日本無刺花椒抗寒性的初步研究
2020-11-23 07:34:42陳政李佩洪唐偉龔霞吳銀明吳軍
現代園藝·綜合版 2020年5期
陳政 李佩洪 唐偉 龔霞 吳銀明 吳軍
摘要:日本無刺花椒(ZanthoxyJum Japonica)為落葉或小灌木,是經濟價值高的重要調味品、香料和木本油料樹種。研究日本無刺枯椒的抗寒性,為其栽培種植提供依據。以朝倉山椒、葡萄山椒、琉錦山椒、花山椒4個種類花椒(Na)為試驗材料,對其進行了人工模擬低溫處理,測定了電解質外滲率(REC)、超氧化物歧化酶活性、過氧化物酶活性、丙二醛含量等抗寒性的生理生化指標并進行比較分析,結果表明:朝倉山椒和花山椒抗寒性較強,葡萄山椒抗寒性次之,琉錦山椒抗寒性最弱。
關鍵詞:日本無刺花椒;生理生化指標;抗寒性
花椒(Zanthoxylum bungeanum Maxim)原產于我國中西部,為蕓香科花椒屬落葉灌木或小喬木。全世界約有花椒屬植物250種,分布于亞洲、非洲、大洋洲、北美洲的熱帶和亞熱帶地區。中國是世界花椒第一生產大國,有39種14變種。四川是我國花椒主產區之一,漢源花椒、茂縣大紅袍花椒、金陽青花椒、洪雅藤椒、蓬溪青花椒是四川花椒中的五朵金花,是全國優質花椒之一。由于花椒樹干、枝、葉具刺,在育苗、栽植、管理和采收等方面有諸多不便,尤其采收難度大、成本高,花椒采收難是花椒生產中亟待解決的關鍵問題,若能以無刺或少刺花椒品種替代有刺花椒,將會大幅度提高采收效率、節約采收成本。為解決生產中花椒刺多的問題,我院于2009年從河北林業科學研究院引進花山椒、朝倉山椒、葡萄山椒和琉錦山椒等無刺花椒品種進行引種栽培試驗。日本無刺花椒(Zanthoxylum Japoni-ca),為落葉小灌木,雌雄異株,無刺或少刺。樹勢直立,萌芽力和成枝力均強。雌花具有雌蕊1-3枚;雄花具有雄蕊5-6枚,花粉量大。果實圓形或橢圓形,油腺點小且密,果皮鮮紅色。果皮、青果、嫩芽、種子等均有重要的應用價值。日本無刺花椒由于無刺或少刺,采摘和管理方便,較“茂汶花椒”“漢源花椒”“九葉青花椒”“藤椒”等花椒品種節約大量采摘用工和管理用工。日本無刺花椒在引種、育苗、栽培機理、修剪技術、施肥技術、風味物質及離體愈傷組織等方面做了大量工作,但對無刺花椒抗寒性研究報道較少,畢君對1a生枝條的細胞膜透性和SOD活性變化、POD活性變化、MDA含量變化、自然越冬的枝條電導率與凍害3種途徑進行抗寒評價,而紅花椒抗寒性研究報道較多。為此,筆者對無刺花椒材料的抗寒性進行初步研究。花椒抗寒性研究與農業生產密切相關。解決花椒寒害的途徑很多,其中選用耐寒的優良花椒品種是最經濟、也是最有效的途徑之一。因此,探索花椒抗寒性生理機制,不僅在理論上有重要意義,而且在生產中也具有廣泛的實用價值。
1試驗材料與方法
1.1試驗地概況
試驗地設在四川省資中縣明心寺鎮,位于29°44.29N,104°56.38E,年均氣溫17.4℃,極端最低氣溫-3.2℃:,極端最高氣溫41.9℃,年均日照時數1246.5h,年均降雨量1007.7mm,降雨量主要分布在5-9月。土壤為砂壤土,pH值7.0.海拔310m。
1.2試驗材料
測定的材料有朝倉山椒、葡萄山椒、琉錦山椒、花山椒,共4個品種,樹齡為2a。
1.2.1朝倉山椒。枝條密集,成抱頭狀生長,新梢上部綠色下部為棕色;樹皮光滑,基本無刺;葉片長寬均值為20.66mm/9.62mm,葉緣為鋸齒狀,向內卷曲,小葉數多為11-15枚;果實圓形,果實縱橫徑均值為4.85mm/4.46mm,臍部有一小突起,有殘留花柱,果皮鮮紅色。油腺點小、密而深陷。花期為3月中下旬,果期為8月中下旬。
1.2.2葡萄山椒。枝條較疏張,生長勢較弱;有皮刺;新梢上部綠色,下部為棕色;葉片長寬均值為24.66mm/12.28mm,葉緣為鋸齒狀,向內卷曲,小葉數多為9-P7枚;果實較大,果實縱橫徑均值為6.45mm/5.07mm,臍部有一小突起,有殘留花柱,果皮鮮紅色。油腺點小且密而深陷,花期為3月中下旬,果期為8月中下旬。
1.2.3琉璟山椒。樹姿較舒張,生長勢較強;無刺;新梢上部綠色,下部為棕色;
葉片長寬均值為21.27mm/10.17mm,葉緣為鋸齒狀,葉片平整,小葉數多9-15枚;果實較大,圓形,縱橫徑均值為5.94mm/5.01mm,臍部有一小突起,有殘留花柱,果皮鮮紅色。油腺點小、密而深陷。花期3月中下旬,果期為8月下旬-9月上旬。
1.2.4花山椒。樹勢強,枝條粗壯;無刺;葉片長寬均值為25.67mm/12.26mm,葉緣為鋸齒狀,向內卷曲,小葉數多為13-15枚;成花容易且花粉量極大,是比較理想的授粉品種。花期為3月上中旬。
1.3試驗方法
1.3.1植物材料處理。2019年4月,取樣時間為上午7:00-9:00.剪取健康、無病蟲害的朝倉山椒、葡萄山椒、琉錦山椒、花山椒花椒幼樹樹冠中部外側的4個方位的夏梢頂部向下的第4葉、第5葉(完整葉),將葉片用潔凈的濕紗布擦拭并晾干,按品種、處理分裝于塑料袋中,放人冰箱內進行人工冷凍,處理溫度分別為-6、-4、-2、0、2和4℃并保持恒溫5h。隨后,將各處理葉片放人-20%超低溫冰箱中保存,用于測定SOD活性、POD活性、MDA含量生理指標,設3個重復。
2019年1月上旬在資中試驗地內,選取朝倉山椒、葡萄山椒、琉錦山椒、花山椒4種花椒的健康、無病蟲害的1a生枝條作為供試材料。將各花椒的枝條用潔凈的濕紗布擦拭并晾干,放人冰箱內從0℃開始降溫,降溫速率為4℃/h,處理溫度分別為:-4、-8、-12、-16、-20、-24、-28℃,并保掙叵溫12h。再以同樣的速率升溫至0℃,取出枝條用于REC的測定,以放置室溫下(約為0℃)的枝條為對照(CK),設3個重復。
1.3.2指標測定方法。過氧化物酶(POD)活性測定采用愈創木酚法,超氧化物歧化酶(sOD)活性測定采用NBT(氮藍四唑)光化還原法,丙二醛(MDA)含量測定采用硫代巴比妥酸法,電解質外滲率采用電導率法。數據取各生理指標3次測定結果的平均值。以上測定方法均參考文獻。
1.3.3數據處理。數據處理用EXCEL和DPS(v6.85版)軟件完成。
2 結果與分析
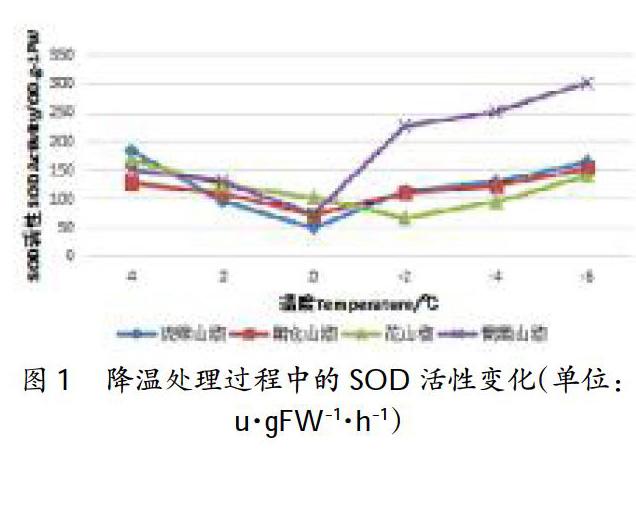
2.1SOD酶活性的變化
在正常條件下,植物細胞內自由基的產生與消除處于平衡狀態,這樣可以保持細胞膜系統的穩定性,維持正常的生理代謝活動。當植物處于各種逆境脅迫時,這種平衡會受到破壞而出現自由基的累積,從而引發或加劇細胞膜脂過氧化作用,造成膜系統受損,嚴重時導致細胞死亡。在低溫逆境條件下,SOD能被活性氧誘導產生,從而減輕對細胞膜的傷害。SOD、POD是植物體內清除自由基的主要酶類,其活性大小反映細胞對逆境的適應能力,也間接表明植物耐寒性的強弱。
植物在低溫脅迫初期,在植物細胞內,SOD主要起清除超氧自由基的作用。圖1所示,各花椒SOD活性隨著低溫脅迫的加強顯示出了先下降后增加的變化趨勢。隨著低溫的加深,4個山椒品系都是從4℃降到0℃時SOD活性下降,從0℃降到-6℃時上升,SOD活性變化呈“V”趨勢。SOD活性上升速率并不均勻,在0℃降到-2℃時SOD活性上升較快,其中葡萄山椒SOD活性上升最快,琉錦山椒次之,朝倉山椒SOD活性上升最慢。花山椒SOD活性的變化趨勢不同,花山椒4℃降到-2℃時SOD活性下降,從-2℃降到-6℃時呈現上升趨勢,活性變化呈“v”趨勢。可見,從SOD活性上升速率來看,葡萄山椒的上升速率最快,琉錦山椒的較快,朝倉山椒的次之,花山椒的最慢。
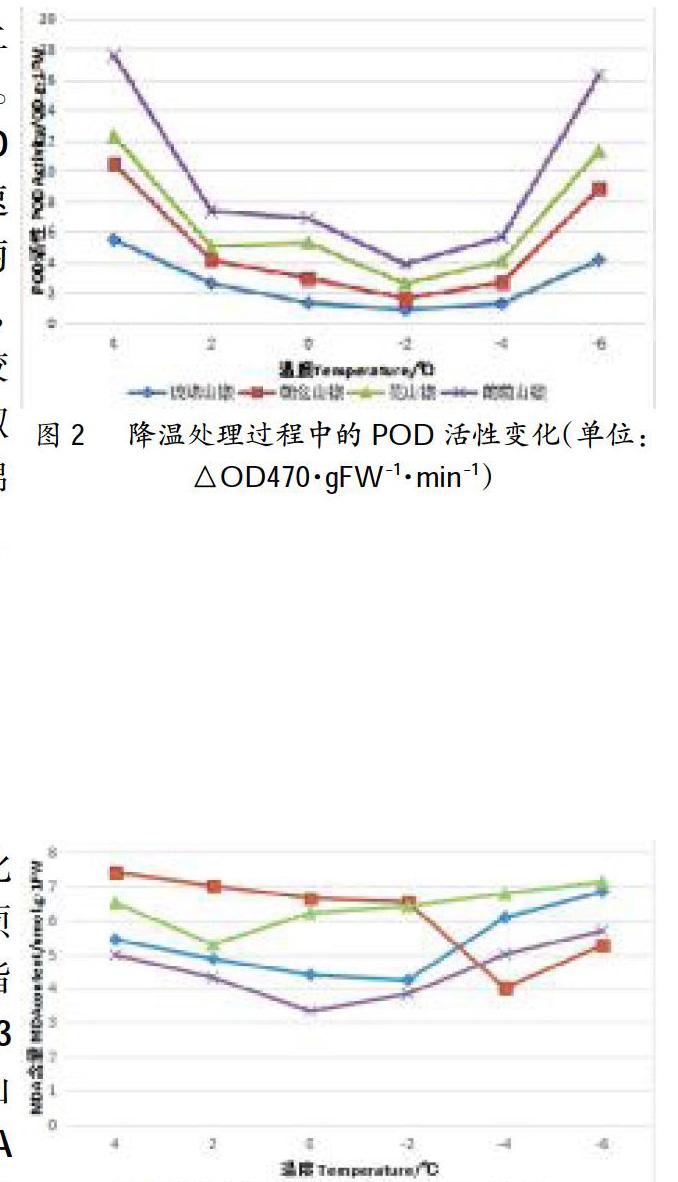
2.2POD酶活性的變化
過氧化物酶(POD)是植物體內普遍存在的活性較高的一種酶,它能夠清除超氧自由基形成H2O2,從而保護植物免受活性氧傷害。圖2所示,4個山椒品系POD活性隨著低溫脅迫的加強顯示出了花山椒先下降后增加后又下降再增加的變化趨勢。隨著低溫的加深,都是從4℃降到2℃時POD活性下降,從2℃降到0℃時上升,0℃降到-2℃時呈現下降趨勢,從-2℃降到-6℃時又呈現上升趨勢,活性變化呈“W”趨勢。朝倉山椒、琉錦山椒和葡萄山椒從4℃降到-2℃時下降,從-2℃降到-6℃時上升,活性變化呈“v”趨勢,POD活性上升速率不均勻,4個山椒品系在-2℃降到-4℃時上升較慢,在-4℃降到-6℃時上升較快,葡萄山椒POD活性上升速率最快。可見,從POD活性上升速率來看,葡萄山椒的最快,花山椒的較快,朝倉山椒的次之,琉錦山椒的最慢。
2.3MDA酶含量的變化
植物在逆境條件下往往發生膜脂過氧化作用,其產物MDA會嚴重損傷生物膜。通常利用它作為膜脂過氧化指標,表示細胞膜脂過氧化程度及對逆境反映的強弱,因此在研究植物寒害及抗寒性時,測定MDA含量的變化可作為一項生理鑒定指標。從圖3可知,4個山椒品系MDA含量隨著低溫脅迫的加強大多顯示出先下降后上升的趨勢。各山椒品系MDA含量上升溫度不同,增幅也不同。花山椒MDA含量從2℃開始上升,2℃降到0℃時其MDA含量上升了0.926umol/g.FW,隨后增幅變緩;葡萄山椒MDA含量從0℃開始上升,0℃降到-2℃時其MDA含量上升了0.504umol/g.FW,-2℃到-4℃時MDA含量上升了1.162umol/g.FW,隨后增幅變緩;琉錦山椒MDA含量從-2℃開始上升,-2℃降到-4℃時其MDA含量上升了1.843umol/g.FW,-4℃降到-6℃時其MDA含量上升了0.754umol/g.FW;朝倉山椒MDA含量從-4℃開始上升,-4℃降到-6℃時其MDA含量上升了1.090u-mol/g.FW。可見,琉錦山椒MDA含量增幅最大,葡萄山椒的次之,朝倉山椒的再次之,花山椒的最小。
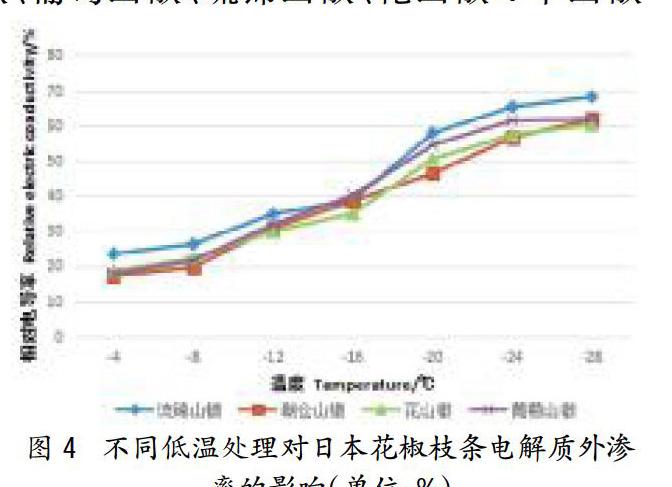
2.4不同低溫處理對花椒枝條質膜透性的影響
朝倉山椒、葡萄山椒、琉錦山椒、花山椒4個山椒品系1a生枝條經低溫處理后,其REC如圖4所示。從圖4可以看出,在一系列冷凍低溫處理下,4種花椒的REC與處理溫度之間關系呈上升的“s”形曲線。但其REC增加的速率并不均勻,處理溫度越低REC變化相對越快,且因種類而異。當溫度高于-16%時,隨溫度降低,枝條的相對電導率緩慢上升,-16~-24℃期間,相對電導率急劇增加,之后趨向平緩。-16℃下降至-20℃時,4個山椒品系相對電導率變化為:琉錦山椒的相對電導率增幅最大,為19.2%;花山椒的相對電導率增幅次之,為15.5%;葡萄山椒的為14.3%;朝倉山椒的相對電導率增幅最小,僅為7.9%。相對電導率急劇增大,說明枝條組織已經受到嚴重傷害,膜透性增加,細胞內電解質大量外滲。由此可知,在相同低溫下,琉錦山椒電解質外滲最多,脂膜損傷程度最重;葡萄山椒與花山椒電解質外滲較少,朝倉山椒電解質外滲最少,脂膜損傷程度較輕,這表明不同花椒種類的抗寒性存在差別。
3 結論與討論
(1)正常情況下,細胞內活性氧與防御系統之間保持著平衡。當遭遇低溫時,活性氧代謝失調,細胞內的活性氧就會積聚,從而對細胞的正常結構與功能起破壞作用,這時細胞內抗氧化防御系統被誘導和激活,以消除活性氧對細胞的損害。試驗結果表明,隨著溫度的下降,SOD和POD和活性呈先下降后升高的變化趨勢,這與劉玲等在花椒的研究結果一致,與畢君在山椒的研究結果存在一定差異,這可能是低溫處理方法不同所致。SOD和POD活性升高,這是4個山椒枝條對低溫脅迫所產生的保護性應激反應。
(2)丙二醛含量與植物抗寒性密切相關,其含量變化可反映植物對低溫條件反應的強弱,抗寒性強的品系枝條含量增幅較小,而抗寒性弱的品系枝條含量增幅較大。本試驗中,琉錦山椒MDA含量增幅最大,葡萄山椒的次之,朝倉山椒的再次之,花山椒的最小。說明琉錦山椒的抗寒性最弱,葡萄山椒的抗寒性較弱,朝倉山椒和花山椒的抗寒性較強。
(3)植物遭受低溫脅迫,生物膜的結構受到破壞,膜透性增大,從而引起電解質外滲,導致相對電導率的增大。因此,相對電導率大小能反映細胞質膜的損傷程度。細胞質膜透性大小與植物的抗寒性密切相關,同樣低溫條件下,細胞質膜透性大,電導率高且上升速度快,表明該植物品種抗寒性差,反之,則抗寒能力強。本試驗中,當處理溫度降低到-16℃,枝條的相對電導率出現了一個快速上升階段,表明枝條受到了嚴重傷害,細胞膜透性發生了不可逆的變化。在-16℃降到-20℃時,琉錦山椒電解質外滲最多,脂膜損傷程度最重;葡萄山椒與花山椒電解質外滲較少,朝倉山椒電解質外滲最少。結果表明:朝倉山椒和花山椒抗寒性較強,葡萄山椒次之,琉錦山椒最弱。測定的4個無刺花椒與畢君的研究結果存在一定差異,這可能是低溫處理方法不同所致。
