兩種麻瘋樹核糖體失活蛋白基因Curcin和Curcin C在種子萌發過程中的表達模式研究
2021-01-28 03:21:08吉柔風丁蒙蒙
四川大學學報(自然科學版) 2021年1期
張 敬, 吉柔風, 楊 千, 丁蒙蒙, 陳 放, 徐 鶯
(四川大學生命科學學院 生物資源與生態環境教育部重點實驗室, 成都 610065)
最初,核糖體失活蛋白是從蓖麻種子中分離純化出來的,被人們命名為ricin[1].之后,隨著研究的不斷深入,已經在14個家族中發現了超過50種核糖體失活蛋白,研究較為清楚的有美洲商陸、麻瘋樹、苦瓜、相思子、絲瓜、樟樹等[2].植物核糖體失活蛋白已被證實具有N-糖苷酶活性、水解酶活性、DNA酶活性等酶學活性和抗真菌、抗病毒、抗腫瘤等生物學活性,關于其酶學活性研究的較為透徹和深入,而生物學活性的作用機制和機理,目前還沒有確切的結論[3].實驗研究表明,麻瘋樹核糖體失活蛋白Curcin具有抗腫瘤、抗真菌等生物學活性.Zhang等人從麻瘋樹子葉中提取出核糖體失活蛋白Curcin C,證明其具有N-糖苷酶活性、翻譯抑制作用、抗腫瘤活性.關于核糖體失活蛋白的作用機制,并沒有得到很清晰的驗證[4].
從麻瘋樹中分離出的核糖體失活蛋白多為Ⅰ型核糖體失活蛋白,Curcin是最早從麻瘋樹中分離出來,是由林娟等人根據前人的經驗,純化出Curcin,該蛋白分子量為28.2 kD,等電點為8.54,并驗證具有N-糖苷酶活性、翻譯抑制活性[5].之后魏琴等人發現了核糖體失活蛋白Curcin 2,Curcin L等與脅迫相關的蛋白,這種特殊蛋白的存在,可能會起到保護麻瘋樹的正常發育生長、抵御不良生理環境的作用[6-7].近年來,Zhang等人從麻瘋樹子葉中分離提取出麻瘋樹核糖體失活蛋白Curcin C,活性實驗表明,Curcin和Curcin C蛋白均具有抗腫瘤活性,但兩者抗腫瘤活性效率差異較大[4,8].植物核糖體失活蛋白可抑制蛋白質合成,也因其酶學活性和生物學活性,在生物醫學領域具有廣闊的應用前景[9].也有研究顯示,西昌地區的麻瘋樹種子核糖體失活蛋白含量最高,可作為核糖體失活蛋白應用的材料來源和研究的重要原材料.這些麻瘋樹核糖體失活蛋白的特性可能與其定位、生物學作用具有緊密的關聯,一直以來是研究的熱點和重點.
麻瘋樹在中國西南和東南地區有著豐富的資源,為了研究和開發這一資源,因此本研究以此為切入點,首先探討麻瘋樹種子萌發和幼苗生長過程的形態學的變化,為后續的研究奠定一定的基礎,其次從基因層面研究麻瘋樹核糖體失活蛋白的在種子萌發和幼苗生長過程中的一個動態變化,以及在各個組織器官中的表達情況,進而推測其在種子萌發和幼苗形成過程中可能發揮的作用.采用生物信息學以及構建含有GFP基因的瞬時表達載體等方法確定了這兩種蛋白的亞細胞定位信息.這些研究對于揭示其存在及發揮作用的機制具有重要的意義,同時也為這兩種蛋白質的結構和功能研究打下了基礎.
2.1 材 料
購買于四川省西昌市金沙江干熱河谷地區的麻瘋樹成熟種子.
2.2 方 法
2.2.1種子萌發 選取干粒質量達到0.68 g以上的麻瘋樹種子置于室溫條件下用水浸泡2 h,將蛭石和營養土等比例混合均勻,加入適量的水,保證濕潤.浸泡完畢后,將種子種入花盆中,萌發條件為30 ℃,光照/黑暗為18 h/6 h;定期澆水使得培養床保持一定的濕度;每天定時觀察并記錄萌發狀態,隨時準備收集各個時期的胚乳、子葉、胚根等樣品.
2.2.2Curcin、CurcinC基因表達分析 按照天根RNAprep Pure植物總RNA提取試劑盒說明書提取材料的總RNA,采用瓊脂糖凝膠電泳檢測核酸的完整度.利用PrimeScriptTMRT reagent Kit (Perfect Real Time) (TAKARA,大連)試劑盒合成單鏈cDNA.反應完成后cDNA產物-20 ℃保存備用.采用SYBR?Premix Ex TaqTMⅡ試劑盒對Curcin、CurcinC基因進行熒光定量PCR,每個樣品均重復三次以避免加樣誤差,實驗數據利用iQ5-Cycler 進行分析,分析方法為ΔΔCt 法.
2.2.3 Curcin、Curcin C蛋白表達分析 采用磷酸鹽緩沖液浸提丙酮沉淀法提取不同組織(胚根、胚軸、子葉、真葉)的總蛋白后進行SDS-PAGE、轉膜、封閉、孵育抗體以及顯色反應.
2.2.4 生物信息學分析 利用在線網站Cell-PLoc2.0(http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2/)對Curcin、Curcin C蛋白亞細胞定位進行預測,利用在線網站(http://www.cbs.dtu.dk/services/SignalP/對Curcin、Curcin C)蛋白信號肽進行預測.
2.2.5 農桿菌侵染煙草葉片表達Curcin/Curcin C-GFP融合蛋白 設計引物并構建pBI221-Curcin/pBI221-Curcin C重組載體,將重組載體轉入大腸桿菌并挑選陽性克隆進行測序,采用質粒大量提取純化試劑盒(Vigorous Plasmid Maxprep Kit)提取pBI221-Curcin/ Curcin C重組質粒,將其轉入農桿菌,挑選陽性單菌落轉入煙草中,注射后培養2~3 d,在共聚焦顯微鏡下進行熒光檢測.
2.2.6 原生質體轉化 參考Dr. Sheen's Lab(http://genetics.mgh.harvard.edu/sheenweb/protocols_reg.html)的操作手冊進行擬南芥原生質體制備,利用PEG介導的方式進行原生質體轉化,取出轉化后的原生質體于載玻片上,熒光共聚焦顯微鏡觀察.
2 結果
3.1 Curcin、Curcin C在種子萌發及幼苗生長過程中的動態變化
為了探究Curcin和CurcinC基因在胚乳和子葉中的表達量情況,我們采用了qRT-PCR實驗對Curcin基因和CurcinC基因在種子萌發及幼苗生長過程中各個時期的胚乳、子葉中的表達水平進行了定量分析,研究結果表明(圖1),Curcin基因在種子萌發過程中的胚乳中的表達呈現C1、C3期逐漸增加,C5、C7、C9開始出現下降,C13、C15、C17期最少,呈現出先增加后減少的趨勢.然而,Curcin在子葉中基本沒有表達[4].CurcinC基因則在種子萌發的初始階段胚乳中C3期表達量最高,隨著萌發過程的進行,表達量逐漸下降,在C15、C17期幾乎檢測不到.在子葉中,CurcinC基因的表達經歷了“從無到有,先上升后下降”的過程,CurcinC的轉錄產物首先在C3期出現,其表達量在C5時期達到最大值,隨后迅速下降,到C9期時,其水平不到最高期的10%,到C15期就低至到無法檢測的水平了.
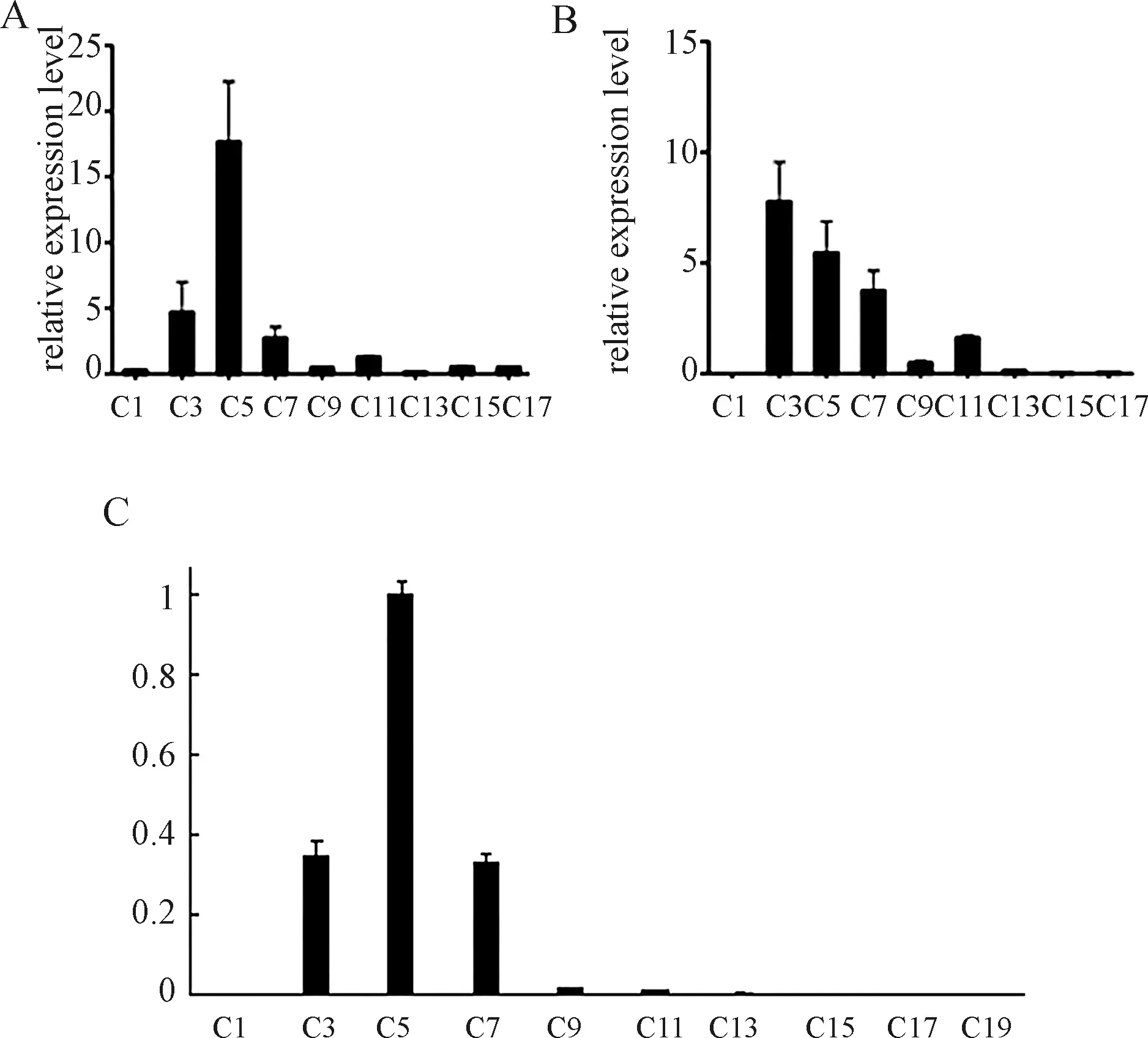
圖1 不同萌發時期Curcin,Curcin C基因在胚乳、子葉中的表達分析
3.2 Curcin、Curcin C在幼苗中的組織特異性表達
為了探究核糖體失活蛋白在幼苗中各個組織中的表達情況,我們收集了C11時期的幼苗,分別以該時期的根、下胚軸和真葉為對象進行熒光定量實驗和Western blot雜交鑒定.熒光定量結果顯示(圖2A),可以看出,在子葉、真葉組織中Curcin基因的表達量比CurcinC基因的表達量低,Curcin基因在這四種組織中的表達量都特別低;對于CurcinC基因來說,在真葉和子葉中的表達量很高,而在根和下胚軸中幾乎檢測不到.Western雜交實驗顯示(圖2B、C),Curcin蛋白在根、下胚軸、子葉和真葉中幾乎不表達;Curcin C蛋白則在根、下胚軸中不表達,而在子葉和真葉中表達,且在子葉中表達量高于真葉的表達量.熒光定量實驗檢測Curcin、CurcinC基因的表達情況與Western實驗檢測結果基本一致.
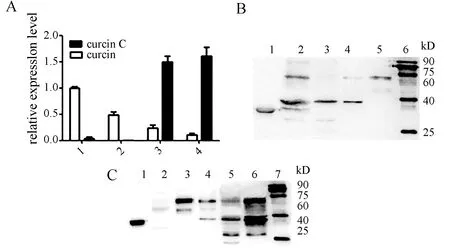
圖2 Curcin, Curcin C組織特異性表達分析
3.3 Curcin和Curcin C的亞細胞定位研究
為了進一步確定Curcin 、Curcin C蛋白在亞細胞結構上的定位,進而研究其在種子萌發和幼苗生長中所具有的功能.我們首先利用在線網站(http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2/、http://www.cbs.dtu.dk/services/ SignalP/) 對Curcin、Curcin C蛋白信號肽、亞細胞定位進行預測,結果顯示兩種蛋白都含有信號肽且都定位在細胞壁上.為了對預測結果進行驗證,我們分別構建了Curcin:GFP和CurcinC:GFP融合蛋白的植物表達載體,以35S:GFP為對照載體,分別用根癌農桿菌介導侵染煙草葉片法和原生質體轉化法讓其在植物細胞中瞬間表達,在激光共聚焦顯微鏡下檢測熒光信號,確定亞細胞定位信息.實驗結果如下:陽性對照組為圖3A(g、h、i),可以看到在煙草表皮細胞的細胞膜(壁),細胞核,細胞質中都有表達,Curcin、Curcin C蛋白的結果分別如圖3A(a、b、c和d、e、f),在細胞膜(壁)上可以看到熒光信號,由此可以初步的確定這兩種蛋白定位在細胞壁或者細胞膜上.之后進行亞細胞定位實驗,結果如下,陽性對照如圖3B(a、b、c、d),在細胞質中檢測到了熒光信號,而對于實驗組(圖3B(e、f、g、h和i、g、k、l))未檢測到熒光信號.結合這兩種方法的實驗結果,可以從側面間接的說明這兩種蛋白可能定位在細胞壁上,通過分泌到細胞外發揮相應的生物學作用.該結果與網站預測的結果基本一致.
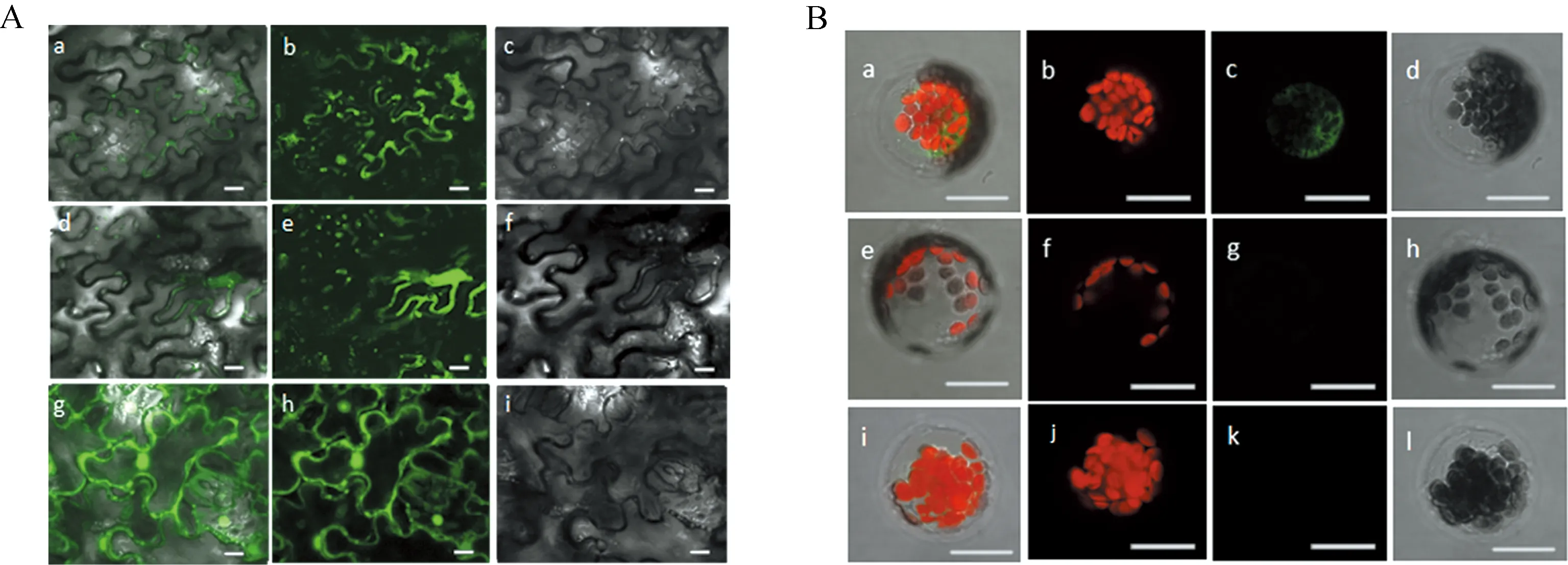
圖3 Curcin, Curcin C蛋白亞細胞定位分析
4 討 論
Curcin、Curcin C蛋白是分別從麻瘋樹的種仁和子葉中提取分離純化出來的,為了更深入的了解這兩種蛋白的生物學功能,本文首先對麻瘋樹種子萌發和幼苗生長過程中各個組織器官的動態變化進行了相關的研究,通過收集不同萌發時期的胚乳和子葉材料,探討了麻瘋樹種子萌發和幼苗生長過程中Curcin、CurcinC基因在胚乳和子葉中的表達情況,并推測其在萌發及幼苗生長中所發揮的生物學作用,進而加快探索核糖體失活蛋白在生物體內存在意義的進程.之后對Curcin、Curcin C蛋白的組織特性了進行了相關的研究,研究發現,Curcin基因在這四種組織中的表達量都特別低,可以認為幾乎不表達,結合Curcin基因在種子萌發中的動態變化的研究,以及Curcin 蛋白的抗真菌、抗病毒活性等,由此推測,該蛋白可能是在種子萌發過程中,既作為一種貯藏蛋白,為種子后續的萌發提供營養物質,又作為一種防御性蛋白維護種子的正常萌發起到一定的作用.Curcin C蛋白在真葉和子葉中都有表達,由此推測它可能是一種誘導性蛋白,在發育過程、脅迫,病毒感染等生物學過程中表達,起著保護植物正常生長的作用.
通過對核糖體失活蛋白Curcin、Curcin C的生物信息學的分析得出這兩種蛋白質的亞細胞定位預測及信號肽的分析,對于后續的對這兩種核糖體失活蛋白的研究奠定相應的基礎.蛋白質在亞細胞結構中的定位,直接影響著其在生物體內功能作用的發揮,為了進一步探究兩種核糖體失活蛋白的定位,之后通過將核糖體失活蛋白Curcin、CurcinC基因構建綠色熒光蛋白表達載體,注射煙草和轉化擬南芥原生質體后,采用激光共聚焦顯微鏡觀察這兩種蛋白的亞細胞結構的定位,綜合兩種實驗結果表明,這兩種蛋白可能定位在細胞壁上.
文獻研究結果表明:Ⅰ型核糖體失活蛋白在亞細胞結構中定位主要有:細胞間隙處(dioicin 1、dioicin 2、Gypsophilin)、細胞壁和細胞膜之間的擬壁體(皂草素)以及葉肉細胞的細胞壁基質(PAP)[10-13].眾所周知,核糖體失活蛋白對核糖體具有一定的抑制作用,而這種特殊的定位分離了蛋白與自身細胞核糖體,使其自身細胞不會受到核糖體失活蛋白的毒害作用[14].并且,一般來說膜定位的蛋白質一般會存在至少一個跨膜結構域,通過對這兩種蛋白進行跨膜結構域的預測,發現并未含有跨膜結構域,結合相關文獻及實驗結果,從而推測這兩種蛋白定位在細胞壁上,通過分泌到胞外來發揮相應的生物學功能.
