抗條銹病普通小麥-濱麥6BS·6NsS附加易位系的分子細胞遺傳學鑒定
2020-12-17 08:49:52杜少帥林益達于軍偉楊曉瑩吉萬全王長有
麥類作物學報 2020年8期
杜少帥,林益達,于軍偉,杜 欣, 楊曉瑩,吉萬全,王長有
(1.西北農林科技大學農學院,陜西楊凌 712100; 2.農業部作物基因資源與種質創制陜西科學觀測實驗站,陜西楊凌 712100)
小麥條銹病是由小麥條銹菌(Pucciniastriiformisf.sp.tritici(Pst))引起的最具毀滅性的小麥真菌病害之一,可導致嚴重的經濟損失。培育、種植抗條銹病品種是控制小麥條銹病最經濟、有效的策略,然而,因為病原菌毒性小種的變異,大多數品種(基因)會很快或最終喪失條銹病抗性[1,2]。因此,迫切需要培育新的抗條銹病材料或品種。
濱麥(Leymusmollis(Trin.) Pilger,2n=4x=28,NsNsXmXm) 是禾本科(Poaceae)小麥族(Triticeae)的一個異源四倍體材料,農藝性狀優良,且對小麥條銹病(stripe rust)、白粉病(powdery mildew)等多種真菌病害表現高抗或免疫,是改良普通小麥(TriticumaestivumL.)的重要種質資源。濱麥在小麥遠緣雜交中有著廣泛的應用,通過染色體工程產生了諸多類型的小麥-濱麥衍生材料(八倍體小濱麥[3]、部分雙二倍體[3]、附加系[4-7]、代換系[8-10]、易位系[11-12]和漸滲系)。攜帶濱麥抗條銹基因的小麥-濱麥衍生材料是小麥抗性育種重要的橋梁材料,其中,易位材料攜帶較少的不利基因,有較大的育種價值,關于小麥-濱麥易位材料的報道較少。易位材料通常由附加系和代換系衍生而來[13]。人工誘導產生易位如殺配子染色體[11,14-16]誘發易位的頻率較高,而染色體自發易位的頻率較低,但也有一定幾率發生[12,17]。
M13063A-1來自于小麥-濱麥二體異附加系M13063-3-3[18]經過2年嚴格自交的衍生后代。M13063-3-3是普通小麥7182和濱麥雜交后,用普通小麥山農20雜交一次,然后多年自交得到的小麥-濱麥衍生系,附加了一對濱麥第6部分同源群染色體,抗條銹菌生理小種條中31(CYR31)、條中32(CYR32)和條中33(CYR33)。本研究運用細胞學、原位雜交和分子標記等技術,結合形態學以及條銹病抗性調查,對自發易位系M13063A-1進行了鑒定,為該材料在小麥改良育種和抗病育種中的應用提供依據。
1 材料與方法
1.1 材 料
普通小麥7182、輝縣紅和小麥-濱麥衍生后代M13063A-1均于2018-2019年種植在西北農林科技大學試驗田,濱麥種植于室內光照培養箱中,PCR引物、華山新麥草(Psathyrostachyshuashanica,2n=2x=14,NsNs)基因組DNA、烏拉爾圖小麥(Triticumurartu,2n=2x=14,AA)基因組DNA、粗山羊草(Aegilopstauschii,2n=2x=14,DD)基因組DNA和以上所有材料都由西北農林科技大學農學院小麥遠緣雜交與染色體工程實驗室提供。條銹菌生理小種CYR31和CYR32,由西北農林科技大學植物保護學院 提供。
1.2 方法
1.2.1 細胞學鑒定
分別于3月下旬和4月中旬在田間取材料的根尖和幼穗。根尖、幼穗的固定和鏡檢按照Yang等[3]的方法進行。用Olympus BX-43 (Japan)顯微鏡觀察并拍照。
1.2.2 原位雜交
種子在23 ℃萌發2~3 d,直至幼根長至2~3 cm,將根尖分生區切下,根尖的固定按照Zhang等[19]的方法進行。染色體制片采用滴片法[20]。基因組原位雜交(genomicinsituhybridization,GISH)按照Yang等[3]的方法進行。熒光原位雜交(fluorescenceinsituhybridization,FISH)按照Han等[21]的方法進行,結果參考Tang等[22]發表的普通小麥中國春和綿陽11的FISH核型。用Olympus BX53(Japan)熒光顯微鏡觀察并拍攝圖像,用Photoshop CC(Adobe,USA)分析并處理圖像。
原位雜交探針:按照Yang等[4]的方法,用CTAB法提取幼嫩葉片的DNA。用濱麥基因組DNA探針(綠)進行減數分裂中期ⅠGISH;用寡核苷酸探針Oligo-pSc119.2(綠)、Oligo-pTa535(紅)和濱麥基因組DNA探針(綠)進行連續的FISH和GISH(FISH-GISH);用華山新麥草基因組DNA探針(綠)進行GISH;用烏拉爾圖小麥基因組DNA探針(綠),濱麥基因組DNA探針(綠)和粗山羊草基因組DNA探針(紅)進行多色GISH(multi-color GISH,mc-GISH)。以濱麥基因組DNA探針(綠)、Oligo-pSc119.2(綠)分別和三種寡核苷酸探針Oligo-pTa71-2[22](紅,著絲粒探針)、Oligo-119[23](紅)、Oligo-60[23](紅)結合進行GISH,來探究2B、6B和7B染色體的FISH信號模式以及易位染色體中小麥染色體片段的來源,試驗結果分別參考Oligo-pTa71-2、Oligo-119、Oligo-60在B基因組染色體上產生的信號模式[22-23]進行分析。
1.2.3 分子標記分析
分布在小麥第6同源群染色體上的18個PLUG標記、176個EST標記和106個SSR標記被用來確定M13063A-1所攜帶的濱麥染色體片段的同源群歸屬。PCR和電泳分析按照Zhu等[24]的方法進行。
1.2.4 農藝性狀和條銹病抗性鑒定
在2019年6月收獲植株后,調查并記錄M13063A-1和普通小麥7182的株高、分蘗、穗長、小穗數、小穗粒數。M13063A-1與普通小麥7182之間的差異顯著性用T檢驗進行分析。
在試驗植株成株期,于田間環境條件下接種條銹菌生理小種條中31和條中32的混合菌種進行M13063A-1的成株期條銹病抗性鑒定,以感病對照輝縣紅作誘發行,將孢子均勻地撒在噴濕的植株上進行人工誘發。當輝縣紅充分發病時,按照Bariana等[25]的“0~4”級分級標準對M13063A-1、7182和輝縣紅進行抗病等級鑒定。
2 結果與分析
2.1 細胞學鑒定結果
根尖分生區細胞染色體數目觀察結果(圖1a)顯示,M13063A-1體細胞中含有44條染色體。花粉母細胞鏡檢結果(圖1b)顯示,其減數分裂中期Ⅰ染色體配對構型為2n=44=22Ⅱ,其中一對染色體提前分離;在減數分裂后期Ⅰ(圖1 c)同源染色體可以均等分離。此外,在減數分裂中期Ⅰ細胞中沒有觀察到多價體形成。這些結果說明M13063A-1在細胞學上穩定遺傳。

a:有絲分裂中期;b:減數分裂中期Ⅰ;c:減數分裂后期Ⅰ。箭頭指示提前分離染色體。
2.2 原位雜交結果
用濱麥基因組DNA探針對M13063A-1根尖細胞染色體組進行GISH,結果(圖2b)顯示,M13063A-1含有兩條易位染色體。用濱麥基因組DNA探針對M13063A-1幼穗細胞染色體組進行GISH,結果(圖2a1~圖2a4)顯示,兩條易位染色體在減數分裂中期Ⅰ配對成二價體(圖2a1~圖2a3),在減數分裂后期Ⅰ均等分離到細胞兩極(圖2a4),這表明M13063A-1可以穩定遺傳。在減數分裂中期Ⅰ,有一對普通小麥染色體提前分離(圖2a3),在實際觀察中,也有一對易位染色體提前分離的情況出現(未提供照片)。在減數分裂中期Ⅰ細胞的GISH結果中,并未觀察到多價體的形成。用華山新麥草基因組DNA探針對M13063A-1根尖細胞染色體組進行GISH(圖2c),也觀察到兩條易位染色體,這表明M13063A-1攜帶的濱麥染色體片段來自Ns基因組染色體。
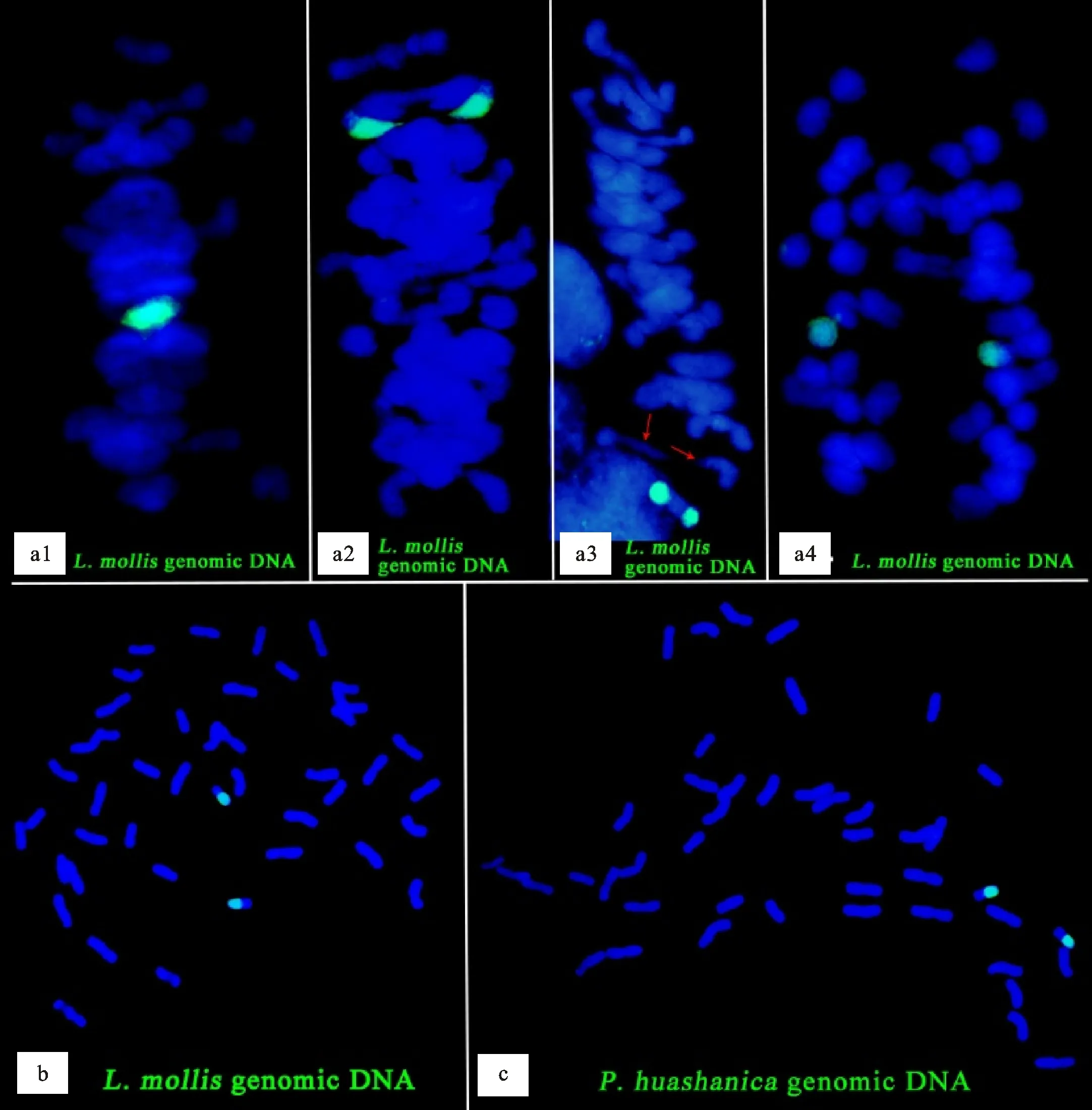
a1、a2、a3:用濱麥基因組DNA探針進行減數分裂中期Ⅰ原位雜交;a4:用濱麥基因組DNA探針進行減數分裂后期Ⅰ原位雜交;b:用濱麥基因組DNA探針進行有絲分裂中期原位雜交;c:用華山新麥草基因組DNA探針進行有絲分裂中期原位雜交。紅色箭頭指示提前分離的一對普通小麥染色體。
FISH-GISH結果(圖3a、圖3b)顯示,M13063A-1含有42條普通小麥染色體和一對易位染色體。易位染色體長臂是濱麥染色體片段,短臂是普通小麥染色體片段。寡核苷酸探針Oligo-pTa535在易位染色體長臂末端即濱麥染色體端部產生了微弱的紅色信號。42條普通小麥染色體中,2B、6B、7B染色體暫時不能分辨清楚(如圖3a中白色六角星所示)。圖3c是根據圖3a和圖3b整理的M13063A-1的核型,寡核苷酸探針Oligo-pSc119.2(綠)和Oligo-pTa535(紅)可以區分M13063A-1中的42條普通小麥染色體,并且B基因組染色體上的綠色信號各不相同,這說明只用Oligo-pSc119.2就可以區分B基因組染色體。
用烏拉爾圖小麥基因組DNA探針(綠)、濱麥基因組DNA探針(綠)和粗山羊草基因組DNA探針(紅)進行mc-GISH,結果(圖4a)顯示,A基因組染色體(顯示白色)、B基因組染色體(顯示藍色,即無信號)和D基因組染色體(顯示紅色)分別有14條,且濱麥基因組DNA探針(綠)在易位染色體長臂上產生了明亮的綠色信號,在短臂上無信號(顯藍色),說明易位染色體的短臂可能來源于B基因組染色體。因為只用寡核苷酸探針Oligo-pSc119.2就可以區分B基因組染色體,所以用濱麥基因組DNA探針(綠)、Oligo-pSc119.2(綠)分別和三種在B基因組染色體上產生特異信號的寡核苷酸探針Oligo-pTa71-2(紅)、Oligo-119(紅)、Oligo-60(紅)結合進行GISH,確定了M13063A-1中2B、6B和7B染色體的FISH信號模式(圖4b1~圖4b3、圖4c)。如圖4b1和圖4c所示,隨體探針Oligo-pTa71-2在1BS、6BS和易位染色體短臂上都產生了明顯的紅色信號,但1BS上的紅色信號明顯比6BS和易位染色體強烈,說明易位染色體的短臂是6BS,而且紅色信號產生在易位染色體短臂的末端,說明易位染色體不含有6B的隨體。
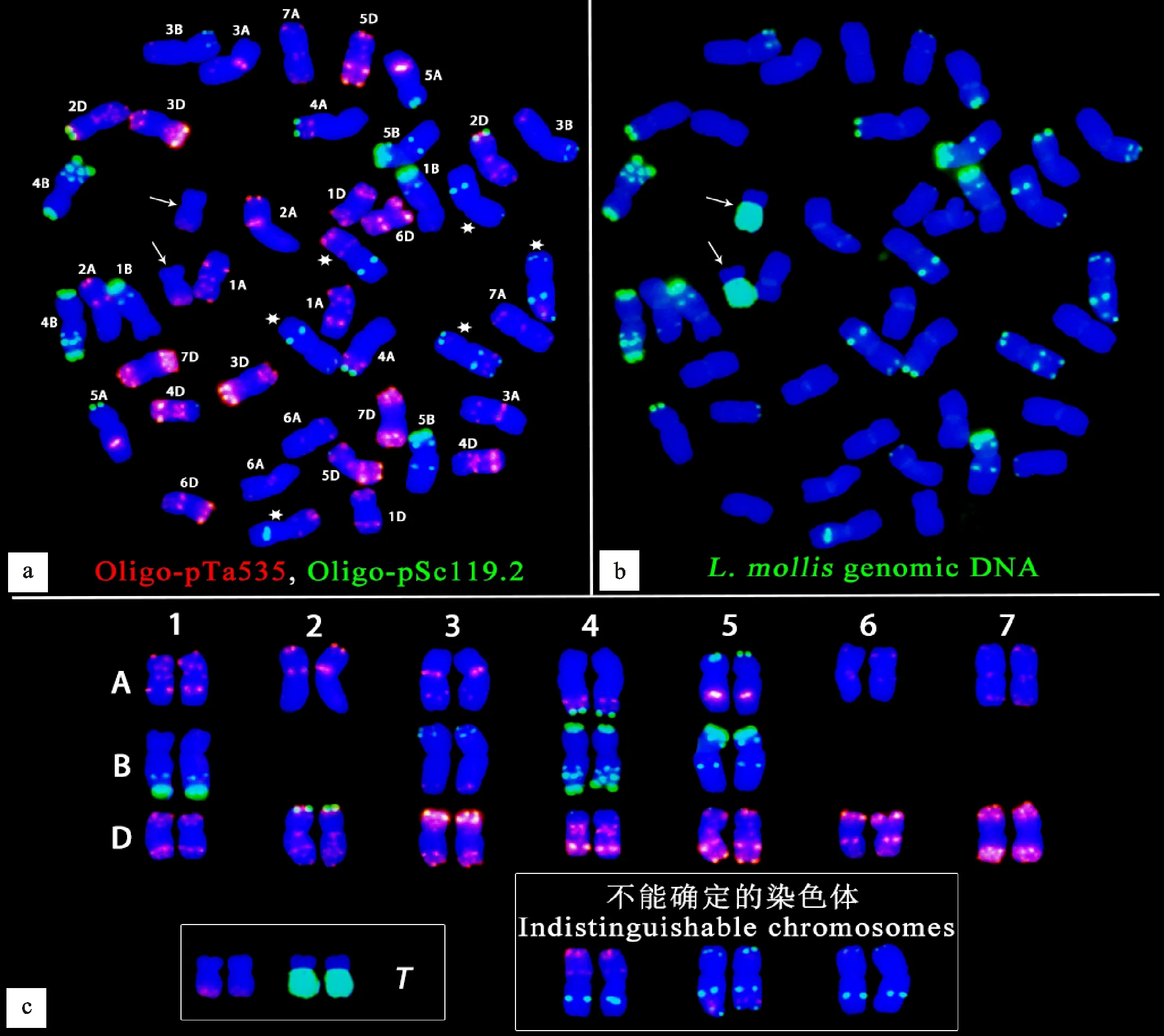
a:FISH;b:GISH;c:核型。白色箭頭指示易位染色體,白色六角星指示暫時不能分辨的3對普通小麥染色體。T表示易位染色體。
2.3 分子標記分析結果
在定位于第六同源群的PLUG、EST和SSR標記中,最終篩選出1個PLUG特異標記(TNAC1677)、3個EST特異標記(BQ159615、CD454198、CD454353)和2個SSR特異標記(Xwmc672、Xcwm532)(表1),以上6個標記均在濱麥和M13063A-1中擴增出了明顯的特異性條帶(圖5)。TNAC1677、BQ159615、CD454198和CD454353位于小麥第六同源群染色體短臂,結合以華山新麥草基因組DNA為探針的GISH結果(圖2c),說明M13063A-1攜帶的濱麥染色體片段為6NsS。
2.4 農藝性狀和條銹病抗性
農藝性狀鑒定結果(表2和圖6a、圖6b、圖6c)表明,M13063A-1的株高極顯著低于親本7182,穗長極顯著小于親本7182。在成株期,M13063A-1、7182和輝縣紅的條銹病抗性等級分別為1級、3級和4級(圖6d),這表明M13063A-1在成株期抗條銹菌生理小種條中31和條中32。
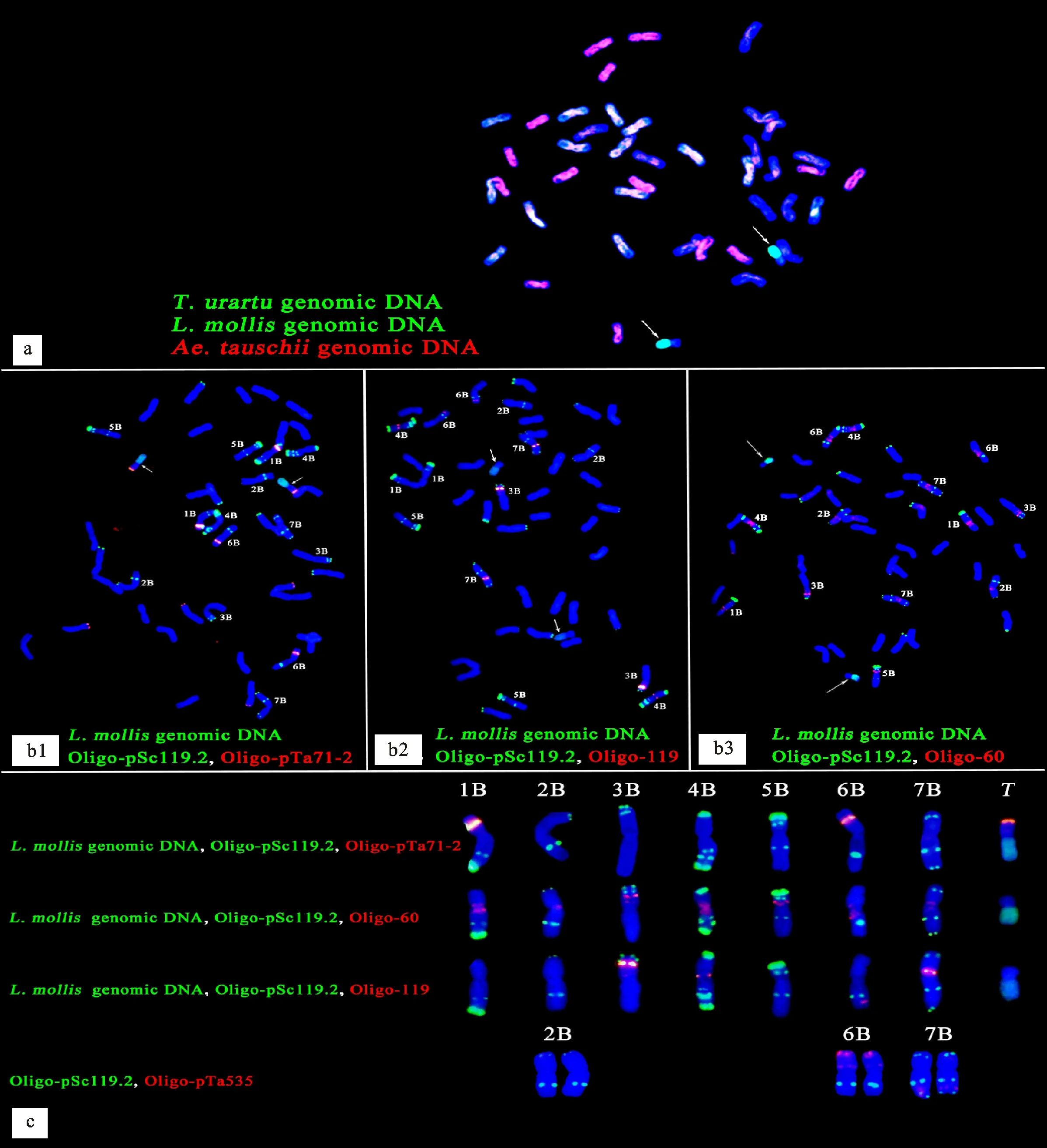
a:以烏拉爾圖小麥基因組DNA(綠)、濱麥基因組DNA(綠)和粗山羊草基因組DNA(紅)為探針的多色GISH;b:分別以Oligo-pTa71-2(紅)、Oligo-119(紅)、Oligo-60(紅)和濱麥基因組DNA(綠)、Oligo-pSc119.2(綠)為探針的GISH(b1:Oligo-pTa71-2;b2:Oligo-119;b3:Oligo-60);c:根據圖4 b1、b2、b3整理的B亞基因組染色體和易位染色體信號模式。白色箭頭指示易位染色體。T表示易位染色體。

表1 M13063A-1攜帶的濱麥6NsS連鎖的PLUG、EST和SSR標記Table 1 PLUG,EST and SSR polymorphic markers mapped on Lm#6NsS in M13063A-1
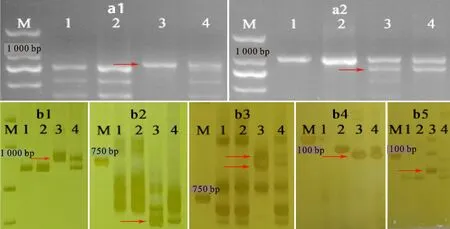
M:D2000;1:7182;2:山農20;3:L.mollis;4:M13063A-1;a1:TNAC1677-HaeⅢ;a2:TNAC1677-TaqⅠ;b1:BQ159615;b2:CD454198;b3:CD454353;b4:Xwmc672;b5:Xcwm532。箭頭指示特異條帶。

表2 M13063A-1及其親本7182的主要農藝性狀比較Table 2 Agronomic traits of M13063A-1 and its parent 7182

a:植株;b:穗;c:小穗和籽粒;d:條銹病抗性;1:7182;2:濱麥;3:M13063A-1;4:輝縣紅。
3 討 論
3.1 附加易位系M13063A-1形成的原因
在二體異附加系M13063-3-3進行減數分裂時,6B染色體和6Ns外源染色體發生同源重組是形成6BS·6NsS易位染色體的前提。由于部分同源性,一條6B染色體與一條6Ns外源染色體配對且發生易位,形成6BS·6NsS易位染色體和6BL·6NsL易位染色體,而另一條6B染色體和6Ns染色體不配對或者進行配對但是不發生易位。在形成配子時,產生含有21條普通小麥染色體和一條6BS·6NsS易位染色體的雄配子和雌配子,雄配子和雌配子結合,形成含有42條普通小麥染色體和一對6BS·6NsS易位染色體的合子,進而形成了附加易位系M13063A-1。
3.2 M13063A-1中的染色體結構變異
Han等[13]在小麥-中間偃麥草衍生系Z4、Z5和Z6中發現復雜的易位和重組,這些結構變異都是自發產生的,其中,Z5體細胞染色體數目為44條,在普通小麥基因組上附加了一對易位染色體,該易位包含A組、D組和中間偃麥草的染色體片段。M13063A-1和Z5的不同是其易位只涉及小麥B基因組染色體。這種附加易位系出現的原因尚不確定,有待進一步探究。
在M13063A-1減數分裂中期Ⅰ細胞的GISH結果中,并未觀察到6BS·6NsS易位染色體和6B染色體形成的多價體,這與預期結果不符。(GAA)10[26](圖片未提供)和寡核苷酸探針Oligo-pTa535、Oligo-119、Oligo-60都未在易位染色體片段上產生信號,結合原位雜交結果,推測易位染色體片段是6BS上包含次縊痕的一小段片段,易位染色體不含有6B染色體的隨體和著絲粒,其著絲粒是濱麥6Ns著絲粒。端粒花束的形成和著絲粒配對在同源染色體的配對過程中起著至關重要的作用[27],但是隨體是否在1B和6B染色體的同源配對過程中起作用尚不清楚。易位染色體不含有6B的著絲粒和隨體,這可能是M13063A-1在減數分裂中期Ⅰ沒有多價體形成的原因。
3.3 M13063A-1抗條銹病基因的來源
攜帶外源染色體的衍生材料是利用外源優異基因的重要種質資源。Bao等[1]和Li等[2]都曾在小麥-濱麥衍生系中定位到了來自濱麥的條銹病抗性基因。在本研究中,雖然M13063A-1抗條銹病,且其親本之一7182不抗病,但因缺少另一親本山農20的條銹病抗性表現,所以無法確定M13063A-1的抗條銹病基因來源。因此,M13063A-1的條銹病抗性是否來源于濱麥有待進一步鑒定。
