土壤病毒的研究進展與挑戰*
2021-01-05 13:25:30王光華劉俊杰朱永官
土壤學報 2020年6期
關鍵詞:研究
王光華,劉俊杰,朱 冬,葉 茂,朱永官,4
(1. 中國科學院東北地理與農業生態研究所/中國科學院黑土區農業生態重點實驗室,哈爾濱 150081;2. 中國科學院生態環境研究中心,北京 100085;3. 中國科學院南京土壤研究所/中國科學院土壤環境與污染修復重點實驗室,南京 210008;4. 中國科學院城市環境研究所,廈門 361021)
病毒是個體非常微小、結構簡單,由蛋白質外殼包被的內部含有遺傳物質核酸的非細胞型生物實體。按照核酸的組成,病毒可劃分為雙鏈DNA(dsDNA)、單鏈DNA(ssDNA)、雙鏈RNA(dsRNA)、正單鏈RNA(ssRNA+)和負單鏈RNA(ssRNA-)。通常病毒被認為是不具有酶系統,不能夠獨立繁殖生活,病毒的繁殖必須依賴于寄主系統,所以長期以來病毒是否被視為生物一直存在爭議[1]。但這種現象目前有所改變,科學家們從多種環境中發現了一些感染原生動物的病毒粒子,其尺寸較大,被稱為巨大病毒,如梅格病毒(Megavirus)和潘多拉病毒(Pandoravirus)粒子直徑分別達到440 nm和1 000 nm,其基因組大小分別為1.26 Mbp和2.5 Mbp,這些巨大病毒具有獨特的酶系統[2-4],有的病毒還被其他病毒(噬病毒體Virophage)侵染[2,5],這些發現動搖了學術界對生命之樹的認知,有觀點認為病毒可看做為生命的第四個域——病毒域[6]。事實上,病毒可以侵染生命之樹上其他三域的生物,通常學術界將侵染細菌和古菌的病毒,即侵染原核微生物的病毒稱為噬菌體。目前國際上分離獲得的細菌病毒有6 000多株,而古菌病毒僅100多株[7]。與細菌病毒遺傳物質組成不同,古菌病毒的遺傳物質多是dsDNA,少數是ssDNA,尚未發現含有RNA的古菌病毒[8]。有觀點認為病毒可能較地球上所有生物的共同祖先露卡(LUCA)出現的更早[9],地球上所有具有細胞結構的生物都能夠被一種甚至多種病毒所侵染[10],病毒在侵染過程中基因不斷地突變,也不斷地與寄主間進行遺傳物質的交流,從而確保病毒能夠生存下去,也使寄主生物獲得新的基因,產生新的性狀,促進了地球生物的進化演替。
土壤是各種生物的重要棲息地,土壤中存在著數量巨大,種類繁多的病毒生命體。雖然學術界普遍意識到病毒在土壤各種生態過程中可能起到重要的作用,但由于受到土壤的異質性、多樣性,以及研究手段等方面的限制,目前國際上對土壤病毒研究的重視程度遠落后于流動的海洋環境[11-13]。本文從生態學角度,基于目前國內外病毒研究現狀,尤其是針對土壤病毒的研究給予回顧,期望起到拋磚引玉的作用,引起國內同行對土壤病毒研究的關注和重視。
1 土壤病毒形態與數量
1.1 土壤病毒的形態
病毒大小通常是納米級的,普通光學顯微鏡無法捕捉到病毒的身影,必須通過透射電子顯微鏡(TEM)才能觀測到。病毒形態多種多樣,目前發現古菌病毒形態有16種,而細菌病毒形態有9種,可見雖然目前得到的古菌病毒純分離株遠少于細菌病毒,但其形態多樣性卻高于細菌病毒[7]。土壤中的古菌病毒和細菌病毒多以具有頭尾結構的有尾噬菌體為主。有尾噬菌體是dsDNA病毒,按其尾部形態特征可劃分為短尾(Podophage)、長尾(Siphophage)和肌尾(Myophage)噬菌體[7]。土壤中其他病毒形態還有絲狀、桿狀、棒狀、瓶狀、水滴狀、球狀和二十面體狀等[8]。真核生物病毒沒有頭尾結構,形態更加多樣,有的蛋白質外殼上還有修飾成分,與原核生物病毒有很大的不同(圖1)。

圖1 侵染細菌、古菌和真核生物的dsDNA病毒分類及形態示意圖(改自Prangishvili等[14])Fig. 1 Classification and morphological diagrams of the dsDNA viruses that infect bacteria,archaea and eukaryotes(Modified from Parangishvili et al. [14])
1.2 土壤病毒存在形式
病毒在土壤中的存在形式大致可歸為3種:少部分病毒以游離狀態存在于土壤溶液中;大部分病毒被吸附在土壤顆粒上;還有一部分是溫和性噬菌體(lysogenic or temperate phage),其基因組插入在寄主基因組上,以溶原(prophage)狀態存在于細胞內[11,15]。溫和性噬菌體在土壤中普遍存在,尤其是在寄主分布不均勻[16]、寄主數量少[17-18]或寄主細胞生長狀況不好[17,19]的情況下,噬菌體以溶原狀態伴隨著寄主細胞繁殖而繁殖,避免受到外面不利環境條件的影響,從而確保噬菌體在寄主種群中能夠長期存活繁殖下去的最佳方式。一般而言,在某個環境中病毒與細菌比值(VBR)數值小時,噬菌體主要以溶原狀態存在,而當VBR數值大時,則以烈性噬菌體(lytic phages)為主[20-22]。相比于營養貧乏的海水環境,營養豐富的土壤中溶原性噬菌體占比較高[20,23]。溶原性噬菌體也是極端干熱沙漠條件下噬菌體主要存在形式[24],而在冷沙漠中噬菌體主要以游離的烈性噬菌體為主[21]。溶原狀態噬菌體在土壤中存在,也可通過土壤細菌全基因組測序而發現,有研究表明許多細菌基因組中含有1個或多個前噬菌體元件(prophage element)[25],其中默冗子(moron)是前噬菌體元件的一個重要部分[26-27]。默冗子在噬菌體基因組中不具有編碼功能,但當它整合在細菌基因組中則通過基因溶原性轉換(lysogenic conversion),使細菌獲得新的性狀和功能,提高細菌環境生存適應能力[28-30]。
1.3 土壤病毒數量
目前地球生物圈上到底有多少病毒還沒有準確數據,普遍認為全球病毒數量>1031[31]。有學者依據不同生境中病毒與細菌或與微生物的比值(VBR或VMR),推算出全球病毒粒子的數量為4.80×1031,其中在沉積物和土壤中數量分別占到87%和10%,而在海洋水體中只占2.7%,可見土壤環境中病毒數量非常巨大[32]。不同環境中病毒豐度存在較大差異,將病毒顆粒(virus-like particles,VLPs)用熒光核酸染料染色,然后在熒光顯微鏡(EFM)下計數發現,每毫升海水中VLPs在105~107個之間[33-34],河口或湖泊淡水中VLPs可達到每毫升108個[35];而在森林土壤和農田土壤中,每克土壤VLPs數量分別在1.31×109~4.17×109個和0.87×109~1.1×109個之間[36]。最近,Williamson等[37]對熱帶沙漠、寒帶沙漠、森林、農田和濕地等環境下的土壤病毒和細菌數量進行Meta數據分析發現,病毒在沙漠土壤中豐度最低,森林土壤中豐度最高,土壤類型與病毒數量高度相關(r=0.803,P<0.001);發現土壤病毒數量與土壤細菌數量呈高度正相關(r=0.647,P<0.001);發現土壤病毒數量與土壤pH呈顯著負相關(r= -0.352,P=0.009)。同時他們也認為土壤溫度和濕度也是影響土壤病毒數量的重要土壤因子,但由于數據的缺乏,兩者之間的相關性尚未建立起來[37]。需要指出的是,目前對土壤病毒數量的研究工作主要是針對土壤中游離的和被土壤顆粒吸附的病毒,尚無法大規模地開展針對土壤中溶原性病毒的計數研究,所以現有觀察到的數據可能遠低于土壤中病毒的真實數量。
由于大部分病毒被吸附在土壤顆粒上,如何從土壤中將病毒高效率地分離提取出來是準確測定病毒數量的前提條件。美國學者Wommack團隊[38]對比研究了10%的牛肉膏溶液、250 mmol·L-1甘氨酸溶液、10 mmol·L-1焦磷酸鈉溶液和1%檸檬酸鉀溶液從粉砂壤土和砂壤土中提取病毒的效果,發現1%檸檬酸鉀溶液是最佳的土壤病毒浸提劑。在此基礎上,Trubl等[39]改進了提取劑配方,發現1%檸檬酸鉀溶液+10%磷酸鹽緩沖液+5 mmol·L-1的EDTA溶液+150 mmol·L-1硫酸鎂溶液對富含有機質的泥炭和沼澤土壤中病毒提取效率最高,提取出的病毒數量是其他提取劑的2倍。而張輝等[40]以MS2和φx174作為指示病毒,研究發現含有0.04 mol·L-1焦磷酸鈉的3%牛肉膏溶液提取效果最好,用此提取劑指示病毒在紅壤土、紅黏土和潮土上的回收率達63%~98%,但在黃泥土上的回收率僅30%。可見同一種病毒提取劑的提取效率因土壤類型而異。需要注意的是,從土壤中提取病毒溶液的操作過程多伴隨采用超聲波、渦旋、劇烈震蕩,有的甚至采用珠打法(bead-beating)等物理分散技術,這些劇烈的分散過程可以導致有尾噬菌體尾部結構折斷或丟失,從而降低了觀察到的有尾噬菌體比例[41-42]。
VMR或VBR數值大小也能間接地反映出環境病毒數量的高低。VBR在不同環境中的變幅很大,如在海洋和湖泊水體中數值在3~25之間[33-34,43],而在稻田水中變幅在0.11~72之間[44]。與水體環境相比,土壤中VBR變幅更大,有研究發現森林和農田土壤中的VBR分別為10和3 000[36],而在沙漠土壤中其數值僅0.15~1.66[45]。在根際微域環境中,有研究發現每克小麥根際土壤中VLPs是1.18×109個,與非根際土壤病毒數量沒有顯著差異,從而導致根際VBR為0.27,而非根際土壤是4.68[41]。這一發現表明盡管根際土壤細菌數量非常大(根際效應),但根際細菌病毒并沒有體現出同步的增加規律,即根際細菌病毒數量沒有體現出根際效應。這種現象與通常認為的細菌-病毒生長模型“殺死優勝者Kill-the-Winner”不符合[34,46]。最近,來自國外科學家在對珊瑚礁后生動物黏膜表面細菌和病毒之間相互作用研究中發現一個新的細菌-病毒生長模型,即“搭乘優勝者Piggyback-the-Winner”[47]。在這個模型中,病毒以溶原狀態將病毒基因整合到寄主細胞體內,隨著寄主細胞繁殖而繁殖,避免與其他病毒競爭,也避免觸動宿主自身的免疫系統。這種“搭乘優勝者”模型是否是植物根際細菌病毒的生存策略還未見直接證據支持。此外,也有學者認為根際細菌病毒數量與非根際土壤無差異的原因也可能與根際細菌受到根系分泌物選擇的作用而不易于被病毒侵染有關[13]。
1.4 土壤病毒生物量
全球病毒數量保守估計在1031以上,那么這些病毒生物量是多少呢?Cobián Güemes等[32]根據一個典型噬菌體含有50 kbp的DNA質量為0.054 fg(飛克),而殼蛋白質質量為0.028 3 fg,推算出全球4.80×1031VLPs的質量是3.95×1015g(3.95 Pg或3.95 Gt)。按照典型病毒化學計量學C︰N︰P =20︰6︰1計算[48],全球病毒生物量估算有2.92 Pg C、0.88 Pg N和0.15 Pg P。如果全球原核生物含有350~500 Pg C[49],那么病毒生物量碳約占原核生物量碳的0.58%~0.83%。而另一篇文獻認為全球所有生物只有大約550 Gt C,其中細菌為70 Gt C,古菌大約為7 Gt C,病毒含有大約0.2 Gt C[50],這一估算數值較Cobián Güemes等[32]報道數據縮小了10倍,但兩篇文獻均表明病毒生物量相對于原核生物生物量而言非常小。與細菌細胞C︰N︰P的化學計量為60︰16︰1相比[51],細菌病毒顆粒富含N和P,尤其是P元素。雖然病毒生物量相比原核生物和真核生物而言非常小,但其數量巨大,周轉快,在全球生物地球化學循環中起到非常重要的作用[52]。以海洋環境為例,有研究估測每天有1028個細菌被病毒感染而裂解,其中大約30%是具有光合作用的藍細菌,60%是異養細菌[52-53]。海洋中某些區域有超過5%的可溶性有機磷和氮來源于病毒顆粒[48]。有報道指出海洋環境中大約6%~25%的碳循環量是由病毒驅動的[33],但在土壤環境中,病毒是如何及在多大程度上驅動碳循環過程還鮮見報道。
2 土壤病毒多樣性
由于病毒不能獨立繁殖,而大量環境微生物的不可培養性,無疑限制了對病毒多樣性的認知。全球到底有多少種病毒尚無確切的答案。現有的報道均為基于推測的結果,例如有文獻報道[54-55],全球有1 000萬種微生物(主要是細菌和古菌),而每種微生物可被10種不同的病毒侵染,則全球會有超過1億種的病毒[56-57]。筆者認為這種推測有可能高估了病毒的多樣性,因為隨著研究的不斷深入,發現病毒與被侵染寄主之間具有很強專一性的觀點被不斷修正[58],如有些細菌病毒在細菌分類屬、目,甚至在門水平上能夠跨物種感染不同寄主[59-60]。最近,Williamson 等[37]基于病毒宏基因組數據分析發現土壤中病毒基因型(genotype)在1 000~1 000 000之間;與土壤相對應,海洋中病毒基因型在532~129 000之間,在淡水環境為400~40 000之間[61]。可見土壤病毒多樣性的上限和下限均高于水體環境中的病毒,充分說明土壤病毒是一個巨大的未知基因資源庫。
2.1 分子標記基因解析病毒多樣性
采用保守性的引物,利用PCR技術對原核微生物的16S或真核生物18S rRNA基因片段進行擴增,然后對PCR產物進行基因信息解析,是目前研究環境微生物基因多樣性和群落組成的主流技術。然而對于病毒而言,由于其基因組的高度變異性和雜合性,在病毒基因組中尚未發現保守序列設計出通用引物,用于環境病毒基因多樣性解析[62]。但有研究發現,針對一些特定病毒家族,其基因組中編碼某些結構或功能蛋白的氨基酸片段序列高度保守,可基于此序列設計出簡并性引物,然后從環境中提取eDNA,直接進行PCR擴增目標基因,測序分析特定病毒家族遺傳基因多樣性及分布格局[63]。2014年Adriaenssens和Cowan[64]在一篇綜述文章中歸納出多個病毒標記基因可用于解析環境特定病毒多樣性及其生態學研究。需要說明的是這些標記基因多是針對海洋、湖泊等水體環境中細菌病毒基因多樣性研究的,適用于土壤細菌病毒研究最常用的標記基因是g23,該基因是編碼T4型噬菌體主要殼蛋白的結構基因。該基因最先是由Filée等[65]發現可用于研究海洋中T4型噬菌體多樣性,隨后Jia等[66]證明g23基因也可適用于研究稻田土壤中T4型噬菌體基因多樣性,從而開啟了g23基因在土壤病毒研究中應用的序幕[67-68]。針對該基因,中國科學院東北地理與農業生態研究所農田分子生態學科組從我國東北稻田、濕地和黑土農田中捕捉到數百條不同的T4型噬菌體g23基因,發現旱地黑土農田T4型噬菌體分布與其在海洋和湖泊水體環境中明顯不同,而與稻田環境相近,從黑土農田獲得的基因序列中建立了多個新的T4型噬菌體g23基因群[69-70]。發現即使在相同稻田環境,中國東北稻田T4型g23基因群集(Assemblage)與日本稻田不同,由此得出T4型噬菌體分布受到地理分隔和生態過程的雙重調控的觀點[70-71]。此外,我們還對g20基因(編碼藍藻肌尾噬菌體殼組裝蛋白)[72-73]、DNApol基因(編碼藍藻短尾噬菌體DNA聚合酶基因)[74-75]、輔助代謝基因psbA(編碼藍藻噬菌體光合蛋白基因)[76]和phoH(編碼噬菌體磷酸鹽調節基因)[77-78]在東北稻田和濕地中基因組成及多樣性進行了研究。揭示出這些標記基因表征的噬菌體群落結構組成在稻田和濕地環境中與海洋環境完全不同,并且在濕地與稻田間也存在差異,建立了多個獨特的稻田和濕地噬菌體類群。由此可見,一些用于研究海洋環境噬菌體多樣性的標記基因也適合用于陸地生態環境,說明細菌病毒有著共同的起源祖先;而基于分子標記基因群集表征的特定噬菌體群落結構在海洋、濕地、稻田,乃至旱地土壤中的不同,也從一個側面顯示出細菌病毒從海洋到陸地生態系統的演替過程。
2.2 RAPD-PCR解析病毒多樣性
由于尚未發現通用引物適應于病毒多樣性研究,如何解析環境特別是土壤中病毒多樣性是一個挑戰性課題。隨機擴增多態性DNA標記(Random Amplified Polymorphic DNA ,RAPD)是采用隨機的10個堿基核苷酸序列同時作為正向和反向引物進行PCR擴增,是對未知序列基因組進行多態性分析的技術[79]。Winget和Wommack[80]首次將該技術運用到美國切薩皮克灣水體病毒群落研究,他們根據擴增產物凝膠電泳圖譜中條帶的位置(長度)、亮度(豐度)分布情況,分析出不同樣本間病毒群落結構及其相似度,然后對條帶進行切膠測序,分析病毒組成。這種分析方法與DGGE-PCR技術解析環境微生物群落結構流程基本類似。利用該技術,Helton和Wommack[81]進一步分析了切薩皮克灣沉積物中病毒基因多樣性,發現沉積物病毒結果存在明顯的時空分變化;Srinivasiah等[82]揭示出南極不同地點土壤病毒群落結構存在明顯差異。中國科學院沈陽應用生態研究所徐慧團隊[83]采用該技術并結合高通量測序技術,對長期灌溉和施用尿素的稻田土壤中病毒和細菌群落豐度及多樣性進行動態解析,發現添加尿素延遲了病毒豐度峰值的出現,說明病毒對氮肥添加非常敏感。需要注意的是,運用該技術的前提條件是要確保提取到的DNA沒有寄主細胞遺傳物質的干擾,然后才可進行PCR擴增。此外,也要考慮到單個隨機引物擴增的偏好性,會導致在對病毒群落結構解析時存在偏差。故此,應用該技術時,建議采用多個隨機引物進行擴增,然后將不同引物擴增得到的凝膠電泳條帶圖譜整合為一個數據集,再進行病毒群落結構解析。
2.3 宏基因組學解析病毒多樣性
雖然RAPD-PCR技術在一定程度上能夠對不同樣本病毒群落結構進行解析,但該技術尚不能對病毒成員進行定性描述,還需后期大量的條帶切膠測序工作。隨著基因測序技術的飛速發展,高通量測序技術已成為研究環境微生物組的主流技術。其中宏基因解析技術目前成為不依賴于特定引物擴增的環境病毒組研究的熱點領域[84-87]。與RAPD-PCR研究相同,環境病毒宏基因組學解析的前提條件是去除細胞生物遺傳基因在病毒遺傳物質中的污染。為此,研究者們在病毒基因提取、純化等方法學研究上開展了許多工作,包括利用不同孔徑微孔濾膜的切向過濾法(TFF)、核酸酶消化處理去除病毒濃縮液中細胞生物遺傳物質、10%聚乙二醇(PEG)溶液沉淀濃縮病毒顆粒、濃度為2.9 mg·L-1的FeCl3溶液作為病毒絮凝劑的絮凝-過濾-再懸浮方法(FFR)、20%蔗糖墊超離心純化法或不同密度CsCl溶液超離心純化法等[85,88-90]。需要指出的是,這些方法對環境病毒的分離過程的第一步均是采用將提取到的病毒溶液通過0.2 μm的微孔濾膜而富集,這一步驟無疑會將一些尺寸較大的病毒,如巨大病毒等排出在外[91]。2009年Nature Protocols上發表一篇文章,詳細介紹了針對水體樣品宏病毒基因組學研究的方法[90],該文研究策略可推廣應用到土壤病毒宏基因組解析,不同之處在于土壤有前處理,也就是從土壤中高效地分離提取出病毒溶液的過程。在開展環境宏病毒基因組學研究另一個需要注意的事項是提取到的病毒遺傳物質量的多少。雖然環境病毒數量明顯高于寄主生物,但由于提取效率低和病毒基因組小的原因,從常規的樣本用量(如水樣20 L,土樣10~50 g)中提取到的病毒遺傳物質量很少,不能滿足后續宏基因組研究的需要,需要經過體外擴增技術以增加病毒遺傳物質的濃度,達到測序的要求。目前常用的方法有兩種:一是多重置換擴增(MDA)技術,即采用φ 29 DNA聚合酶進行擴增技術[87,90];二是連接擴增(LASL)技術[92]。需要引起注意的是兩種擴增技術在擴增產物上存在偏好性,采用MDA傾向于擴增出ssDNA病毒,而采用LASL得到的病毒數據全部來自于dsDNA病毒[93]。其他研究土壤病毒宏基因組結果也證實MDA法傾向于獲得大量的ssDNA病毒[94-95]。這樣的結果與土壤中實際情況不符合,因為土壤病毒主要以噬菌體為主,且以dsDNA的有尾噬菌體占有較高的比例。為了避免體外擴增產生的偏好性,有研究者采用加大土壤樣品用量,甚至到幾千克,然后從土壤中大量富集病毒顆粒,提取病毒遺傳物質,直接進行宏基因組測序的方法[96-97]。需要指出的是,上述研究方法主要是針對DNA病毒,對于環境RNA病毒宏基因組研究則需要采取其他的技術,通常采用的是RP-SISPA(Random Priming-mediated Sequence-Independent Single Primer Amplification)基因擴增,然后再進行高通量測序[98-99]。圖2匯總了基于DNA水平對土壤病毒研究的主要方法及流程圖。

圖2 土壤病毒研究的主要方法及流程圖Fig. 2 Main methods used in researches on soil viruses and their flowcharts
制約環境病毒宏基因組研究的另一個瓶頸問題是對基因數據解析的困惑。與細菌等微生物數據庫相比,病毒基因數據庫非常匱乏,從而導致絕大多數環境病毒宏基因組序列缺少參比對象,基因來源不清,被劃分為未知基因,即使在剩下已比對上的基因中,也僅有非常小的一部分被確定為病毒,大部分基因與來源于細胞生物基因,尤其是細菌具有較高的同源性[84,87,100]。例如中國科學院生態環境研究中心賀紀正團隊對海灘和稻田土壤的病毒宏基因組分析發現,僅9.4%~40.5%的序列可以被注釋為已知的微生物信息,而在這些已知信息中被注釋為病毒的占到82%~97%[101]。Trubl等[86]對瑞典一個凍融地帶泥炭和沼澤土壤病毒宏基因組分析發現,盡管采用了嚴格的去除寄主遺傳物質的步驟,但僅19%的序列被鑒定為病毒。Williamson等[37]總結國際上8個土壤病毒宏基因組數據庫發現,有54.5%~97.3%的序列為未知來源和功能的序列。同樣,最近中國科學院生態環境研究中心韓麗麗團隊[102]對玉米根際與非根際土壤病毒宏基因組分析發現,僅0.26%~1.54%的序列被確認為病毒。大量源自病毒宏基因中的序列為未知基因,所以病毒也被喻為生物“暗物質”[57]。由此可見,高比例的環境病毒宏基因序列與已知的病毒數據不匹配,一定程度上妨礙了對病毒群落結構及生態功能的深入解析,這也反映出對環境病毒分離、純化和測序的重要性。只有基于病毒基因數據庫不斷擴大、完善基礎上,對環境病毒宏基因組解析才能夠更全面、更深入。目前國外學者相繼開發出多款數據分析軟件,如VIROME、VirSorter、Metavir和VirusSeeker等,用于環境病毒宏基因組數據解析[103-106]。
3 土壤病毒的生態功能
雖然土壤病毒數量巨大且廣泛存在,但我們對土壤病毒生態功能的認知尚處于初級階段,主要的功能理解多源自病毒基礎知識和海洋生態系統的相關研究結果,尚缺乏直接源自土壤中的實驗佐證。2018年德國哥廷根大學的Kuzyakov和Mason-Jones[15]撰文從5個角度對土壤病毒的生態功能給予概念性的描述:1)病毒路徑(Viral shunt):即土壤微生物被病毒感染裂解死亡,導致微生物中的碳氮磷硫等營養元素釋放出來被其他微生物和植物利用的過程。在海洋中由病毒驅動的碳循環量占海洋生態系統碳循環總量的6%~26%[33],在土壤環境中,營養元素循環在多大強度上受到病毒驅動還鮮見報道;2)促進土壤微生物快速周轉,保持活力狀態(Forever young):即由于土壤病毒的感染,導致土壤細菌快速更替,從而使存活的細菌始終處于年輕狀態,具有旺盛的代謝活力;3)病毒調節“胞外酶代謝”(Viral regulatory gate of EXOMET)。胞外酶代謝是指微生物細胞體內酶釋放到環境中,起到分解有機物質的作用。這個概念是2013年Maire等[107]首次提出的,他們發現經過γ-射線處理幾周的土壤已經沒有微生物存活,但這些土壤培養后還釋放出相當于未照射土壤處理的20%~60% CO2排放量。從這個現象中,我們不難理解土壤病毒感染微生物后,導致微生物裂解、胞內酶釋放到土壤中,間接地促進了土壤有機物分解轉化;4)通過微生物死體穩定性轉化,促進土壤固碳作用(C sequestration by microbial necromass stabilization)。土壤有機質是土壤碳庫穩定存在形式,Liang等[108]分析發現50%以上的土壤有機質來源于微生物死亡細胞,即通過微生物碳泵的續埋作用(Entombing effect)促進土壤固碳[109]。病毒導致微生物死亡,形成了大量的殘體碎片,可能促進了土壤有機質形成,起到固碳作用。需要說明的是,微生物細胞被病毒裂解后形成的死亡殘體與細胞受到非生物因素致死形成的殘體,哪種途徑更有利于促進土壤有機質形成尚無答案;5)引起土壤微域尺度C、N和P化學計量學分異(Microscale divergence of C/N/P stoichiometry)。這種作用主要是基于寄主細胞和病毒之間C、N和P化學計量學比值的差異,細菌病毒較細菌富含N和P,尤其是P元素。病毒感染細菌細胞裂解后,細菌細胞中有大約10%的碳、15%的氮和33%的磷營養轉變成病毒顆粒成分,不能被植物和其他生物利用,從而影響生態系統的養分循環。這種由于微生物和病毒之間元素化學計量差異而引起的磷營養限制僅在海洋環境中有報道[48],在土壤環境中,由于病毒生物量很小,以及土壤中養分含量相對較高的緣故,筆者推測病毒通過這種方式影響土壤養分循環的作用可能微乎其微。
上述土壤病毒功能主要是基于病毒基礎知識認知推測得來的。從病毒生態學角度來分析,土壤病毒還有以下幾方面功能:1)土壤病毒起到由上而下(Top-down)調控微生物群落結構的作用,從而間接地影響到土壤生態功能[110-111]。如向土壤中接種烈性根瘤菌噬菌體能夠降低根瘤菌數量,從而降低根瘤量[112];向土壤中接種青枯病菌噬菌體,通過噬菌體療法能夠降低作物青枯病的危害[113]等;2)土壤病毒還是基因水平移動的載體,病毒與寄主之間不斷地進行基因交流或突變,使得病毒和微生物獲得新的性狀和功能,共進化推動了地球生物群落不斷演替[46];3)土壤病毒基因組攜帶輔助代謝基因(Auxiliary metabolic genes,AMGs),協助寄主微生物驅動生物地球化學循環。如最近對紅樹林土壤和農田土壤研究均發現病毒宏基因組中含有多種水解多糖類物質的CAZymes基因,推測這些基因在促進土壤碳循環中起到重要作用[102,114];4)土壤病毒可通過介導抗性基因的水平轉移及調節宿主菌代謝通路來協助宿主抵抗低營養、輻射和污染等多種生態脅迫,對維持受脅迫生態環境中的微生物群落結構及功能穩定具有重要意義[115-116]。但目前關于低營養脅迫下的這一功能進行了較多研究,而針對污染環境(如農藥污染、石油污染和重金屬污染等)中,噬菌體協助宿主適應環境的相關研究還很少。圖3概況了以土壤病毒為中心,病毒起到的直接或間接生態功能。
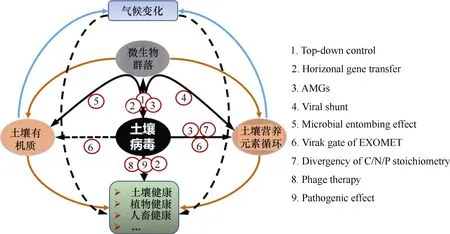
圖3 土壤病毒主要生態功能示意圖Fig. 3 Diagram of the main ecological functions of the viruses in soil
4 展 望
土壤中蘊藏著巨大的生物多樣性,僅以原核生物為例,其在土壤中的多樣性較其他生態系統的總和高出3個數量級[117-118],鑒于土壤微生物多樣性高和巨大的生態功能,世界許多國家相繼實施了一系列重要的土壤微生物組研究計劃,中國科學院也于2014年啟動了“土壤-微生物系統功能及其調控”戰略先導專項,取得了一系列重要發現和成果[119]。這些研究主要涉及到具有細胞結構的微生物,很少涉及土壤病毒。進入21世紀,海洋領域病毒生態學研究發展迅速,科學家們發現病毒尤其是噬菌體在海洋生態系統物質循環、能量流動,以及維持海洋生物多樣性和生物進化等方面起到重要的作用[31,52,120-121]。與海洋研究相比,針對土壤病毒研究卻進展緩慢[11,37]。鑒于土壤病毒存在狀態,以及土壤種類多樣性和土壤環境的高度異質性,土壤中病毒組成較海洋等水體環境更復雜、多樣性更高,研究的挑戰性也會更大[12]。目前世界各國科學家已意識到了土壤病毒的重要作用和研究的緊迫性,相繼發表了一些綜述性文章報道土壤病毒研究進展,呼吁學術界對土壤病毒研究的重視[11-13,15,37,122]。本文梳理了一些土壤病毒研究的最新進展,從這些研究結果看,我們對土壤病毒的認知還很有限,尤其是對土壤病毒生態功能展示方面還缺乏直接的證據,缺乏將土壤病毒群落與寄主群落演替耦合起來的研究策略。基于此,認為未來對土壤病毒研究應在以下方面給予加強。
4.1 重視對土壤RNA病毒的研究
目前多數研究結果是針對土壤DNA病毒,較少涉及到土壤RNA病毒,而許多侵染真核生物的病毒是RNA病毒,尤其是導致人、畜和家禽流行性疾病的病毒,以及絕大多數的引起植物病害的病毒都是RNA病毒,如引起新冠肺炎病毒、各種禽流感病毒、煙草花葉病毒和馬鈴薯Y病毒等。有關這些病原性的RNA病毒在土壤中生存狀態、遷移,以及活性受那些土壤因素影響,是否具有感染能力等問題還缺少系統性的研究。
4.2 關注對土壤溶原性噬菌體的研究
現階段對土壤病毒研究多采用基于微孔膜過濾富集病毒顆粒的方法,這種方式無疑將大部分溶原性噬菌體屏蔽在外,而土壤中溶原性噬菌體是土壤病毒主要存在形式,缺乏對溶原性噬菌體的研究,也限制了對土壤整體病毒多樣性及生態功能的認知。
4.3 關注土壤病毒資源的開發,提高土壤病毒研究的市場經濟價值
土壤病毒不僅是最大的基因寶庫,許多病毒資源也具有非常大的應用價值,如從土壤環境中篩選出能夠高效裂解動植物致病細菌的噬菌體資源,應用噬菌體療法防控動植物病害是一個非常有前景的研究領域[123-124]。此外,病毒還是天然的生物納米材料,其不但擁有納米材料的優越性能,還具備可快速復制的優勢,同時其遺傳物質較小,易于被基因工程技術改造,因此被越來越廣泛地應用到納米材料生成技術的各個方向,包括顯示屏和電池等[125-126]。
4.4 注重對微生物病毒純培養株的分離及基因組解析研究
目前針對病毒純分離株全基因組的解析量遠小于具有細胞結構的微生物,導致病毒全基因組數據庫量小,大量土壤病毒宏基因組數據無法判斷來源,妨礙了后續解析工作。因此在今后研究過程中,在采用分子生物學技術研究土壤病毒生態學問題的同時,也要關注微生物病毒純培養株全基因組分析工作,尤其是對古菌病毒和真菌病毒株的研究。這些基礎性的工作是擴大環境病毒數據庫的前提,是開發新分析軟件和建立新分析平臺的保障,也是開啟土壤病毒基因寶庫的鑰匙。
4.5 加強土壤病毒與土壤微生物的耦合研究
除部分土壤病毒能夠直接引起動植物病害和人類疾病外,土壤病毒主要是通過調控寄主微生物群落結構組成、影響群落演替而間接地體現出病毒的生態功能。目前國際上對土壤病毒的研究還局限于盤點其多樣性和數量豐度等層面,對病毒生態功能的研究還很少。多數對土壤病毒的研究未能與土壤微生物群落變化聯系起來,這無疑會妨礙我們對病毒生態功能的認知。故此,在今后的研究中,應加強對土壤病毒與土壤微生物的耦合研究,通過精巧的控制實驗設計,研究揭示出病毒在構建健康土壤環境、調控根際微生態、促進植物生長,乃至影響全球氣候變化等方面的重要作用。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
