南亞熱帶混交人工林樹種豐富度與土壤微生物多樣性和群落組成的關系
2020-12-14 09:26:42宋戰超劉世榮胡佳佳明安剛李朝英伍俊廷史作民盧立華蔡道雄
生態學報 2020年22期
關鍵詞:研究
宋戰超,王 暉,*,劉世榮,胡佳佳,明安剛,陳 海,李朝英,伍俊廷,史作民,盧立華,蔡道雄
1 中國林業科學研究院森林生態環境與保護研究所,國家林業和草原局森林生態環境重點實驗室, 北京 1000912 中國林業科學研究院熱帶林業實驗中心,廣西友誼關森林生態系統國家定位觀測研究站, 憑祥 532600
土壤微生物在森林生態系統中發揮著不可替代的重要作用,積極參與到森林生態系統養分轉化和循環[1],對維持森林生態系統穩定和功能以及植物多樣性至關重要[2]。森林土壤微生物群落結構和多樣性受林下凋落物性質和土壤根系分泌物等生物因子及土壤環境等非生物因子的共同影響[3- 5]。同時,土壤微生物通過對土壤理化性質和土壤肥力作用,影響森林植被生物量和生產力[6]。
近年來,研究生態系統地上和地下生物多樣性之間的聯系一直是生態學研究的熱點。針對土壤微生物群落與植物群落關系的研究越來越多,但多以草地生態系統或天然林生態系統為主。Zak等[7]研究了植物多樣性對土壤微生物的影響,發現隨著植物多樣性的增加,土壤真菌生物量明顯增加。青藏高原天然草原的研究發現,隨著植物多樣性的增加,真菌α和β多樣性分別相應增加[8]。Chen等[9]在我國亞熱帶森林的不同樹種多樣性梯度實驗研究顯示,樹種特性是主導土壤細菌和真菌群落多樣性和結構變化主要因素,且對土壤真菌的影響比土壤細菌效果更強,而樹種多樣性并非是主要影響因子。基于我國亞熱帶森林生物多樣性與生態系統功能實驗,發現了不同功能型土壤真菌驅動亞熱帶森林群落多樣性的作用模式[10]。有關生物多樣性與生態系統功能關系的研究表明,生物多樣性是生態系統功能的主要驅動力[11]。不同樹種在森林群落中占據不同的高度和空間從而創造了更多的生態位,促進了群落利用陽光等環境資源的能力,提高了森林生態系統生物量和生產力。而我國人工林以單一樹種種植為主,由于生物多樣性下降,森林各種功能與生產力得不到充分發揮,降低了森林適應能力和穩定性,還會造成地力衰退、森林系統功能退化等問題[12]。如在我國南方地區大面積分布的桉樹人工純林,叢枝菌根真菌的種豐度隨種植年份的增加逐漸下降,優勢種Funneliformsgeosporus的豐度在2年生和10年生林地中顯著降低[13]。但是,樹種多樣性增加使森林植被組成和生態系統結構、功能趨于復雜,更多樹種混交后樹種多樣性會隨之增加以及樹種功能特性發生重疊,不同氣候區、不同類型森林生態系統植物種類和環境等條件差異也比較大,因此尚需通過研究進一步增強對森林植物多樣性與土壤微生物多樣性關系以及潛在機制的認知,進而提高有關土壤微生物對“樹種多樣性與土壤理化性質和肥力”關系影響機制的理解。
亞熱帶森林生態系統是我國陸地碳匯潛力的主要來源[14]。我國南方亞熱帶地區樹種多樣性豐富,人工林的面積和蓄積均居全國之首。Huang等[15]通過野外多樹種新造林實驗研究發現隨樹種多樣性增加林分生產力顯著增加。本研究面向人工林生態系統固碳增匯功能提升的國家需求,針對土壤微生物群落組成與樹種多樣性關聯機制這一關鍵科學問題,選取我國南亞熱帶經近40年近自然管理形成的多樹種、多層次、天然更新良好的異齡混交人工林生態系統為對象,采用高通量測序技術研究隨樹種豐富度變化土壤微生物α多樣性和群落組成的變化,以及主要的生物和環境驅動因子,旨在探討多樹種混交如何對南亞熱帶人工林土壤微生物生物量和群落結構組成產生影響,為進一步揭示樹種多樣性與地下土壤微生物多樣性關系提供科學依據,并為制定該區域人工林通過樹種豐富度合理組配調控提升生態系統功能的經營策略提供科技支撐。
1 材料和方法
1.1 研究區概況與實驗設計
研究區位于廣西壯族自治區憑祥市中國林業科學研究院熱帶林業實驗中心國家林業和草原局廣西友誼關森林生態系統定位研究站(22°06′50″—22°07′27″N,106°46′48″—106°47′22″E)。該地區屬典型南亞熱帶季風氣候區,海拔約300 m,年平均氣溫20.5—21.7℃,年平均降雨量1200—1500 mm,多集中在4—9月份。主要地貌類型以低山丘陵為主,土壤以磚紅壤和紅壤為主,成土母巖主要有泥巖夾砂巖、礫狀灰巖和石灰巖等。本研究的試驗區在杉木林采伐跡地,自1979年選取21個鄉土樹種以塊狀混交布局建立的試驗林,經過近40年的人工經營和天然更新,試驗區已形成多樹種、復層結構的混交人工林,試驗區面積100 hm2。
在試驗區內隨機設置54個20 m × 20 m樣地,并對每個樣地內胸徑(Diameter at breast height,DBH)≥1 cm林木的胸徑、樹高和坐標進行每木調查。依據每個樣地內的樹種豐富度將54個樣地劃分成10組,形成由1—10個樹種的豐富度梯度。其中第一組(1個樹種)有7個樣地,第二組(2個樹種)有7個樣地,第三組(3個樹種)有3個樣地,第四組(4個樹種)有4個樣地,第五組(5個樹種)有4個樣地,第六組(6個樹種)有7個樣地,第七組(7個樹種)有5個樣地,第八組(8個樹種)有10個樣地,第九組(9個樹種)有3個樣地,第十組(10個樹種)有4個樣地,各樣地的樹種組成詳見附件7。
1.2 樣品采集與處理
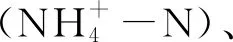
1.3 土壤理化性質、細根生物量和碳化學組分的測定
土壤pH值采用FE20 pH計測定,土水比為1∶2.5。采用重鉻酸鉀外加熱法和凱氏定氮法分別測定土壤有機碳和全氮含量[16]。采用鹽酸和硫酸萃取后分光度法測定土壤有效磷含量[17]。采用2 mol/L KCl浸提,靛酚藍比色法測定銨態氮,紫外分光光度法測定硝態氮[18]。
土樣過2 mm篩時,挑選出直徑 ≤ 2 mm的細根活體,用水反復沖洗去除附著在根系表面的泥土后,放置烘箱用65℃烘干至恒重,用電子天平稱重(精確到0.001 g),依據如下公式計算各樣地的細根生物量:
細根生物量(g/m2)=平均每個土芯中細根干重(g)/[π(Φ/2)2× (m2/104cm2)](Φ=5.0 cm)
采用13C固體核磁共振波譜(13C CPMAS NMR)分析土壤有機碳不同化學組分比例。13C固體核磁共振波譜被分為四組化學位移區域,以此區分土壤有機碳的不同組分:烷基碳(δ=0—45,alkyl C),氧烷基碳(δ=45—110,O-alkyl C),芳香碳(δ=110—160,aromatic C),羰基碳(δ=160—210,carbonyl C)。
1.4 土壤微生物總DNA提取和高通量測定
稱取約0.25 g經冷凍干燥處理的土壤樣品,使用MO BIO PowerSoil(MO BIO Laboratories,Carlsbad,USA)試劑盒提取土壤微生物總DNA,采用NanoDrop分光光度計進行定量提取DNA的質量和濃度,并用1%瓊脂糖凝膠電泳對提取的DNA質量進行檢測,合格后用于構建文庫;以各土壤樣品微生物總DNA為模板,以細菌V4和V5區特異性引物(15F 5′-GTGCCAGCMGCCGCGGTAA- 3′;926R 5′-CCGTCAATTCMTTTGAGTTT- 3′)[19]和真菌ITS1區特異性引物(ITS1F 5′-CTTGGTCATTTAGAGGAAGTAA- 3′;2043R 5′-GCTGCGTTCTTCATC GATGC- 3′)[20]進行PCR(polymerase chain reaction)擴增;采用Illumina MiSeq測序平臺對PCR擴增產物進行雙端測序分析,微基生物科技(上海)有限公司完成測序。
土壤微生物目標片段的PCR產物測序完成后,對有效序列進行質控、拼接和精確去雜等處理,得到優化序列。優質序列利用Usearch軟件基于97%的相似水平進行分類操作單元(operational taxonomic units,簡稱OTUs)。根據Silva細菌數據庫[21]和Unite[22]真菌數據庫利用 RDP Classifier進行物種注釋和分類。利用QIIME(Version l.7.0)軟件依據樣本文庫的OTUs豐度信息計算土壤樣品的細菌和真菌多樣性(Faith′s Phylogenetic Diversity,Faith′s PD指數)。利用R語言軟件VEGAN包對土壤細菌和真菌β多樣性進行了距離冗余分析(Distance-based redundancy analyses,db-RDA),并篩選主要解釋變量。
1.5 統計分析
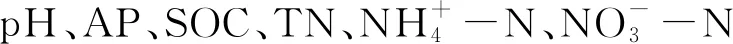
2 結果與分析
2.1 樹種豐富度與土壤細菌和真菌α多樣性的關系
通過對本研究中的人工林樹種豐富度梯度序列樣地建立不同樹種豐富度梯度(1—10個樹種)與土壤細菌α多樣性之間的相關關系,樹種豐富度與土壤細菌α多樣性的關系如圖1所示。Pearson相關分析的結果表明,土壤細菌α多樣性隨樹種豐富度增加整體間存在增加的趨勢,但影響不顯著(P>0.05)。通過對土壤細菌α多樣性和土壤理化性質關系的分析,發現土壤細菌α多樣性與土壤pH和芳香碳組分呈顯著正相關(P< 0.05),而與有機碳、銨態氮、碳氮比、烷基碳組分及細根生物量呈顯著負相關(P<0.05)。采用逐步回歸分析揭示土壤理化性質與土壤細菌α多樣性之間的關系,結果表明土壤pH和銨態氮是土壤細菌α多樣性變化的主要影響因素(表1)。
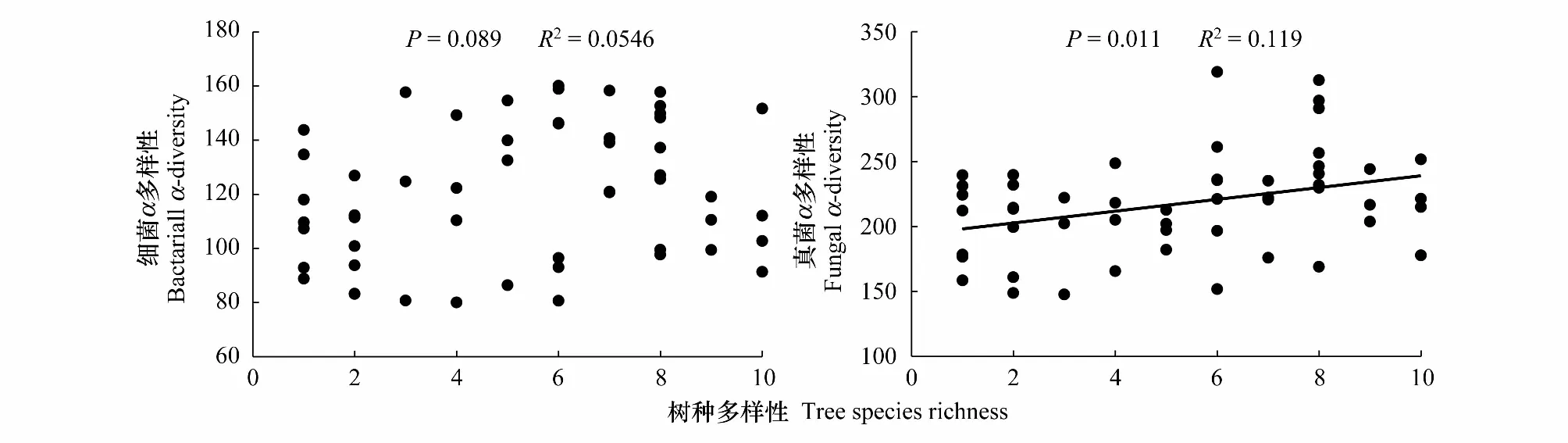
圖1 樹種豐富度與土壤細菌α多樣性和真菌α多樣性的關系Fig.1 Relationship of tree species diversity on soil bacterial α-diversity and fungal α-diversity
樹種豐富度與土壤真菌α多樣性的關系如圖1所示。Pearson相關分析的結果表明,土壤真菌α多樣性與樹種豐富度之間存在顯著正相關關系(P<0.05),隨樹種豐富度增加,土壤真菌α多樣性顯著提高。通過對土壤細菌α多樣性和土壤理化性質關系的分析,發現土壤真菌α多樣性與土壤pH和碳氮比呈顯著正相關(P<0.05),而與細根生物量呈顯著負相關(P< 0.05)。逐步回歸分析表明土壤pH和有效磷是土壤真菌α多樣性變化的主要影響因素(表1)。

表1 土壤細菌和真菌α多樣性與環境因子的多元統計結果
2.2 不同樹種豐富度土壤細菌和真菌群落組成變化
對不同樹種豐富度土壤微生物群落組成進行PerMANOVA分析,結果發現土壤細菌β多樣性在不同樹種豐富度之間存在顯著差異(表2)。對54個土壤樣品進行Illumina MiSeq高通量測序,數據經過濾優化,共得到1964785個細菌優化序列,平均每個樣品36385個序列,共得到4769個OTUs。在綱水平上土壤細菌群落共鑒定48綱,相對豐度大于1%的綱共15個。樣品中土壤細菌群落主要由酸桿菌綱(13.19%)、α變形菌綱(13.06%)、γ變形菌綱(9.85%)、β變形菌綱(6.00%)和δ變形菌綱(5.61%)組成,其他微生物種類以低豐度(小于5.00%)存在,如:纖線桿菌綱(3.45%)、浮霉菌綱(0.80%)。樹種豐富度增加變化改變了土壤細菌β多樣性,在不同樹種豐富度梯度下α變形菌綱、迷蹤菌綱、γ變形菌綱、浮霉菌綱、Ignavibacteria(未命名)和螺旋體綱差異顯著(P<0.05)(圖2)。

圖2 不同樹種豐富度與土壤細菌群落和真菌群落組成變化Fig.2 Diversity of different tree species and composition of soil bacterial community and fungal community 細菌群落中Aα變形菌綱;G:γ變形菌綱;P:浮霉菌綱;E:迷蹤菌綱;I:Ignavibacteria,未命名;S:螺旋體綱;真菌群落中A:傘菌綱;L:茶漬綱;T:銀耳綱;**表示顯著水平P< 0.01,*表示顯著水平P< 0.05

表2 不同樹種豐富度梯度下土壤細菌和真菌群落PerMANOVA分析
在不同樹種豐富度梯度下土壤真菌β多樣性存在顯著差異(表2)。真菌測序共得到2068537個優化序列,聚集在8124個OTUs中,平均每個樣品38306個序列。在綱水平上土壤真菌群落共鑒定34綱,相對豐度大于1%的綱共8個。樣品中土壤真菌群落主要由傘菌綱(23.38%)組成,其他種群豐度相對較低,如茶漬綱(3.07%)、銀耳綱(2.84%)。土壤真菌群落中的傘菌綱、茶漬綱和銀耳綱在不同樹種豐富度梯度下差異顯著(P<0.05)(圖2)。
2.3 環境變量對土壤細菌和真菌群落組成的影響
采用db-RDA分析來評估環境變量對土壤細菌群落組成的影響。分析結果顯示,土壤細菌群落在第一主軸(橫軸)主要由土壤pH和芳香碳組分貢獻,累計解釋變量達到92.86%。第二主軸(縱軸)累計解釋變量為5.81%,主要由土壤硝態氮貢獻。土壤細菌群落組成與土壤環境因子密切相關,土壤pH、硝態氮和芳香碳組分是樹種豐富度梯度間土壤細菌群落組成差異的主要影響因素(圖3)。
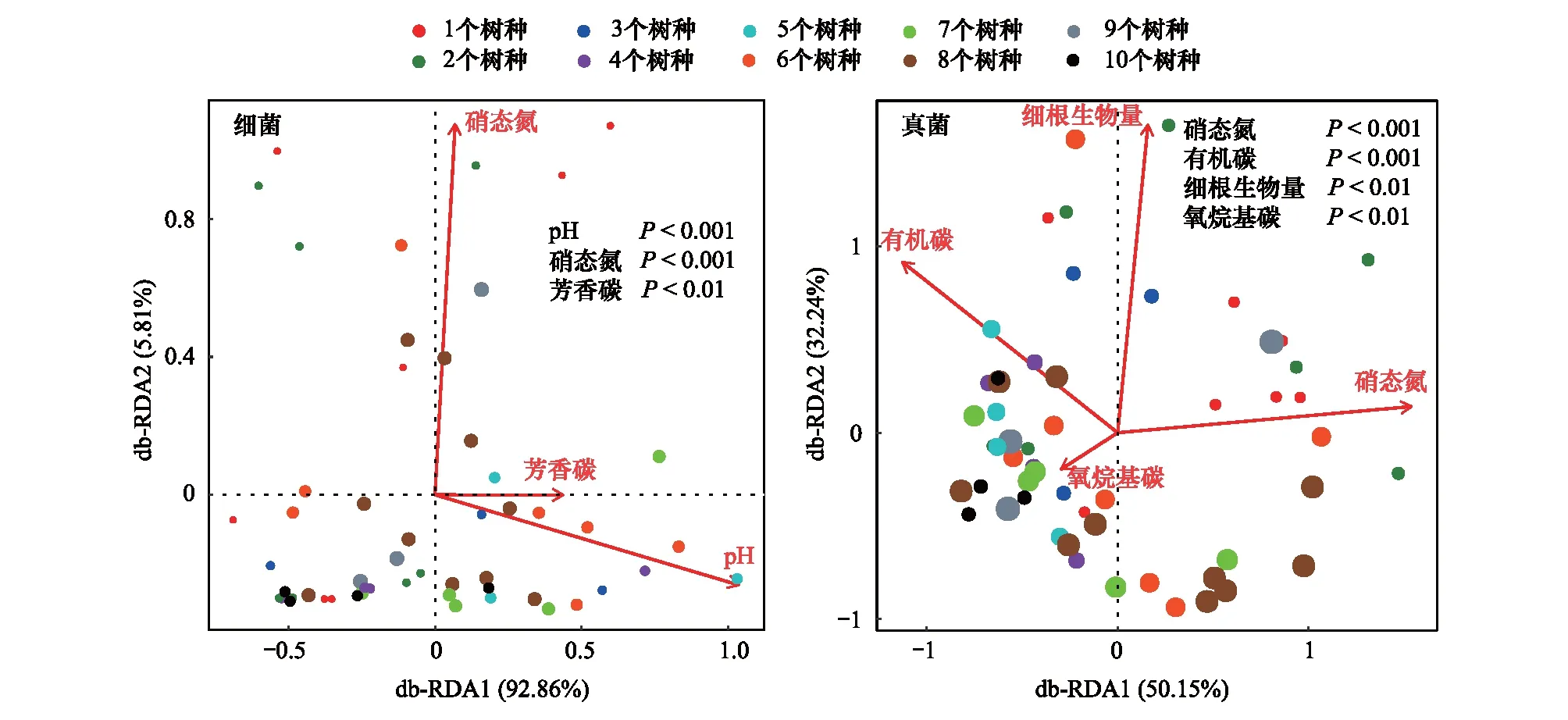
圖3 土壤細菌和真菌群落組成與環境變量的db-RDA分析Fig.3 Distance-based redundancy analyses (db-RDA) plot showing the relationship of abiotic factors to community composition of bacteria and fungi
不同樹種豐富度梯度下環境變量與土壤真菌群落組成的db-RDA分析如圖3所示。兩軸共解釋土壤真菌群落結構82.39%的變異,其中第一主軸(橫軸)累計解釋變量為50.15%,主要由土壤硝態氮貢獻;第二主軸(縱軸)累計解釋變量為32.24%,主要由細根生物量貢獻。結果表明土壤硝態氮、有機碳、細根生物量和氧烷基碳組分是影響土壤真菌群落組成在不同樹種豐富度梯度間差異的主要因素。
3 討論
3.1 南亞熱帶混交人工林樹種豐富度與土壤微生物α多樣性的關系
前人有關植物多樣性與土壤微生物群落組成的研究主要在草原[25]和天然林生態系統[26]開展,并發現植物多樣性增加會提高土壤真菌α多樣性。本研究中也得到了相似的結果,在南亞熱帶經近40年近自然管理,形成的多樹種、多層次、天然更新良好的異齡混交人工林生態系統中,土壤真菌α多樣性隨著樹種豐富度增加而提高(圖1)。通過人工混交不同鄉土樹種,進而構建成自然恢復并天然更新的混交人工林生態系統中樹種豐富度與土壤微生物α多樣性的關系趨近于天然草地和森林所呈現的規律的現象一般性的解釋是有更高物種豐富度的生態系統能更有效地利用各種資源,有更高的生產力并創造了更多的生態位從而容納了更多的土壤微生物(互補假說)[5,27]。但在本研究中還發現不同樹種豐富度下土壤細菌α多樣性差異不顯著,說明了樹種豐富度與土壤真菌α多樣性的相關性較之與土壤細菌α多樣性的相關性更明顯。Johnson[28]等研究認為細菌多樣性和植物多樣性不相關,這與本研究結果一致。有研究發現,土壤微生物更依賴于某種特定植物的存在(單一假說),而不是植物多樣性本身[29- 30]。因此,在下一步的研究中,將更深入的開展樹種功能特性對土壤微生物功能類群影響機制的研究,并進一步辨析地下微生物過程的變化是由樹種豐富度還是樹種特性主導。
本研究中相關分析結果表明,土壤細菌和真菌α多樣性與土壤理化因子呈現不同程度的相關性,說明該區域土壤理化因子是影響微生物數量的關鍵因素。Shen等[31]在長白山研究了6種典型植被類型,分析了隨海拔梯度土壤細菌群落組成和多樣性的變化趨勢,發現土壤細菌分布主要受土壤pH驅動。朱平等[32]通過研究祁連山4種不同植被類型對土壤微生物的影響也得出了相似的結論。在本研究中,試驗區內土壤細菌α多樣性與土壤pH和銨態氮顯著相關(P<0.05),印證了土壤pH同樣是影響本研究中南亞熱帶多樹種人工林生態系統樹種豐富度之間土壤細菌群落分布主要因子。本研究還發現樹種豐富度增加顯著提高了土壤pH,從而改善了研究區內土壤微生物生存環境,可能是土壤微生物多樣性增加的原因。莊林杰等[33]研究了從湖泊到岸邊深層土壤中土壤微生物變化情況,發現細菌多樣性與土壤銨態氮呈負相關,與本研究結果一致。其原因可能是土壤中銨態氮增加導致土壤酸化[34],而通常認為土壤細菌群落在中性pH條件下豐富度和多樣性較高[35],本研究中pH與土壤細菌多樣性的正相關關系也為此提供了依據。在森林生態系統中樹種豐富度增加會改變森林凋落物輸入量和凋落物分解過程,進而影響土壤中養分含量。有研究表明,真菌群落相較于細菌群落更直接的依賴于樹木凋落物,且許多真菌與樹種根系共生形成菌根,使得真菌類群與樹木的生物營養相互作用更強[36],這可能是本研究中真菌α多樣性受土壤有效磷影響的原因。
3.2 南亞熱帶混交人工林樹種豐富度變化影響土壤微生物群落組成
本研究的多樹種混交人工林生態系統中樹種豐富度增加顯著改變了土壤微生物群落組成,與以往在我國亞熱帶天然林開展的研究結果相一致[9]。土壤微生物分布廣泛且適應性強,但在特定區域的土壤微生物群落組成受到林下特定環境因素的調控。許多研究表明,在森林生態系統中植物群落多樣性對土壤微生物群落結構具有顯著影響[28,37]。土壤細菌群落以酸桿菌和α變形菌為主,但就具體細菌菌群而言,隨樹種豐富度增加菌群并沒有表現出明顯增加或者減少的趨勢。Xia[38]等人對我國典型森林土壤細菌群落的研究發現,土壤細菌豐度排在前五位的菌群分別為放線菌、酸桿菌、α變形菌、疣微菌和浮霉菌。本研究中不同樹種豐富度下土壤細菌群落中α變形菌差異性顯著。由此可見,樹種豐富度增加通過改變相對豐度較高的土壤細菌菌群使土壤細菌群落組成差異性顯著(圖2)。在本研究中土壤真菌群落組成變化也表現出相似的現象,傘菌綱作為土壤真菌群落中的重要組成菌群,在不同樹種豐富度梯度下差異性顯著(圖2)。
人工林不同樹種組成可通過影響森林生態系統中凋落物數量和質量,根系生物量、根系周轉和根系分泌物等對土壤理化性質和土壤養分產生不同的影響,進而改變土壤微生物的生物量和群落組成[3-4,36]。土壤pH參與土壤生物地球化學循環過程并對土壤理化性質具有重要的調節作用,是影響細菌群落組成的重要因子。本研究中db-RDA分析表明土壤細菌群落組成差異中的98.67%能夠被土壤pH、硝態氮和芳香碳組分三個環境變量所解釋,其中土壤pH占主要貢獻(圖3),結合土壤pH對土壤細菌α多樣性的顯著影響,說明了土壤pH是影響土壤細菌群落組成和多樣性的主要因素。適宜的pH為土壤微生物提供了良好的生存條件,土壤pH值能夠通過影響土壤基質的組成、化學性質從而改變土壤微生物多樣性和群落組成。本研究中土壤硝態氮、有機碳、細根生物量和氧烷基碳組分是導致土壤真菌群落差異的主要影響因素(圖3)。Wang[24]等人研究表明在南亞熱帶氮肥添加導致土壤中銨態氮和硝態氮含量增加,從而改變土壤pH,這可能是土壤硝態氮影響土壤真菌群落組成的原因。土壤有機質作為土壤理化性質的重要指標之一,是土壤養分循環轉化的核心。土壤有機碳含量的變化能夠影響土壤肥力和土壤微生物群落的變化[39]。王淼等[40]研究結果也表明土壤理化性質特別是土壤養分狀況的差異是土壤微生物群落結構差異的主要影響因素。樹種豐富度增加,往往會對根系的分布和生物量產生較大影響[41],通過改變根系分泌物從而導致土壤微生物群落變化。字洪標等[42]通過對青海7種不同林分土壤微生物群落結構的研究表明,細根生物量是導致土壤微生物群落差異的因素之一,與本文研究結果相一致。
4 結論
本研究選取了南亞熱帶多樹種混交人工林為研究對象,研究了人工林樹種豐富度對土壤微生物多樣性和群落組成的影響。結果表明,樹種豐富度增加對土壤微生物多樣性和群落結構具有一定的影響,其中土壤真菌表現出比細菌更加敏感的響應。土壤pH是影響土壤細菌多樣性和群落組成的主要影響因子,土壤碳氮組分和細根是影響土壤真菌多樣性和群落組成關鍵因素。研究結果說明了南亞熱帶不同鄉土樹種塊狀混交后天然更新形成多樹種混交林過程中,樹種組成和多樣性的變化通過改變土壤理化性狀和根系生物量對土壤微生物群落有顯著影響。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
