,其中PC1 可以解釋總方差的46%,PC2 可以解釋總方差的36%;LDMC、LTD 和SLA 對PC1 的貢獻率較大;LA 對PC2 的貢獻率較大(圖3c)。第一主成分突出反映植物抵御外界干擾的能力,第二主成分突出反映植物對光的吸收和對碳的獲取能力。南亞熱帶灌木層葉性狀前2個主成分可以解釋葉性狀總方差的77%,其中PC1 可以解釋總方差的42%,PC2 可以解釋總方差的35%;LA和LTD 對PC1 的貢獻率較大;SLA 和LDMC 對PC2的貢獻率較大(圖3d)。第一主成分突出反映植物對光的吸收和對碳的獲取能力,第二主成分突出反映植物抵御外界干擾的能力。中亞熱帶灌木層葉性狀前2 個主成分可以解釋葉性狀總方差的83%,其中PC1 可以解釋總方差的49%,PC2 可以解釋總方差的34%;LA 和LTD 對PC1 的貢獻率較大;SLA和LDMC 對PC2的貢獻率較大(圖3e)。第一主成分突出反映植物對光的吸收和對碳的獲取能力,第二主成分突出反映植物抵御外界干擾的能力。
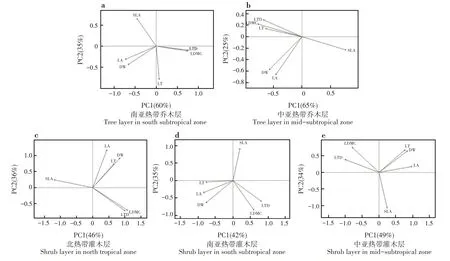
圖3 葉性狀的主成分分析Fig.3 Principal component analysis of leaf traits
3 結論與討論
3.1 不同氣候條件下植物葉性狀的比較
LA隨著緯度的升高,總體呈下降趨勢。葉面積隨著年均溫的降低而減小[9],與本研究結果一致。緯度越高,氣溫越低,葉接受光能和同化二氧化碳的時間縮短,引起葉片小型化。小葉的呼吸和蒸騰成本更低[10],可以降低植株的維持消耗。LT 隨著緯度的升高,呈上升趨勢。植物葉片LT的增加是植物對低溫適應的結果[11]。LDMC 能反映植物獲取資源的能力,LDMC 越高,植物獲取資源的能力越強[12]。LDMC 隨著緯度的升高,呈上升趨勢,SLA 則相反。吳漫玲等[13]對星斗山混交林木本植物不同海拔梯度的研究表明,隨海拔升高,溫度降低,LT 和LDMC 升高,LA 和SLA 降低,與本研究結果相似。低溫是植物SLA 沿海拔梯度上升而下降的根本原因,在低溫環境下,葉肉細胞增大和細胞壁特別是表皮細胞壁增厚,所以LT增加,SLA下降[14]。LTD隨著緯度的升高,呈上升趨勢。鐵桿蒿(Tripolium pannonicum)在森林帶(溫度較高的植被帶)擁有最低的LTD[15],與本研究結果一致。在高溫環境下,通過增加LTD 可防止高溫傷害[15]。
葉結構性狀在不同生活型適應策略不同。喬木層和灌木層的LT和LTD隨緯度的升高,均呈上升趨勢,SLA 則相反。SLA 表征植物潛在相對生長速率或最大光合速率,能指示植物在不同環境下利用資源的能力[16]。本研究表明,喬木層SLA 均小于相應氣候帶的灌木層,這與莫燕華等[17]的研究結果一致。SLA 大的植物捕獲光的能力強,通常光線越弱的冠層形成的葉子SLA 越大[18]。灌木層光線相對喬木層弱,SLA 的投資大。喬木層植物相對于灌木層植物具備更好地適應資源貧瘠的能力,灌木層植物則在獲取資源(如光照)的能力方面相對較強[19]。本研究表明,喬木層LT 均小于對應的灌木層,喬木層LDMC 大于灌木層。黃小等[20]對不同生活型葉性狀的研究表明,LT表現為喬木<灌木<木質藤本,LDMC 表現為喬木>灌木>木質藤本,與本研究結果一致。喬木層植物在葉片受到強光輻射時,通常會加大對LDMC的投資[17]。
3.2 植物葉性狀之間的相關性
植物功能性狀間緊密的相關關系普遍存在,這是由于物種間在生長發育及形成適應性機制的過程中相互作用,表現出相對應的權衡策略[21]。LA與LT在馬尾松喬木層和灌木層均呈極顯著正相關,這與滇中喀斯特植物[22]的研究結果一致。DW 與LA 和LT、LDMC 與LTD 在馬尾松喬木層和灌木層均呈極顯著正相關,這與滇中喀斯特植物[22]、瑯岐島優勢植物[23]等研究結果一致,表明DW隨LA和LT的增大而增大,LDMC 隨LTD 的增大而增大,進一步表明LDMC 越大LTD 越高,葉也越堅韌,越能抵抗不良的生長物理因子[24]。
DW 與SLA、LDMC 與LA、LA 與LTD、LT 與SLA和LTD、SLA 與LTD 在馬尾松灌木層均呈顯著負相關,這與不同林齡馬尾松人工林針葉植物[25]、井岡山鹿角杜鵑(Rhododendron latoucheae)群落植物[21]、桂林巖溶石山檵木(Loropetalum chinense)群落常見植物[17]和星斗山常綠落葉闊葉混交林木本植物[13]研究結果一致。SLA和LDMC是葉片最為關鍵的性狀,能綜合反映植物利用資源的能力[26],是植物適應生境的關鍵因子,具有重要的生態學意義。LDMC與SLA在馬尾松喬木層和灌木層均表現為顯著負相關,即隨著LDMC 的增加,SLA 減小,使得葉片內部水分向葉片表面擴散的阻力增大,降低植物內部水分散失[27]。
3.3 植物葉性狀適應策略
主成分分析表明,在6 個葉性狀指標中,LA、LDMC、LTD 和LT 可作為反映南亞熱帶喬木層葉性狀的重要指標,主要表征植物抵御外界不利環境和對水熱環境條件適應的能力。LDMC 和LA 可作為反映中亞熱帶喬木層葉性狀的重要指標,主要表征植物抵御外界干擾及不利環境的能力。喬木層植物葉片LDMC 均大于相應氣候帶的灌木層,表明喬木層植物具有“緩慢投資-收益”,即“保守型”葉經濟譜的特點[28-29]。SLA、LTD 和LA 可作為反映北熱帶、南亞熱帶和中亞熱帶的馬尾松中齡林灌木層葉性狀的重要指標,表征植物獲取資源及對干濕環境的適應能力。灌木層植物葉片SLA均大于相應氣候帶的喬木層,表明灌木層植物具有“快速投資-收益”,即“獲取型”葉經濟譜的特點[28-29]。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
