應用東鄉野生稻回交重組自交系分析水稻耐低氮產量相關性狀QTL
2020-11-23 08:37:04吳婷李霞黃得潤黃鳳林肖宇龍胡標林
中國水稻科學 2020年6期
吳婷 李霞 黃得潤 黃鳳林 肖宇龍 胡標林
(1 江西省農業科學院 水稻研究所/水稻國家工程實驗室(南昌),南昌 330200;2 中國水稻研究所 國家水稻改良中心/水稻生物學國家重點實驗室,杭州 310006; 3 湖南省水稻研究所 農業農村部長江中下游秈稻遺傳重點實驗室,長沙 410125;*通信聯系人, E-mail: ylxiao2012@163.com,hubiaolin992@126.com)
水稻是我國最重要的糧食作物之一。水稻高產對確保國家糧食安全和社會穩定尤為關鍵,因而一直是水稻育種和生產的重要目標。化肥(尤其是氮肥)對促進水稻高產發揮了重要作用。氮素是影響水稻生長發育和產量的重要必需營養元素,也是影響稻田生態系統的關鍵營養元素。據統計,我國2/3以上耕地為中低產田,其中瘠薄型占27%[1]。我國南方稻區中低產田特別是低產田生態環境脆弱、基礎地力差,而低產田缺氮嚴重制約水稻增產潛力。增施氮肥成為保障產量重要措施,然而過量施用氮肥不僅增加水稻生產成本、降低生產效益[2],還會導致氮肥利用率下降[2]以及土壤及水體污染[3-5]等稻田農業生態系統問題,進而對生態環境和農業可持續發展構成威脅。由此,綠色超級稻的“第二次綠色革命”理念和國家農業化肥農藥零增長行動應運而生,耐低肥、節水抗旱、抗病蟲等“少投入”新品種的選育和推廣勢必助推我國綠色農業發展。提高水稻耐低氮能力或氮高效利用力能較好地解決這一水稻生產中的矛盾[6-8]。發掘耐低氮基因并研究其遺傳機理,將促進水稻耐低氮品種的分子育種,提高水稻氮肥有效利用率、生產效益,并改善土壤。
水稻耐低氮是一個復雜的數量性狀,受主效基因和和微效QTL 控制[9]。雖然通過傳統育種方法進行水稻耐低氮育種取得一些成效,但進展不大。然而,隨著生物技術的發展,QTL 分析成為解析水稻耐低氮等數量性狀的有效方法,為水稻耐低氮分子育種打下基礎。迄今,不同研究已定位到大量水稻不同生育時期的耐低氮性狀QTL,這些QTL 廣泛分布于水稻12 條染色體上[9-18]。在營養生長期耐低氮 QTL 方面,Nguyen 等[10]利用 IR64/Azucena 秈粳交群體檢測到63 個苗期農藝、生理性狀QTL 和14個 氮 肥 利 用 率 QTL ; Mahender 等[11]利 用HAN/WTR-1、CH448/WTR-1 和 Z413/WTR-1 等 3套遺傳群體進行了3 種施肥劑量、移栽后4 個不同發育階段的農藝生理性狀QTL 分析,定位到19 個相關QTL,且低氮耐性QTL 只在特定發育階段特異表達,與水稻不同發育階段基因的時空表達相關。而生殖生長期耐低氮 QTL 定位方面,馮躍等[12-13]利用協青早B/中恢9308 秈秈交群體在施氮和不施氮條件下分別鑒定到52 個產量相關性狀QTL 和17個株高、抽穗期的 QTL ; Cho 等[14]利用Dasanbyeo/DR22183 秈粳交群體在兩種氮肥水平下檢測到 20 個控制稻谷氮含量、秸稈氮含量、地上部總氮含量、收獲指數、產量、稻草收獲量和氮素利用率 QTL;Wei 等[15-16]利用珍汕 97/明恢 63 秈秈交群體分別檢測到15 個耐低氮性狀QTL、11 個氮素利用率QTL 和11 個產量QTL,并認為育種中可以實現水稻產量和氮肥利用率同步改良。相似地,Zhou 等[17]對產量、氮素利用率和氮素吸收率 QTL分析發現,分子聚合高氮素吸收和氮素利用率QTL位點有利于培育高產量、低需氮水稻品種。此外,Tong 等[18]對 3 種施氮水平下水稻產量及其構成因子進行了 QTL 分析,在高氮、中氮和低氮水平下分別檢測到 15、23 和19 個 QTL,且有 5、3 和5對上位性 QTL 與環境存在顯著互作,說明水稻耐低氮機制非常復雜。另外,上述研究所用遺傳群體主要為亞種間組合群體,而野生稻與栽培稻種間遺傳群體缺乏,野生稻耐低氮相關基因發掘有待加強。
在已定位的水稻耐低氮 QTL 中,由于低氮鑒定環境和評價性狀不同,不同研究間結果缺乏可比性,同時難以真正應用于水稻耐低氮分子育種,有待進一步精細定位和克隆。迄今已經分離一批涉及氮素吸收調控的基因,如qNGR9、OsARG、OsOAT、OsNRT2.3、OsPTR6、OsNAR2.1、NRT1.1B、ARE1、TOND1、OsNR2以及OsNPF6.1[19-27]。其中TOND1可提高水稻苗期耐低氮能力及稻谷產量[24];OsARG基因通過增加低氮下水稻單株穗粒數實現增產[25];NRT1.1B基因編碼硝酸鹽轉運蛋白,通過調控水稻根系微生物組來改變根際微環境,從而影響水稻秈粳亞種間的氮肥利用效率,且秈稻型NRT1.1B在水稻馴化中經歷人工選擇[26-27];OsNPF6.1基因編碼硝酸鹽轉運蛋白,為野生稻的稀有等位基因,可增強水稻氮吸收和氮肥利用率,提高低氮條件下的稻谷產量[22]。這些氮素調控基因的克隆以及qRDWN6BX[28]、qNUE6[29]等主效 QTL 精細定位為耐低氮水稻品種分子育種奠定良好的基礎,然而有關野生稻耐低氮基因克隆研究薄弱。野生稻作為栽培稻的野生祖先種,蘊藏著大量耐逆基因,是水稻育種改良的優異種質資源。
東鄉野生稻位于江西省東鄉縣崗上積鎮,是迄今發現分布最北的普通野生稻,具有耐寒、耐旱、耐貧瘠等耐逆特性。東鄉野生稻長期生存于貧瘠的自然環境(其原生境土壤全氮含量為 0.09%~0.16%[30]),耐低氮能力較強,是水稻耐低氮育種和基礎研究的重要資源。胡標林等[31]利用協青早B//東鄉野生稻/協青早B BC1F12回交重組自交系篩選耐低氮綜合指標,為耐低氮水稻選育提供了參考。本研究應用相同的遺傳群體考查兩種氮素水平下株高、穗長、有效穗數、穗實粒數、穗總粒數、著粒密度、結實率、千粒重和單株產量等9 個農藝性狀,進行耐低氮 QTL 分析,旨在鑒定耐低氮性狀QTL 以及東鄉野生稻有利等位基因,為水稻耐低氮分子育種提供參考依據。
1 材料與方法
1.1 試驗材料
研究材料包括237 份協青早B//東鄉野生稻/協青早B BC1F12的回交重組自交系及其親本,其中,協青早B 是矮敗型三系不育系協青早A 的保持系,屬亞洲栽培稻,東鄉野生稻是普通野生稻,屬亞洲栽培稻的祖先種。1998 年以東鄉野生稻為母本、協青早B 為父本進行雜交產生F1,再與協青早B 回交獲得BC1F1。2000 年在浙江杭州種植BC1F1群體,經單粒傳法至 BC1F5,2003 年在江西南昌種植BC1F6,繼續通過單粒傳法自交6 代,2009 年獲得237 份BC1F12回交重組自交系。
1.2 試驗設計
于 2012 年 5 月-11 月期間,將 237 份 BIL 株系及其親本種植于江西省農業科學院水稻研究所試驗田,期間日平均氣溫為25.7℃,降水量為1013 mm。試驗田統一翻耕平整后,取耕作層土壤(0?20 cm)土樣交江西省農業科學院農業資源環境研究所分析土壤氮、磷、鉀等含量:全氮1.69 g/kg,堿解氮178.2 mg/kg,有效磷11.5 mg/kg,有效鉀69.0 mg/kg。設未施氮(0 kg/hm2)和施氮(300.0 kg/hm2)2 個不同氮肥處理,磷、鉀肥施用同常規栽培一致,每個處理重復3 次,隨機區組排列。不同處理之間筑埂包膜以防串灌。施氮處理施尿素(折合純氮)300.0 kg/hm2,施過磷酸鈣125.0 kg/hm2,施氯化鉀125.0 kg/hm2,其中60%氮肥和全部的磷、鉀肥在秧苗移栽1 周后施用,40%氮肥在拔節期進行施用;未施氮處理除不施氮肥外,其他肥水管理同施氮處理一致。5 月20 日播種,6 月16 日單本栽插,株行距16.6 cm ×19.8 cm,各株系種3 行,每行10 株。試驗用水為農田灌溉用水,田間管理按常規栽培方式進行。在9 月6 日采取耕作層土樣由江西省農業科學院農業資源環境研究所進行檢測,分析土壤氮、磷、鉀等含量。其中,未施氮土壤全氮 1.64 g/kg,堿解氮152.6 mg/kg;施氮土壤全氮2.0 g/kg,堿解氮173.6 mg/kg。
1.3 性狀考查
1.3.1 株高
成熟后,隨機取各株系中間3 株植株測量株高,從植株基部測量至主穗穗頂的高度(不含芒長),并求平均值用于統計分析。
1.3.2 產量性狀
成熟后,收獲各株系中測定了株高性狀的3 個單株,室內自然晾干后,進行室內人工考種。分別考查有效穗數、每穗實粒數、每穗總粒數、結實率、穗長、著粒密度、千粒重和單株產量等8 個產量構成因子,分別計算3 個重復性狀均值用于統計分析。
1.4 耐低氮性QTL 分析
前期研究[32]構建了協青早B//東鄉野生稻/協青早 B 回交重組自交系群體的遺傳圖譜,該圖譜含133 個 SSR 和 12 個 InDel 標記,覆蓋水稻基因組1620.90 cM,標記間平均距離11.17 cM。結合該遺傳圖譜,應用軟件Windows QTL Cartographer 2.5(http:// statgen. ncsu.edu/qrtcart/WQTLCart.htm),采用復合區間作圖法(composite interval mapping,CIM)進行QTL 檢測。以LOD 值2.5 作為檢測閾值,將LOD值最高處所對應的位點作為一個QTL,并統計該 QTL 的加性效應和表型貢獻率。采用McCouch[33]提出的命名原則進行QTL 命名。
2 結果與分析
2.1 BIL 群體及其親本在施氮和未施氮條件下9個表型性狀的表現
考查BIL 群體及其親本在施氮和未施氮條件下的株高、穗長、著粒密度、千粒重、每穗實粒數、每穗總粒數、結實率、有效穗數和單株產量等9 個農藝性狀,結果列于表1。
親本東鄉野生稻與協青早B的各農藝性狀在施氮與未施氮兩種條件下變異較大,且其變化趨勢和幅度不同,其中雙親的有效穗數、每穗總粒數和單株產量等 3 個性狀測定值在不同處理間變化大于12.86%。未施氮條件下協青早B 的有效穗數和單株產量比施氮分別減少5.3 穗和6.6 g,每穗總粒數增加10.9;而未施氮下東鄉野生稻的有效穗數和單株產量卻較施氮分別增加了2.8 穗和3.5 g,每穗總粒數減少 7.7,說明氮素處理對有效穗數、每穗總粒數和單株產量等3 個性狀影響均較大。類似地,BIL群體有效穗數和單株產量平均值在不同處理間變化大于12.91%,其中施氮處理的有效穗數和單株產量分別較未施氮處理下提高了1.9 穗和2.4 g。雙親及BIL 群體的穗長、結實率、穗實粒數和著粒密度等性狀在兩種氮水平下雙親變化趨勢一致,但變異幅度不同。而雙親及其 BIL 群體的株高和千粒重在施氮和未施氮處理下的變化趨勢不同。這些結果表明,不同性狀對低氮脅迫的反應不盡相同。
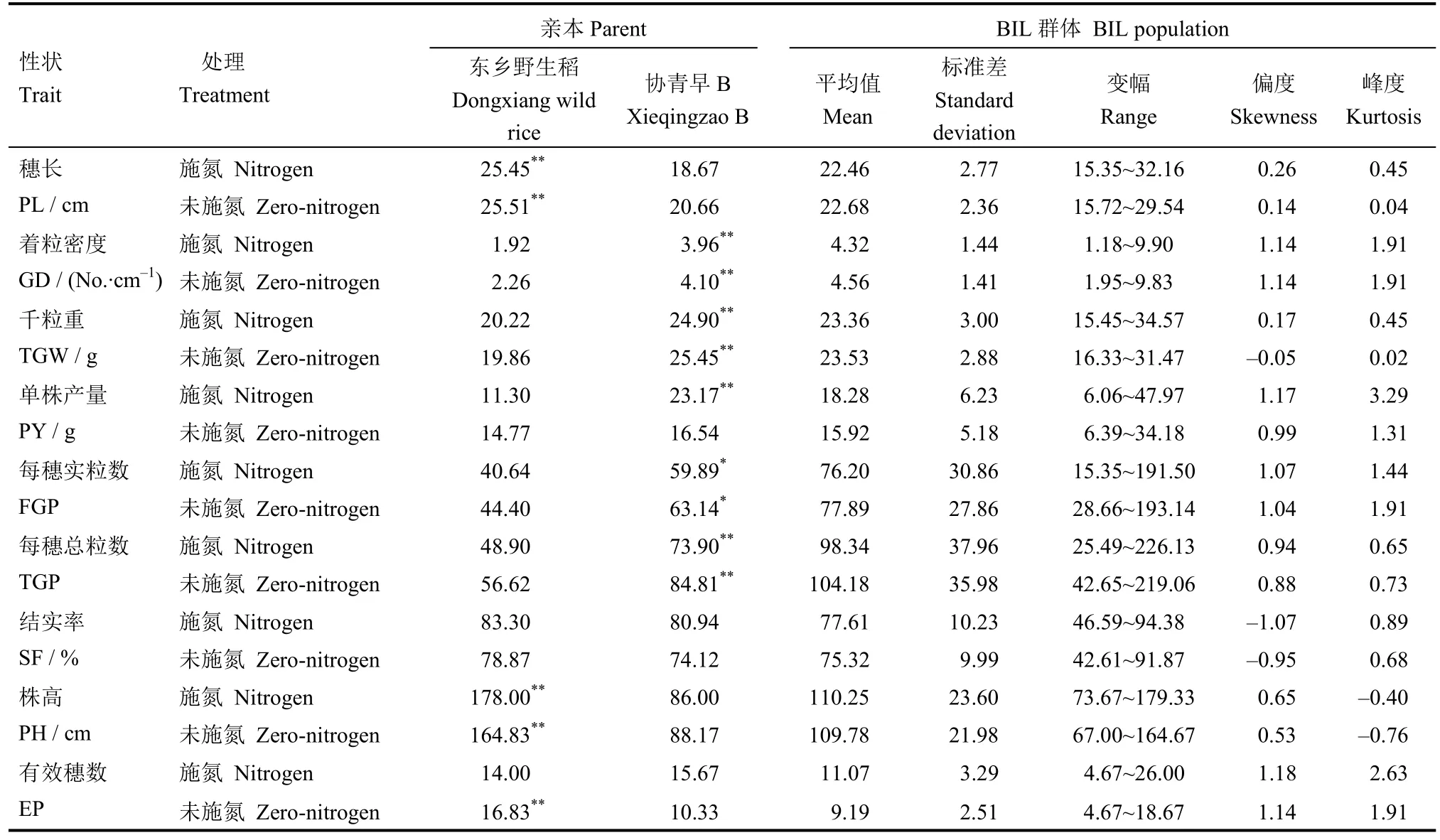
表1 施氮和未施氮條件下協青早B//東鄉野生稻/協青早B BIL 群體及親本的性狀表現Table 1. Phenotypic performance of the Xieqingzao B //Dongxiang wild rice/ Xieqingzao B BILs population and its parents under two nitrogen levels.
BIL 群體的9 個農藝性狀呈連續分布,除了著粒密度、有效穗數和單株產量的峰度值外,其余性狀的峰度值和偏度值均接近于 1,表現為典型的數量性狀遺傳,適合于QTL 分析。
2.2 兩種氮肥水平下性狀間相關性分析
相關性分析(表 2)表明,在兩種施氮處理下均呈極顯著正相關的有 15 對性狀,即每穗實粒數與株高、穗長、著粒密度、每穗總粒數、結實率和單株產量,每穗總粒數與穗長、著粒密度、單株產量和株高,穗長與株高、著粒密度和單株產量,單株產量與結實率和有效穗數,其中4 對(每穗實粒數與每穗總粒數、著粒密度和單株產量,穗總粒數與著粒密度)相關系數在兩種氮肥水平下均較大(0.63~0.96);呈極顯著負相關的有8 對性狀,即有效穗數與穗長、著粒密度、穗實粒數、穗總粒數和株高,千粒重與著粒密度、每穗總粒數和每穗實粒數,但相關系數較小(?0.19~?0.43);有3 對性狀間的相關性僅在未施氮條件下顯著或極顯著;其余10 對性狀間在兩種施氮處理下均不顯著。穗長、著粒密度、千粒重、單株產量、每穗實粒數、每穗總粒數、結實率、株高和有效穗數等9 個性狀在兩種氮肥水平間的相關性均達到極顯著,其相關系數分別為 0.87、0.94、0.91、0.63、0.88、0.92、0.72、0.38、0.61。由此可見,水稻的耐低氮特性可以通過這些產量性狀表示。
2.3 兩種氮肥水平環境下QTL 分析
在施氮和未施氮條件下共檢測到 57 個控制產量性狀的QTL(表3、圖1),分布于除第4 和第8 染色體外的所有染色體上,單個QTL 表型貢獻率為3.17%~63.40%。其中,19 個QTL 在施氮和未施氮條件下均檢測到,且加性效應方向保持不變;16和22 個QTL 分別僅在施氮和未施氮環境下檢測到顯著效應。
2.3.1 穗長
共檢測到 4 個控制穗長 QTL,表型貢獻率為6.66%~32.46%,其中qPL1、qPL5和qPL10在 2個氮肥環境下均被檢測到,qPL6僅在未施氮水平下檢測到。qPL1和qPL5的增效等位基因來自東鄉野生稻,而qPL6和qPL10的增效等位基因來自協青早B。除了qPL6的貢獻率(6.66%)小于10%外,其余3 個QTL 貢獻率(10.24%~32.46%)均大于10%,為主效QTL。
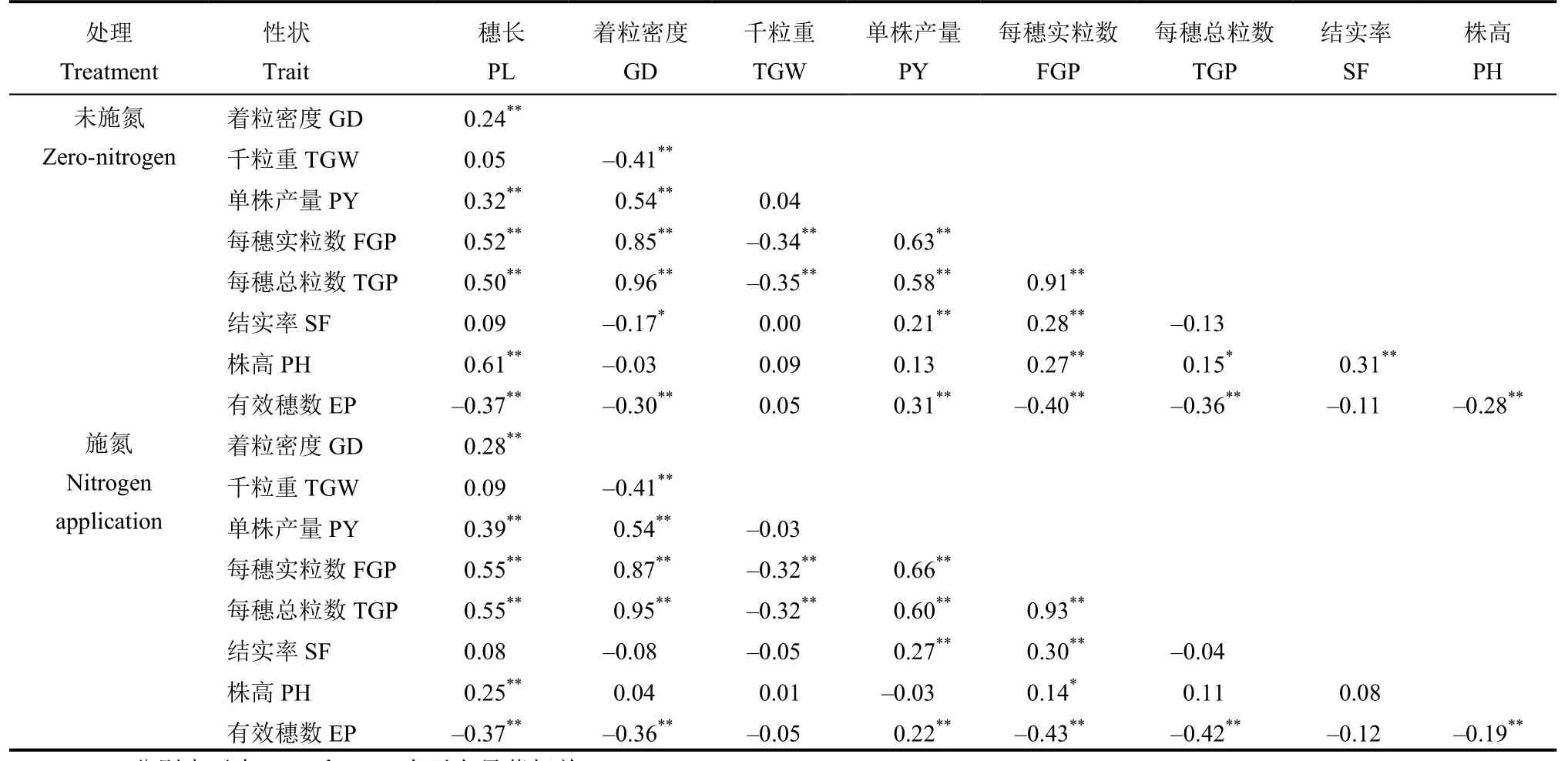
表2 兩種氮素條件下產量相關性狀間的相關性Table 2. Relationships between traits related to grain yield under two nitrogen conditions.
2.3.2 著粒密度
共檢測到 9 個控制著粒密度 QTL,其中qGD1.1、qGD1.2、qGD6、qGD9和qGD11等 5 個QTL 在兩種氮肥環境下均被檢測到,其余4 個僅在單個環境下均被檢測到。qGD1.1的增效等位基因來自東鄉野生稻,未施氮和施氮條件下的貢獻率分別為16.88%和12.60%。qGD9和qGD11在未施氮環境下表現為主效 QTL(其貢獻率分別為 19.52%和11.47%),而在施氮水平下表現為微效QTL(其貢獻率分別為5.41%和6.90%),其增效等位基因均來自東鄉野生稻。qGD3和qGD5僅在未施氮環境下檢測到,表型貢獻率分別為14.38%和18.79%,其增效等位基因均來自東鄉野生稻。剩余4 個QTL的表型貢獻率均較低(3.23%~5.67%),其中,qGD1.2、qGD2和qGD6的增效等位基因均來自協青早B,qGD12的增效等位基因均來自東鄉野生稻。
2.3.3 千粒重
共檢測到 7 個控制千粒重 QTL,分別解釋5.05%~10.36%的表型變異,其中qTGW7和qTGW10在2 個氮肥條件下均被檢測到,剩余5 個QTL 僅在單一環境下被檢測到。qTGW1、qTGW2和qTGW6等3 個 QTL 的增效等位基因均來自東鄉野生稻,而其余4 個來自協青早B。除了qTGW10在未施氮環境下的表型貢獻率為10.36%外,其余的表型貢獻率均較低。
2.3.4 單株產量
共檢測到 5 個控制單株產量 QTL,其中僅有qPY5.1均在 2 個施氮水平下被鑒定到,其余 4 個QTL 僅在單一施氮水平下檢測到。qPY5.1的表型解釋率在施氮和未施氮條件下分別為14.92%和9.21%,東鄉野生稻等位基因分別增加單株產量 2.29 g 和2.09 g。qPY5.2和qPY9.2為主效QTL,其增效等位基因來自東鄉野生稻;而剩余2 個QTL 為微效QTL,其增效等位基因來自協青早B。
2.3.5 每穗實粒數
共檢測到7 個控制每穗實粒數QTL,其中3 個QTL 在2 個施氮水平下均被鑒定到,剩余4 個QTL僅在單一環境下被鑒定到。qFGP1的增效等位基因來自東鄉野生稻,在施氮和未施氮水平下的表型貢獻率分別為23.08%和27.09%。qFGP5和qFGP9的增效等位基因均來自東鄉野生稻,其中在未施氮環境下檢測為主效 QTL(其貢獻率分別為 13.50%和21.89%),在施氮水平下檢測為微效QTL(其貢獻率分別為8.04%和5.28%)。qFGP11的表型貢獻率為10.10%,來自東鄉野生稻的等位基因增加12.49 粒;而其余3 個QTL 的表型貢獻率均較低,增效等位基因除qFGP10外都來自協青早B。
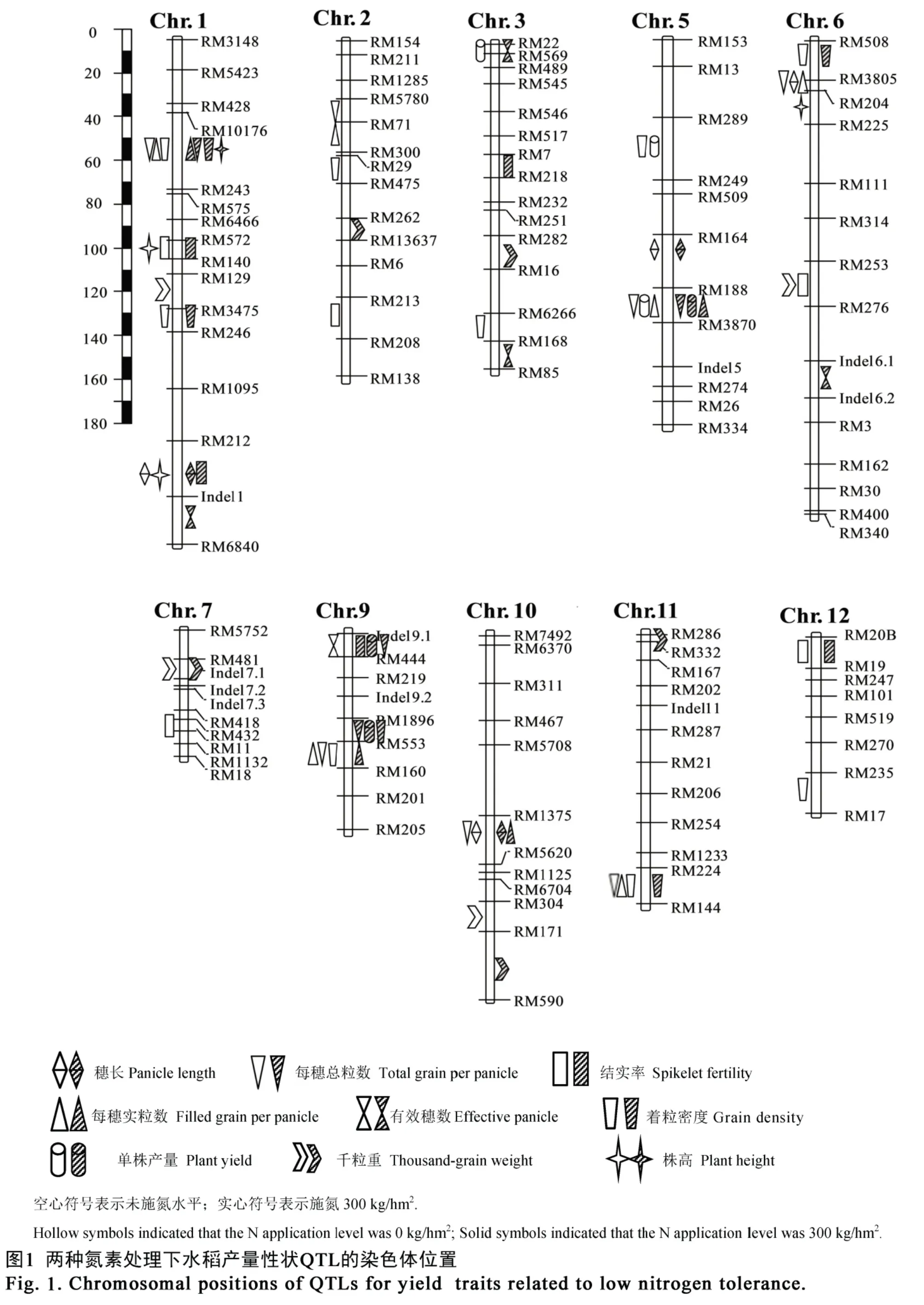
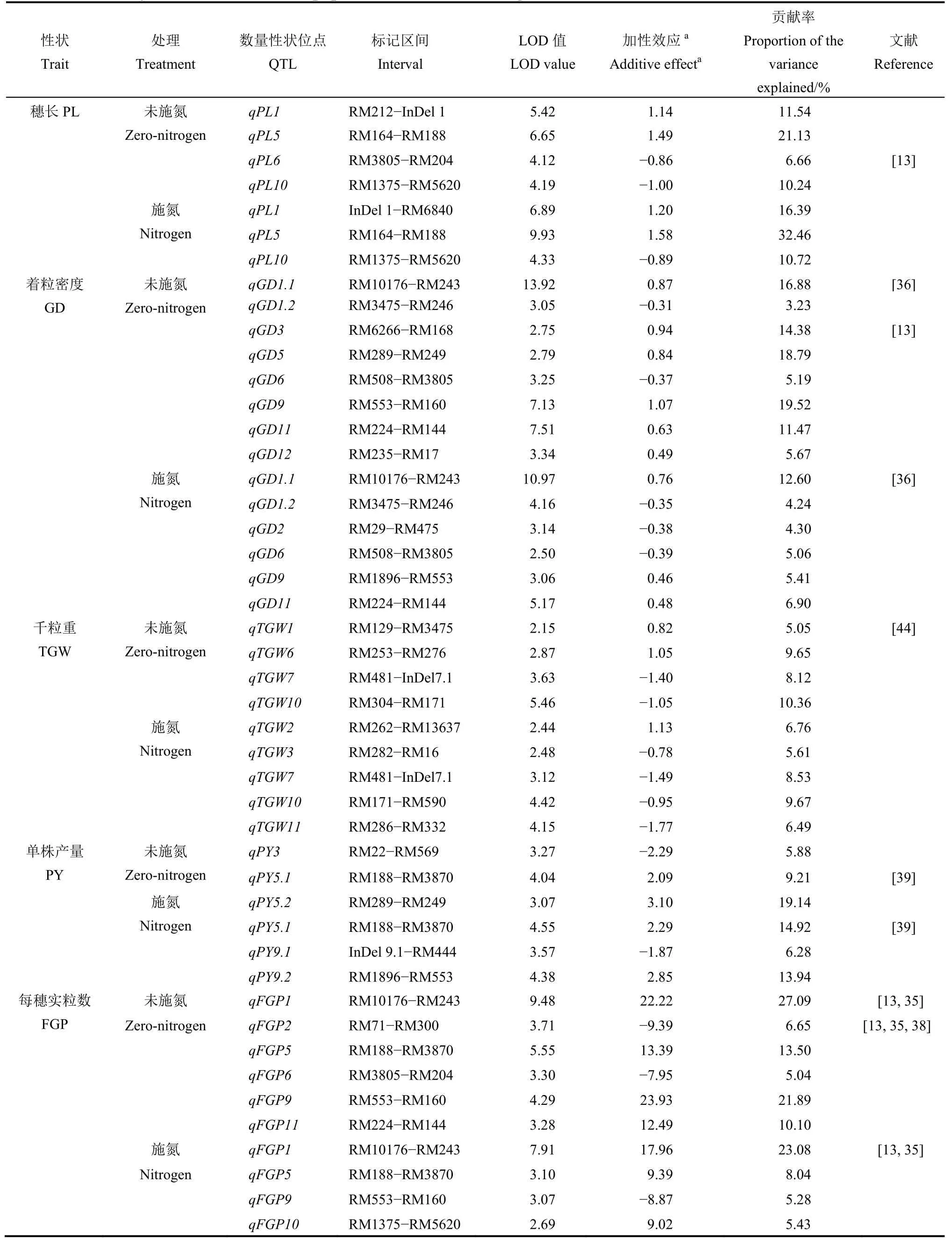
表3 兩個氮素條件下BIL 群體產量性狀的QTLTable 3. QTL for yield traits in the BILs population under two nitrogen conditions.
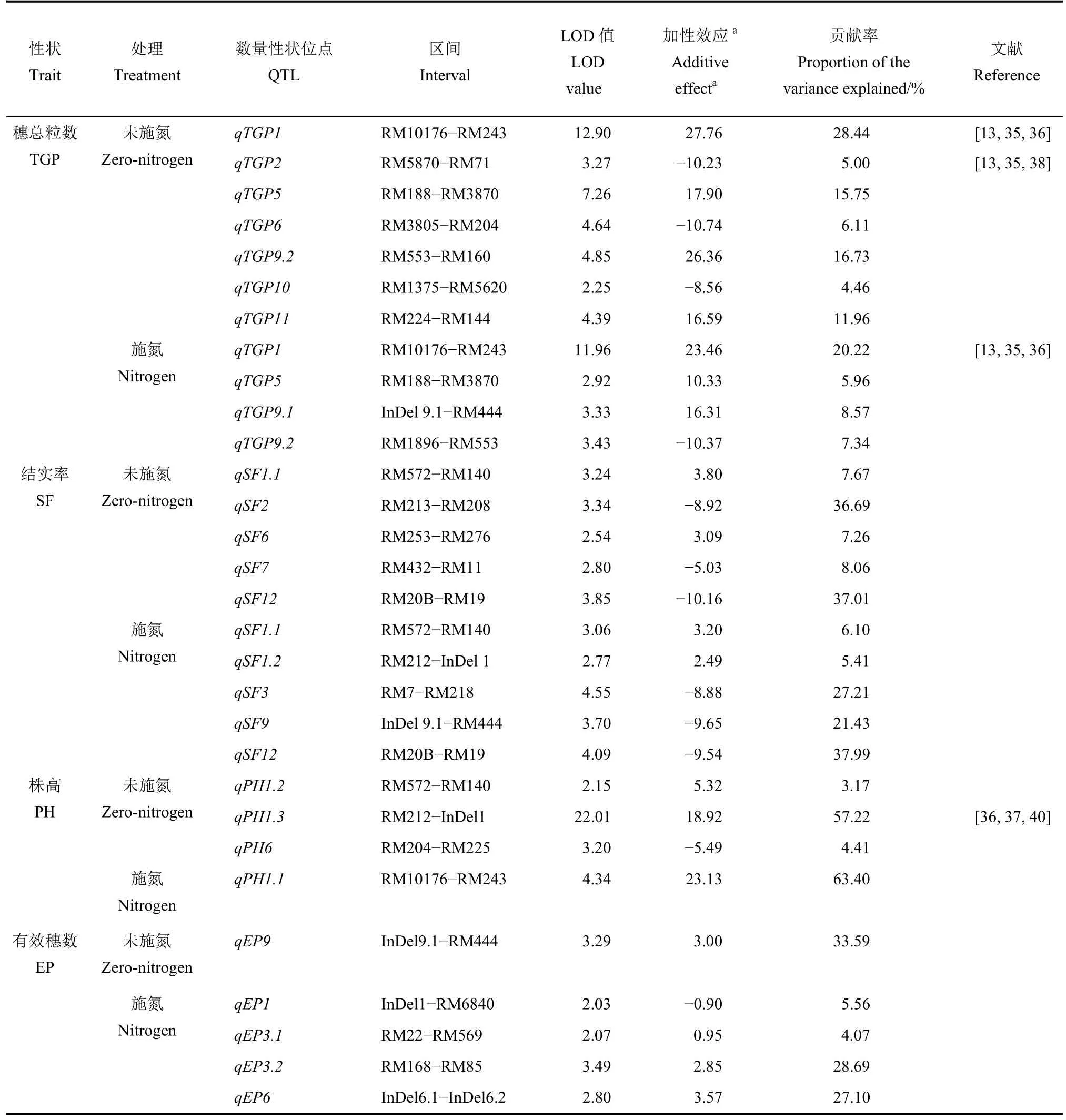
續表3 (Continued Table 3)
2.3.6 每穗總粒數
共檢測到8 個控制每穗總粒數QTL,其中3 個QTL 在2 個施氮水平下均被檢測到,剩余5 個QTL僅在單一環境下被檢測到。qTGP1的增效等位基因來自東鄉野生稻,表型貢獻率最高,在施氮和未施氮水平下分別為 20.22%和 28.44%。qTGP5和qTGP9.2的增效等位基因均來自東鄉野生稻,其中在未施氮環境下檢測為主效QTL(其貢獻率分別為15.75%和 16.73%),而在施氮水平下檢測為微效QTL(其貢獻率分別為5.96%和7.34%)。qTGP11僅在未施氮環境下鑒定到,表型貢獻率為11.96%,其東鄉野生稻等位基因可提高每穗總粒數16.59 粒。其余4 個QTL 的表型貢獻率均較低(4.46%~8.57%),且其增效等位基因均來自協青早B。
2.3.7 結實率
共檢測到8 個控制結實率QTL,其中2 個QTL在2 個施氮水平下均被檢測到,剩余6 個QTL 僅在單一環境下被檢測到。qSF12的表型貢獻率最高,在施氮和未施氮水平下分別為37.99%和37.01%,其協青早 B 等位基因分別提高結實率 9.54%和10.16%。qSF2、qSF3和qSF9等 3 個 QTL 的表型貢獻率分別為36.69%、27.21%和21.43%,其協青早B 增效等位基因分別提高結實率8.92%、8.88%和9.65%。其余4 個QTL 均為微效QTL,其中qSF1.1、qSF1.2和qSF6的增效等位基因均來自東鄉野生稻,而qSF7的增效等位基因來自協青早B。
2.3.8 株高
共檢測到4 個控制株高QTL,其中3 個在未施氮處理下檢測到,1 個在施氮水平下檢測到。qPH1.1和qPH1.3分別解釋表型變異的63.40%和57.22%,其東鄉野生稻等位基因分別增加株高 23.13 cm 和18.92 cm。剩余的2 個微效QTL 均在未施氮條件下檢測到,其中qPH1.2的增效等位基因來自東鄉野生稻,qPH6的增效等位基因來自協青早B。
2.3.9 有效穗數
共檢測到 5 個控制有效穗 QTL,貢獻率為4.07%~33.59%,其中4 個在施氮水平下檢測到,另外1 個未施氮水平下檢測到,未檢測到共同的QTL。qEP9的表型貢獻率最高,為33.59%,其增效等位基因來自東鄉野生稻。在施氮水平下檢測到的4 個QTL 中,qEP3.1、qEP3.2和qEP6東鄉野生稻增效等位基因均來自東鄉野生稻,加性效應分別為0.95、2.85 和3.57;而qEP1的協青早B 等位基因提高有效分蘗0.90 個。
3 討論
氮素作為水稻生長發育必需的大量元素之一,對水稻的產量起著至關重要的作用。而過量使用氮肥會造成水稻品質下降、氮肥利用率低、農業生態環境污染等問題。因此,選育水稻耐低氮品種對農業經濟和生態環境效益具有重要的現實意義。不同水稻品種在耐低氮的特性和氮素的吸收利用效率上存在顯著的基因型差異[6-8,34],這對研究水稻耐低氮或氮肥高效利用的分子機制及培育耐低氮水稻品種至關重要。由于傳統育種時間長且不確定性大,發掘鑒定水稻耐低氮相關基因,結合分子育種與傳統育種等手段,加快選育出耐低氮水稻新品種。
本研究利用東鄉野生稻與協青早B構建的回交重組自交系群體分析兩種氮肥條件下產量相關性狀,結果發現不同氮素水平下各株系的性狀間特別是有效穗數、每穗總粒數和單株產量間存在較大差異,而不同施氮水平各性狀呈極顯著正相關,這表明不同基因型株系以及不同性狀對氮素的響應存在一定差異,與前人報道[15]相似。QTL 分析結果共檢測到57 個產量相關的QTL,其中19 個QTL 在兩種氮肥水平下都檢測到,且加性效應方向保持不變;16 和22 個QTL 分別僅在施氮和未施氮環境下檢測到顯著效應。相似地,Cho 等[14]在高氮條件下和低氮條件下檢測到的QTL 有很大差別;馮躍等[13]在不同氮肥條件檢測到11 個相同的產量性狀QTL和30 個差異表達的產量性狀QTL;Tong 等[18]分別在高氮、中氮和低氮等3 種施氮水平下檢測到15、23 和 19 個 QTL,其中 5、3 和 5 對上位性 QTL 與環境存在顯著互作。這些結果表明控制水稻產量性狀的 QTL 在不同氮素環境下的差異表達,主要與施氮水平間的互作效應有關。
3.1 QTL 定位結果比較
近年來,已定位到大量耐低氮或氮素利用率相關QTL,其中有些在不同遺傳群體和環境中被穩定檢測到的QTL,對精細定位、克隆及分子育種非常重要。基于 QTL 連鎖標記的物理位置,比較分析不同研究中共同檢測到的耐低氮相關QTL,驗證本研究中 QTL 的準確性和可靠性。結果表明,在兩種氮素水平下和僅在低氮水平下被檢測到,且與前人報道相同性狀QTL 分別有4 個和6 個,由此共有31 個新檢測到的耐低氮相關QTL。其中在兩種氮肥水平檢測到的4 個QTL。位于第1 染色體上的3 個 QTL(qFGP1、qTGP1和qGD1.1)與qGD1.1、qFGP1.1、qSP1.1[13]、qGNS1[35]和qSPP-1[36]等相應的穗部性狀QTL 一致,該區間還報道了qGWP1和qPL1.1[13]、qHGW-1b[18]、qTLN-1[36]、qpn-1b和qph-1b[37]以及qPNA1[38]等不同性狀 QTL;在第 5染色體上的qPY5.1與ypp5b[39]的區間一致,在該區域附近還檢測到控制葉片干質量、莖干質量、總鮮質量、葉片數和劍葉氮含量等耐低氮性狀 QTL[10]。6 個僅在低氮水平下被檢測到的 QTL 如下:株高QTLqPH1.3與qPHT-1[36]、qph-1a[37]和qPH1[40]染色體區間相同;qTGW1與q1000Gwt-1.3和q1000Gwt-1.6[44],且該區間還報道了qby-1b[37];位于第 2 染色體的 2 個 QTL(qFGP2和qTGP2)與qFGP2、qSP2[13]、qGNR2a、qGNA2[38]、qGNS2.2[35]等氮素利用率相關 QTL 區間相同或部分重疊,該區間附近還檢測到qCc2c[18]、qSF2和qGNS2.1[35]以及qDWS2.2和qFGN2.4[41]等耐低氮相關性狀QTL;第 3 染色體上的控制著粒密度qGD3與qGD3.1[13]一致,該區間檢測到qTN-III_3[11]、qFGP3.2和qPNP3.1[13]以及qDTF-3和qPTN-3-2[36]等不同耐低氮QTL;qPL6與qPL6.1[13]的染色體區間一致。針對不同遺傳群體能穩定檢測到耐低氮性狀QTL 構建相應的近等基因系,開展相應的QTL等位性分析和分子克隆,最終實現水稻分子育種。
通過對不同耐低氮性狀 QTL 比較發現,本研究檢測到10 個耐低氮性狀QTL 染色體區間與已報道的不同耐低氮性狀QTL 區間相似。第1 染色體檢測到兩個氮水平下的qSF1.1附近區間報道了qsy1b和qtncs1b[14]等氮素利用相關QTL。在2 個氮素水平下均檢測到的穗長QTLqPL1和僅在低氮水平檢測到的qPH1.3與qFGP1.2、qGYn1[15]、qSNA1、qNHI1b[38]、qGY1[41]、qPN1和qGY1[43]等 QTL 染色體區間相同。第3染色體檢測到的qPY3與Senthilvel等[36]報道的qPTN-3-1染色體區間相同。第5 染色體檢測到 2 個氮素水平下的qPL5與qAGB5.2、qAGB5.3和qSVHS5.1[42]。在2 個氮素水平下均檢測到的qGD6以及僅在低氮水平下檢測到的 3 個QTL(qFGP6、qTGP6和qPH6)分別與qTN-II_6[11]、qPH-6b[12]、qpn-6[37]、qDWS6.1和qGY6.1[41]以及qRW-6-2[46]等不同耐低氮性狀QTL 一致。第7 染色體上檢測到控制兩個氮水平下的qTGW7與qHD-7[12]、qGYPP-7a[18]、qpn-7[37]、qNHI7a[38]、qFGN7.1[41]、qPSPF_7.2和qBY_7.1[44]以及葉綠素含量QTLqCc7a[45]等有相似區間。第9 染色體檢測到低氮環境下的qEP9與控制穗實粒數QTLqFGN9[37]一致;在兩個環境下檢測到的qGD9、qFGP9和qTGP9.2與已報道的qgy9[14]、qRGN和qRBN9[16]、qSL9[35]、qPTN-9[36]、qGNR9[38]、qPH9.2、qGY9、qDWS9和qDWR9[41]、qSW-9-3[46]以及控制莖干質量、根干質量和植株干質量QTL[47]等不同耐低氮相關性狀 QTL 的染色體區間相似。兩種施氮水平下的qTGW10區間與qFGPP-10b[18]、qGY10.2和qDWR10[41]、qFGN_10.1、qFGN_10.2和qPSPF_10.3[44]、qYd10[45]以及qBY10.1、qNUP10.1和qNUE10.2[48]等耐低氮性狀QTL 的染色體區間相同。控制兩種施氮環境下的qGD11和低氮環境下的qFGP11、qTGP11與已報道的qSSR11.2[35]、qGNR11b[38]、qGY11[41]以及qNUP11.2和qBY11.1[48]等耐低氮相關 QTL 的染色體區間一致。控制兩種施氮環境下的qSF12與控制氮肥生理利用率QTL[10]和qYd12[45]一致。
近年來,一批參與水稻氮素吸收調控基因相繼被成功分離克隆,為水稻耐低氮或氮肥高效利用機理和品種選育奠定良好的基礎。在低氮環境下的qSF2區間附近報道了一個水稻氯酸鹽抗性基因OsNR2,秈型OsNR2具有更高的硝酸還原酶活性和根部硝態氮吸收活性,提高水稻有效分蘗和產量[23]。水稻氮利用效率基因qNGR9[49]位于 QTL 簇qGD9/qFGP9/qTGP9.2內,編碼 Gγ 亞基,與 Gα 亞基 RGA1 和 Gβ 亞基 RGB1 互作,降低 RGA1 活性或提高RGB1 活性會抑制氮反應,該基因顯性突變造成植株對氮素不敏感和提高氮素的攝取和同化效率,進而仍能保持在低氮條件下獲得較高稻谷產量。硝酸鹽轉運蛋白NRT1.1B[26,27]位于qTGW10區間,其秈型等位基因增加分蘗數和谷粒產量來提高水稻氮素利用率。TOND1[24]位于檢測到低氮環境下的qGD12 區間內,可提高水稻苗期耐低氮能力及水稻稻谷產量。有必要構建協青早B 遺傳背景的氮素吸收調控基因位點差異化的近等基因系,進而分析這些基因與本研究鑒定到的QTL 的遺傳關系。
3.2 QTL 成簇分布
QTL 成簇分布在QTL 定位研究中普遍存在[9],可能是由于基因的多效性和多個基因的緊密連鎖造成的。類似地,本研究檢測到的 43 個產量相關QTL 分別聚集于7 條染色體上的14 個QTL 簇中,表明不同性狀可能涉及到共同遺傳機制。其中,位于第6 染色體上QTL 簇最大,包含qPL6、qGD6、qFGP6、qTGP6和qPH6等5 個QTL,其次是含4個QTL 的QTL 簇5 個,接下來依次為含3 個QTL的 QTL 簇 2 個和含 2 個 QTL 的 QTL 簇 6 個。其中,位于第 6 染色體上 QTL 簇qPL6/qGD6/qFGP6/qTGP6/qPH6均在未施氮條件下檢測到,其增效等位基因均來自協青早B,表明該QTL 簇對水稻耐低氮非常重要。由于初定位區間較大,一般難以探究QTL 簇的成因,因此有待于構建相關QTL 的近等因系將其分解到更小的區間內,從而解析 QTL 簇形成機理或 QTL 的一因多效機理;同時通過精細定位和克隆成簇的QTL,結合標記輔助選擇來實現水稻耐低氮QTL 分子聚合改良。
3.3 東鄉野生稻有利等位基因育種利用
東鄉野生稻棲息于氮素較低的貧瘠原生境,根系吸收氮素較少,通過氮素高利用效率才能生存。通過構建野栽高世代回交群體將東鄉野生稻耐低氮基因導入栽培稻,進而拓寬栽培稻遺傳基礎以及創制耐低氮水稻種質材料,開展“東鄉野生稻”耐低氮 QTL 定位、克隆以及分子育種。本研究檢測到的57 個QTL 中有32 個的增效等位基因來自東鄉野生稻,其中24 個QTL 東鄉野生稻等位基因與水稻耐低氮相關。針對穩定檢測到的耐低氮 QTL(如在兩種氮肥水平下均檢測到 QTL 簇qPL6/qGD6/qFGP6/qTGP6/qPH6)的有利等位基因進行精細定位,并結合分子標記輔助和 QTL 聚合方法,將東鄉野生稻耐低氮等位基因導入到協青早B 等栽培稻中,實現耐低氮水稻品種的遺傳改良。
猜你喜歡
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
礦山安全信息(2022年40期)2022-04-07 02:16:52
當代水產(2021年10期)2021-12-05 16:31:48
軍事文摘(2021年22期)2021-11-26 00:43:51
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
