CD73通過AMPK/mTOR信號通路調控脊髓損傷機制研究①
2020-11-10 01:48:54邵明昊靳明明鄭超君馬曉生呂飛舟
中國免疫學雜志 2020年19期
邵明昊 靳明明 周 萍 鄭超君 朱 巍 馬曉生 呂飛舟
(復旦大學附屬華山醫院脊柱外科,上海 200040)
脊髓損傷(spinal cord injury,SCI)是脊柱損傷最嚴重的并發癥,往往導致損傷節段以下肢體嚴重功能障礙。全世界約有250萬人患有SCI,且以每年約130 000例的速度增加[1]。SCI的病理過程分為原發性和繼發性損傷2個階段,而SCI后的繼發性損傷階段已被證明是關鍵的治療時期,此期間可以實施神經保護治療以阻止神經進一步壞死、促進功能恢復。現有證據表明多種代謝因子在SCI繼發性損傷的發病機制中具有重要作用[2]。
腺苷活化蛋白激酶(AMP-activated protein kinase,AMPK)是能量代謝的主要傳感器,通過與ATP、ADP、AMP等結合,在能量水平下降的反應中發揮重要作用。與ATP結合時,AMPK活性被抑制,在能量釋放期間,細胞內AMP水平升高,導致AMPK活化,并增加細胞分解代謝能力,產生更多ATP。作為調節因子,雷帕霉素靶蛋白(mammalian target of rapamycin,mTOR)在細胞內營養和生長因子豐富時被激活[3]。
外5′-核苷酸酶(CD73)是一種大小約為70 kD的糖基化蛋白,位于外質膜上,其功能是將AMP水解為腺苷和磷酸鹽[2,4,5]。超過85%的小鼠AMP在CD73的作用下迅速降解為腺苷,最終使細胞內cAMP積累,同時抑制局部免疫反應,因此認為CD73與炎癥相關的腦發育疾病密切相關[1,6-8]。此前,課題組確定了CD73在神經保護作用中的內在作用,這種作用是通過介導巨噬細胞/小膠質細胞極化發生的[9-12]。本研究旨在探討CD73對脊髓損傷后AMPK的影響。
1 材料與方法
1.1材料
1.1.1實驗動物 C57BL/6 CD73基因敲除(KO)雄性小鼠由美國俄克拉何馬州醫學研究基金會湯普森教授惠贈,野生型(WT)雄性C57BL/6小鼠購自上海SLAC實驗動物有限公司。所有實驗程序均經復旦大學附屬華山醫院倫理委員會批準并按照其指導方針執行。
1.1.2主要試劑與儀器 TruSeq鏈mRNA-LT樣品制備試劑盒(Illumina,San Diego,California,USA);三唑試劑(Invitrogen,San Diego,CA,USA);ImProm llTM逆轉錄酶(Promega,Madison,WI,USA)二級抗體(Jackson Immunology Research,West Grove,PA,USA);倒置顯微鏡(Ti2,尼康,日本);凝膠成像系統(UVP LLC,Upland,CA,USA);凝膠Pro分析儀軟件(Media controlnetics,Rockville,MD,USA)
1.2方法
1.2.1SCI小鼠建模 咬骨鉗切除小鼠T8~T9椎板,保護硬膜完整,Dumont型鑷子(尖部直徑為0.2 mm)造成脊髓壓迫,脊髓背側壓迫面積為0.2 mm,持續20 s。術后分別飼養,人工排空膀胱,2次/d。 對照組小鼠接受假手術椎板切除術,但脊髓無損傷。
1.2.2qPCR檢測 取小鼠脊髓,TRIzol法提取脊髓損傷組織總RNA并確定總RNA的純度和濃度,使用逆轉錄試劑盒將mRNA逆轉錄為cDNA,取適量cDNA配置PCR 反應體系, 以 GAPDH 為內參。實驗重復3次,采用2-ΔΔCt法計算基因相對表達量。
1.2.3RNA測序 使用TruSeq鏈mRNA-LT樣品制備試劑盒對WT和KO小鼠脊髓組織中RNA進行Illumina RNA測序。Illumina HiSeq 2500對RNA序列庫測序,設置2×50 bp,各樣本讀數為7×107~1×108。隨后對RNA序列數據中的KEGG進行綜合分析。
使用三唑試劑從不同治療組小鼠脊髓組織中提取總RNA,經ImProm llTM逆轉錄酶反轉錄為cDNA。SYBR試劑對mRNA進行定量分析。以GAPDH為內參。數據分析采用2-ΔΔCt法。
1.2.4免疫組化和免疫熒光測定 術后第3天提取脊髓組織樣本,將部分樣品浸泡在1%牛血清白蛋白和0.3%Triton X-100中1 h以阻止非特異性反應。分別以抗CD73、抗Neun、抗IL-1β、抗TNF-α、抗Caspase-3抗體4℃孵育過夜。PBS清洗后用相應的二抗室溫孵育2 h,Dy488和Dy594結合二抗(1∶1 000),以進行免疫組化和免疫熒光評估。成像使用倒置顯微鏡,組織切片分別行HE、Nissl、IHC和TUNEL染色。
1.2.5Western blot分析 用RIPA緩沖液提取不同治療組小鼠脊髓組織總蛋白,BCA法分析其濃度。采用SDS-PAGE分離蛋白樣品,然后將其轉移至硝化纖維素膜。5%脫脂牛奶在TBST中孵育 1 h,以阻止非特異性反應。4℃下將膜與抗體(1∶1 000)孵育過夜。在室溫下將HRP結合的二抗添加到膜中孵育1 h,并用電化學發光法使膜可見。使用凝膠成像系統對結果進行成像分析,凝膠Pro分析儀軟件對結果進行測量。
1.3統計學分析 使用GraphPad Prism 5.02軟件對結果進行統計學分析。兩組間差異采用配對t檢驗,Pearson相關檢驗用于確定兩組間相關性。P<0.05為差異有統計學意義。
2 結果
2.1SCI組織中炎癥和凋亡相關因子增加 qPCR結果表明,脊髓損傷后炎癥因子IL-6、IL-1β、TNF-α和Caspase-3表達提高(圖1)。HE和Nissl染色結果表明脊髓損傷后神經細胞紊亂、腫脹和尼氏體減少更為明顯(圖2A、B)。IHC也證實,與正常對照組相比,SCI組小鼠IL-1β、TNF-α和Caspase-3表達上調(圖2C)。
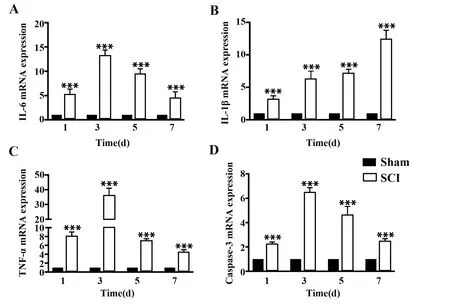
圖1 SCI小鼠中炎癥因子和凋亡相關因子表達Fig.1 Expressions of inflammatory and apoptosis-related factors in SCI ratsNote:Compared with Sham group,***.P<0.001.

圖2 SCI小鼠中脊髓組織損傷情況(×400)Fig.2 Spinal cord tissue damage in SCI rats(×400)Note:A.HE staining;B.Nissl staining;C.IHC staining.
2.2CD73缺失導致脊髓組織損傷加重 SCI后脊髓組織中CD73的表達在mRNA和蛋白質水平上均上調(圖3)。TUNEL染色結果表明,與野生型小鼠相比,SCI后CD73-KO小鼠脊髓組織神經元凋亡增加(圖4)。

圖3 CD73對SCI的影響Fig.3 Effect of CD73 on SCINote:Compared with Sham group,*.P<0.05,**.P<0.01,***.P<0.001.
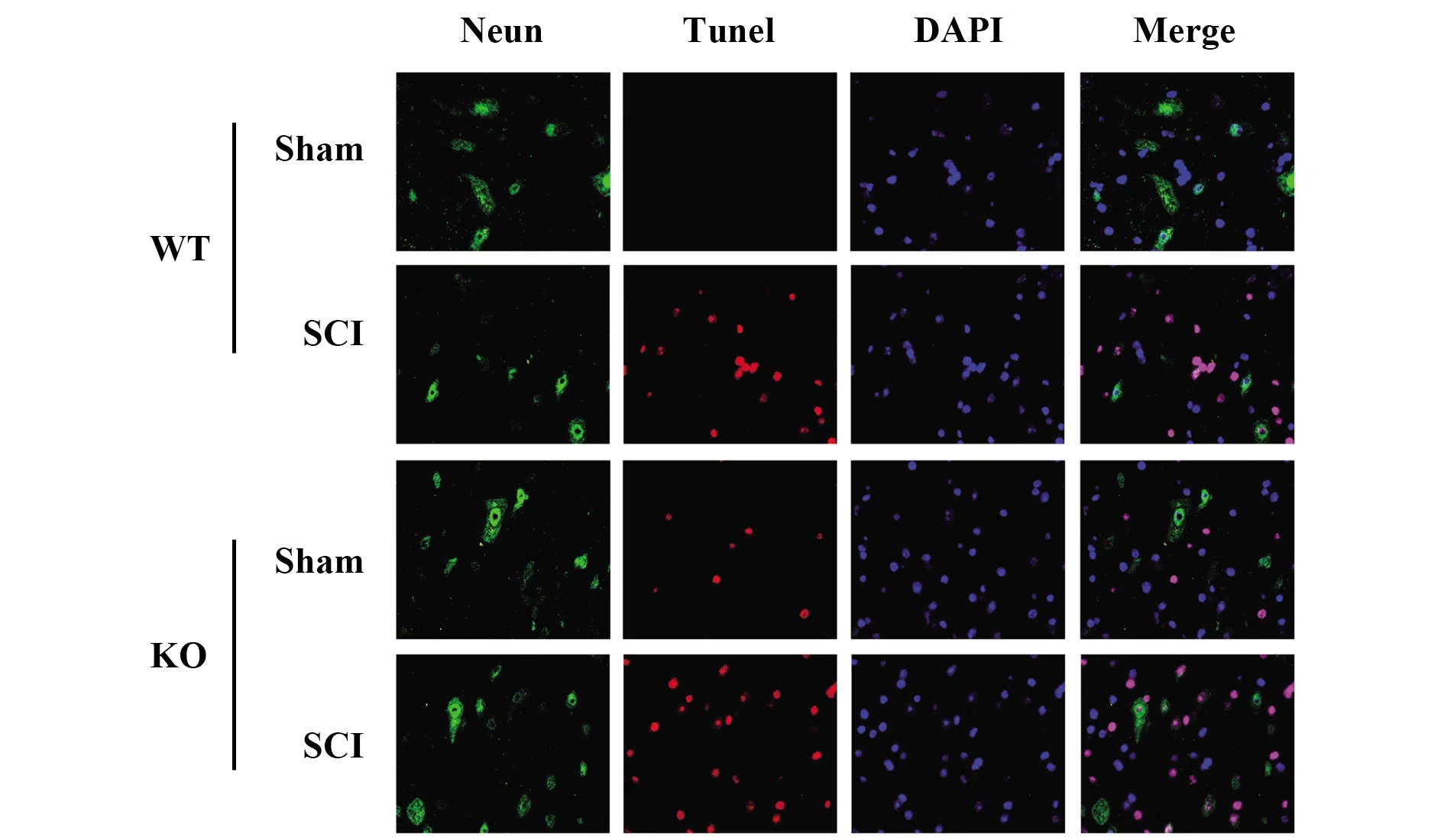
圖4 KO和WT小鼠SCI 7 d后TUNEL染色結果Fig.4 Result of TUNEL staining of KO and WT mice after 7 d of SCI
2.3SCI引起CD73-KO和WT小鼠代謝改變 WT和CD73 KO小鼠的RNA測序結果顯示,WT和假手術動物間存在1 649個mRNAs差異表達,與CD73 KO小鼠間有94個mRNAs差異表達。主要改變途徑包括炎癥反應、細胞對機械刺激的反應、鐵離子穩態、凋亡和AMPK途徑,其中mTOR途徑的下游靶點4Ebp1差異顯著,而AMPK途徑也被顯著激活。表明小鼠SCI后發生了代謝改變,CD73的敲除與以上代謝改變有關。
2.4CD73 KO小鼠AMPK的激活與細胞凋亡有關 與假手術組相比,CD73 KO小鼠的p-AMPK和p-ACC表達升高(圖5),表明SCI后AMPK通路的激活與p-mTOR和4Ebp1低表達相關。提示SCI通過AMPK/mTOR途徑介導細胞凋亡,損傷神經元活性。
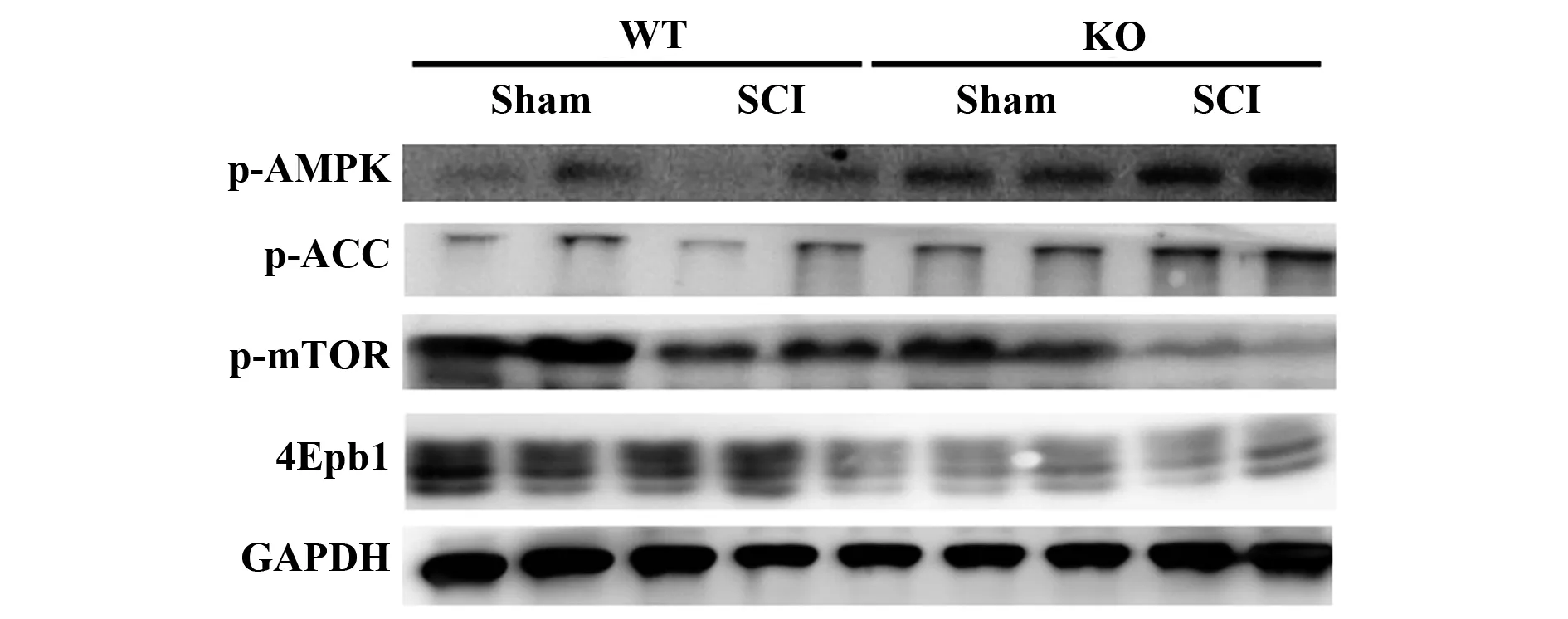
圖5 Western blot檢測p-AMPK、p-ACC、p-mTOR和4Epb1表達Fig.5 Expressions of p-AMPK,p-ACC,p-mTOR and 4Epb1 detected by Western blot
3 討論
SCI或外傷性腦損傷常導致神經病理狀態和隨之而來的功能恢復[10]。細胞凋亡是一種細胞系統內在的基因調控反應,維持正常條件下細胞穩態[8,13]。神經元凋亡是加重SCI的原因之一,既往研究表明,細胞因子的積累能夠激活星形膠質細胞,最終導致神經元凋亡[11,12]。本研究中,TNF-α、IL-1β、IL-6均在SCI組織中明顯增加,可引起嚴重的免疫反應;進一步研究結果顯示,Caspase-3和PARP在SCI小鼠中表達升高,說明SCI小鼠的脊髓組織內細胞凋亡情況更加嚴重。TUNEL結果顯示,細胞凋亡多發生于神經元細胞,且KO小鼠中細胞凋亡更加嚴重。為課題組進一步探究CD73在抑制神經元凋亡中的具體機制提供思路。
已有研究表明AMPK與AMP密切相關。在動物體內,AMPK的活性主要受細胞內AMP/ATP比值調節。體內多種條件,如缺血、缺氧、葡萄糖缺乏、饑餓、電刺激和熱休克,以及一氧化氮、三羧酸循環和氧化磷酸化的抑制劑,包括亞砷酸、抗霉素A、二硝基苯酚和疊氮化合物都可導致AMP/ATP比值上升,引發AMPK磷酸化[14,15]。CD73是一種糖基磷脂酰肌醇蛋白,在催化AMP生成磷酸和腺苷的過程中起重要作用。CD73在神經元和膠質細胞中含量豐富,局灶性腦缺血上調CD73的膠質表達[15]。課題組前期研究發現,CD73缺陷小鼠表現出更嚴重的運動功能障礙,與神經炎癥相關的組織遭到破壞,細胞凋亡增加。本研究觀察到WT、假手術動物和CD73 KO小鼠間mRNAs的差異表達存在于炎癥反應、細胞對機械刺激的反應、鐵離子穩態、凋亡和AMPK途徑。其中以mTOR途徑的下游靶點4Ebp1差異最為顯著,而AMPK途徑也被顯著激活。提示CD73對神經元凋亡的抑制作用可能通過AMPK途徑發揮作用。
既往研究發現,AMPK/mTOR信號通路與細胞凋亡有關[10,15]。本研究發現激活AMPK可抑制mTOR磷酸化,從而誘導神經元凋亡。結果表明CD73 KO小鼠模型AMPK磷酸化水平顯著升高,mTOR水平相應降低等都與細胞凋亡相關,表明CD73下調可能激活AMPK、抑制mTOR和4Ebp1信號從而增加細胞死亡。結果提示CD73/AMPK/mTOR/4EBP1通路在SCI中發揮作用,CD73的調節可能是抗凋亡的生存機制,為SCI治療提供參考。
綜上,SCI小鼠模型中,CD73能夠抑制脊髓繼發性損傷。下調CD73可通過調節AMPK/mTOR信號通路誘導更嚴重的神經元凋亡。CD73可能是SCI治療的靶點之一。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
學苑創造·A版(2020年9期)2020-10-13 09:41:02
中學生數理化·七年級數學人教版(2019年10期)2019-11-25 07:33:58
中學生數理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
中國衛生(2016年3期)2016-11-12 13:23:26
中國衛生(2014年12期)2014-11-12 13:12:52
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
七彩語文·畫刊(2012年3期)2012-04-29 00:00:00