14C-吡蟲啉在苗期油菜中的吸收、轉運與分布特性
2021-01-28 01:37:48李成名聶恩光張素芬葉慶富
核農學報 2021年2期
李成名 聶恩光 張素芬 葉慶富
(浙江大學原子核農業科學研究所/農業部核農學重點實驗室,浙江 杭州 310029)
新煙堿類殺蟲劑作用于昆蟲的乙酰膽堿受體,擾亂其神經活動,對害蟲有觸殺和胃毒作用,其作用機制與天然煙堿類似[1-2]。吡蟲啉(Imidacloprid)作為全球首個商品化的新煙堿類殺蟲劑,迄今已在120 余個國家、數百種作物上登記使用[3-4]。盡管吡蟲啉已投入使用30 余年,但人們對其環境行為的認識仍不夠客觀、全面[5-6]。2007年,Cox-Foster 等[7]首次發現,以吡蟲啉為代表的新煙堿類殺蟲劑會引發“蜂群崩潰綜合癥”。2012年,Henry 等[8]和Whitehorn 等[9]又相繼報道,新煙堿類殺蟲劑會減弱蜜蜂覓食能力并降低蜂群增長速率及蜂王的繁殖能力。另有研究表明,新煙堿類殺蟲劑吡蟲啉在水域環境中分布廣泛[10],且對水生及陸生無脊椎動物(如底棲動物、蜉蝣稚蟲、甲殼蟲和蒼蠅等)具有致死作用[5,11-12]。Hallmann 等[13]研究發現,新煙堿類殺蟲劑吡蟲啉可能通過營養級聯效應引發自然生態系統災害,如在吡蟲啉嚴重污染的水環境區域,無脊椎昆蟲大量死亡,并在營養級聯效應的作用下,食蟲鳥類種群和數量也急劇下降。為了遏止新煙堿類農藥對生態環境的持續性危害,歐盟決定從2013年12月1日起對吡蟲啉、噻蟲嗪和噻蟲胺實施限用政策,美國、加拿大和巴西等國家也相繼實行禁限令或展開相應的審查和復審工作。自2018年9月1日,法國開始在全國范圍內禁止使用新煙堿類殺蟲劑。目前,吡蟲啉仍是國內使用頻度較高的殺蟲劑之一[14-16],因此,研究吡蟲啉在農業生態系統中的行為規律對其科學使用具有重要的現實意義。
油菜(Brassica napusL.)是世界四大油料作物之一。據國家統計局數據顯示,2017年我國油菜籽播種面積665.30 萬hm2,油菜籽產量約為1 327.41 萬t[17]。在油菜生產中,油菜蚤跳甲和油菜黃曲條跳甲等蟲害時常威脅油菜的生長,吡蟲啉可有效抑制該蟲害的發生[18]。然而,目前國內外主要側重于吡蟲啉環境毒理以及殘留監控等方面的研究,而有關其在油菜中的吸收、運轉與殘留定向積累等方面的研究尚鮮見報道。本試驗以14C-吡蟲啉為示蹤劑,采用種衣劑拌種方式處理油菜種子,研究14C-吡蟲啉在苗期油菜植株中的吸收、運轉與殘留分布特征,以期為客觀評價吡蟲啉的環境安全和農產品質量安全提供科學依據。
1 材料與方法
1.1 試驗材料
供試標記化合物14C-吡蟲啉,購自上海啟甄環境科技有限公司。放射化學純度為97.35%,化學純度為98.03%±0.10%,比活度為6.54×105Bq·mg-1。化學結構式如圖1 所示。供試種衣劑為60%高巧懸浮種衣劑(有效成分含量600 g·L-1),登記證號LS99041(FS600),購自拜耳作物科學(中國)有限公司。供試油菜品種為甘藍型油菜浙大619,由浙江大學農業與生物技術學院周偉軍教授提供。
本試驗選用中性黃松土,取自浙江省杭州市浙江大學校內,土壤有機質含量30.5 g·kg-1,pH 值7.02,陽離子交換量(cation exchange capacity,CEC)10.83 cmol·kg-1,粘粒8.0%,粉粒71.3%,砂粒20.8%。
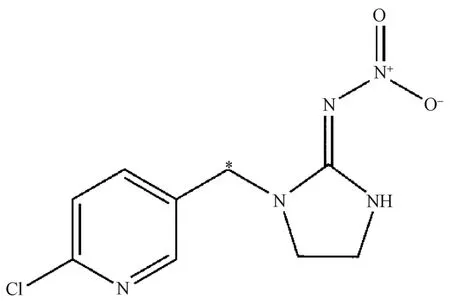
圖1 14C-吡蟲啉化學結構(*表示14C 標記位置)Fig.1 Chemical structure of 14C-Imidacloprid(* represents the position of 14C)
1.2 試驗設計
1.2.114C-吡蟲啉不同藥種比種衣劑的配制與引入 參考Laurent 等[19]和黃芳等[20]吡蟲啉包衣處理油菜種子的方法,并作適當改進。拜耳公司對油菜種子的吡蟲啉種衣劑推薦劑量范圍是4.8~7.2 g·kg-1。本試驗設計的供試藥種比均在拜耳公司推薦的田間使用量范圍內,分別設置了T1(4.8 g·kg-1)、T2(5.4 g·kg-1)、 T3(6.0 g·kg-1)、T4(7.2 g·kg-1)共4 個藥種比,同時以不添加吡蟲啉的油菜種子作為空白對照。取0.5 mL 60%高巧懸浮種衣劑,加水定容至5 mL,配成母液A,有效成分為60 mg·mL-1;另采用比活度為6.54×105Bq·mg-1的14C-吡蟲啉原藥和母液A,制備出有效成分濃度為60.07 mg·mL-1、比活度為2.01×105Bq·mg-1的14C-吡蟲啉母液B。分別吸取40、45、50 和60 μL14C-吡蟲啉母液B,加水定容至500 μL。每組稱取0.5 g 油菜種子(約114 粒)于1 mL 離心管中進行拌種,拌種時用塑料薄膜包裹,防止藥液揮發,振蕩混合均勻,超聲3 min 后,陰干備播。
1.2.2 苗期油菜的培養與取樣 取陰干后的油菜種子播于裝有120 g 土的塑料杯(200 mL)中,每杯播種1 粒。模擬田間條件培養,至三葉始期第一次取樣,而后每隔7 d 取樣一次,每次取樣6 個重復,共取樣5次。
1.3 測定項目與方法
將油菜整株取出,用自來水沖洗干凈附著土壤后,將植株按根、莖、葉分離處理,葉片自下往上記為葉1、葉2、葉3、葉4。將樣品于60℃烘箱中烘干至恒重并分別記錄其干重。取約1 g 干樣于生物氧化燃燒儀中,放射性物質氧化燃燒后轉化為14CO2,經15 mL 閃爍液B(閃爍液B 配方:28 g 2,5-二苯基惡唑,2 g 1,4-雙[2-(5-苯基惡唑)],2.4 L 二甲苯,0.9 L 乙二醇乙醚,超聲0.5 h 溶解后,加入0.7 L 乙醇胺)。吸收靜置后,采用Tricarb-2910TR 液體閃爍測量儀(美國Perkin Elmer 公司)測定14C 放射性活度,并計算吡蟲啉殘留在各樣品中的含量。另外,于三葉始期后21 d時每處理另取1 株油菜幼苗,殺青、烘干、壓片處理,暗室下曝光37 d 后,采用Typhoon FLA 9500 同位素磷屏與多功能激光掃描成像系統(美國General Electric 公司)進行放射性自顯影成像。
根據公式計算14C-吡蟲啉吸收利用率、14C-吡蟲啉分配比、14C-吡蟲啉含量和14C-吡蟲啉轉運系數:
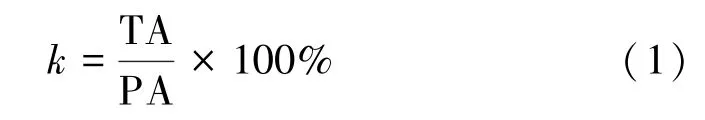
式中,k為14C-吡蟲啉吸收利用率,%;PA 為拌種后每粒油菜種子測定的放射性活度,Bq;TA 為植物體內總放射性活度,Bq。
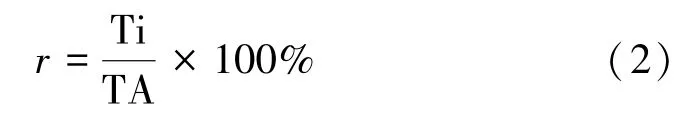
式中,r為14C-吡蟲啉分配比,%;TA 為植物體內總放射性活度,Bq;Ti 分別為根、莖、葉中的放射性活度,Bq。

式中,c為14C-吡蟲啉含量,mg·kg-1;Ai 為干樣中的放射性活度,Bq;a為母液B 中吡蟲啉的放射性比活度,2.01×105Bq·mg-1;Mi 為干樣質量,kg。

式中,TF 為14C-吡蟲啉轉運系數;Cs 為植物體內地上部吡蟲啉含量,mg·kg-1;Cr 為植物體地下部的吡蟲啉含量,mg·kg-1。
1.4 數據分析
數據采用Excel 2016 進行處理,并用Origin 9.1作圖,采用Duncan’s multiple range test 法進行顯著性分析。
2 結果與分析
2.1 油菜種子中14C-吡蟲啉種衣劑的包覆率分析
農藥包覆率體現了該試驗條件下農藥包衣處理種子的藥液利用水平。各處理組的種衣劑包覆率如表1所示。結果表明,每粒種子的包覆量隨處理濃度的升高而增加,藥種比為4.8、5.4、6.0、7.2 g·kg-1時,每粒種子中14C-吡蟲啉種衣劑的包覆量分別為1.18、1.41、1.52、1.81 kBq/粒;各處理組14C-吡蟲啉種衣劑對油菜種子的包覆率在27.83%~29.46%范圍內。
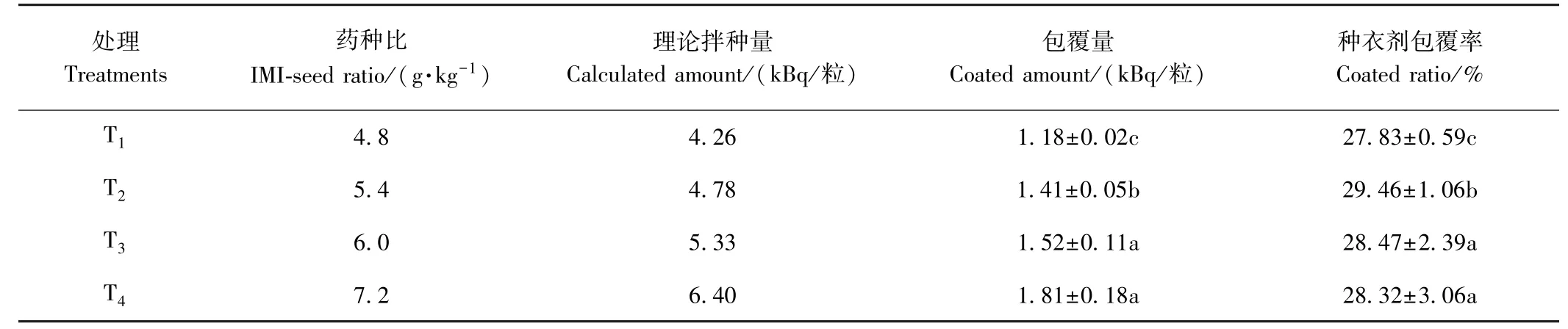
表1 14C-吡蟲啉種衣劑對油菜種子的包覆率Table 1 Coated ratio of 14C-Imidacloprid on rapeseeds by seed coating
2.2 苗期油菜體內14C-吡蟲啉的吸收和殘留動態
苗期油菜對14C-吡蟲啉的吸收利用率隨時間的變化如圖2 所示。結果表明,苗期油菜對14C-吡蟲啉的吸收利用率呈現T3>T4>T2≈T1的規律。在整個培養過程中,低藥種比T1和T2對14C-吡蟲啉的平均吸收利用率分別為9.32%和8.55%,二者之間無顯著差異(P>0.05)。高藥種比T3和T4對14C-吡蟲啉的吸收利用率均顯著高于T1和T2(除T4的28 d 外)(P<0.05),T3對14C-吡蟲啉的吸收利用率在20.20%~28.92%范圍內,T4在14.50%~19.17%之間。此外,整個試驗過程中,T3對14C-吡蟲啉的吸收利用率均顯著高于其他3 個處理組,即藥種比6.0 g·kg-1處理時,苗期油菜對吡蟲啉的吸收效率最高。
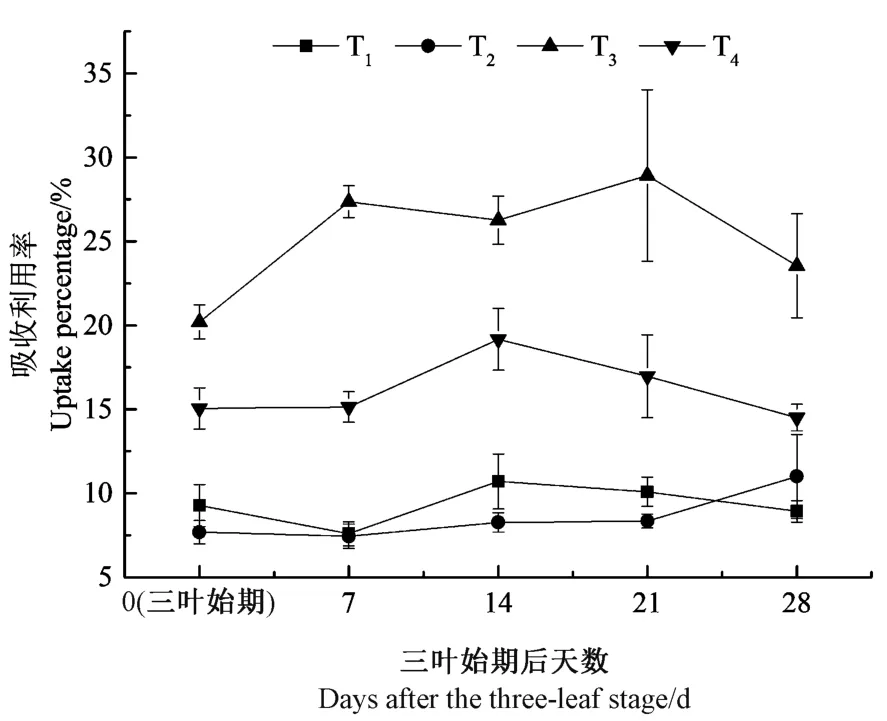
圖2 苗期油菜對14C-吡蟲啉吸收利用率的動態變化(n=6)Fig.2 Dynamics of uptake percentage of 14C-Imidacloprid in rape seedlings (n=6)
苗期油菜植株內的14C-吡蟲啉含量隨時間變化如圖3 所示。統計分析表明,在三葉始期,T2、T3和T4植株內的14C-吡蟲啉含量均顯著高于T1(P<0.05),而T3與T2之間差異不顯著,T4顯著高于T2(P<0.05);隨后的培養時間內,植株體內的14C-吡蟲啉含量表現為高藥種比T3和T4顯著高于低藥種比T2和T1(P<0.05),而2 個高藥種比處理之間無顯著性差異,且兩處理植株內的14C-吡蟲啉含量均隨培養時間延長而下降,分別由7.96 和9.42 mg·kg-1(0 d)下降至4.72 和4.20 mg·kg-1(28 d)。此外,在三葉始期后0 ~14 d 期間,T2植株體內的殘留含量極顯著高于T1(P<0.01),而在三葉期后21 和28 d,植株體內的14C-吡蟲啉含量表現為T2和T1之間差異不顯著,二者均維持在相對較低的水平(1.69 ~1.97 mg·kg-1)。植株體內14C-吡蟲啉的殘留量水平與其殺蟲能力有關,試驗結果顯示,相對較高藥種比T3和T4處理的殺蟲能力顯著高于較低藥種比T1和T2,且藥種比6.0 mg·kg-1處理(T3)能達到與藥種比7.2 mg·kg-1處理(T4)相近的殺蟲能力。
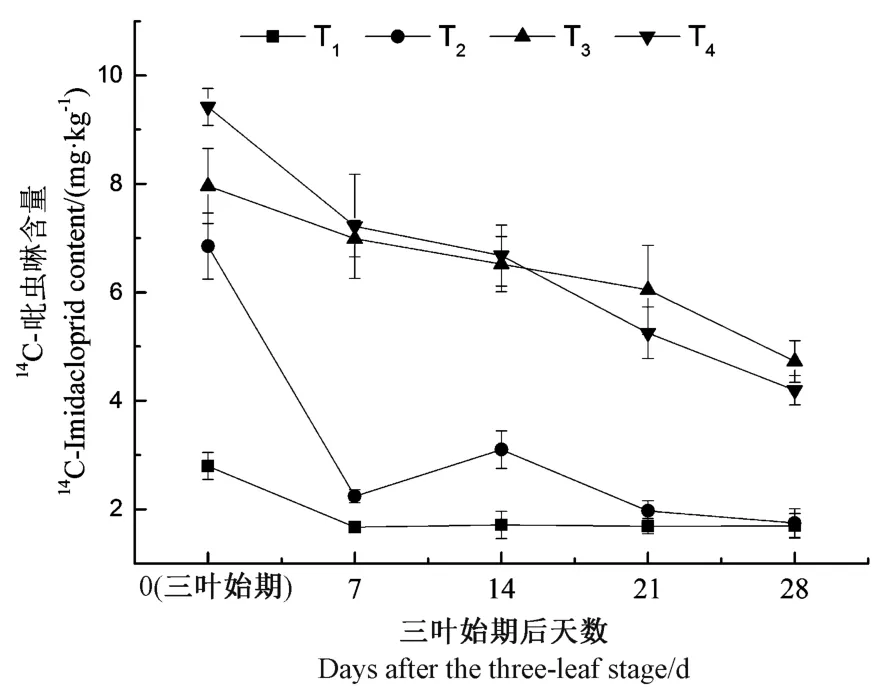
圖3 苗期油菜植株中14C-吡蟲啉含量的動態變化(n=6)Fig.3 Dynamics of 14C-Imidacloprid content in rape seedlings (n=6)
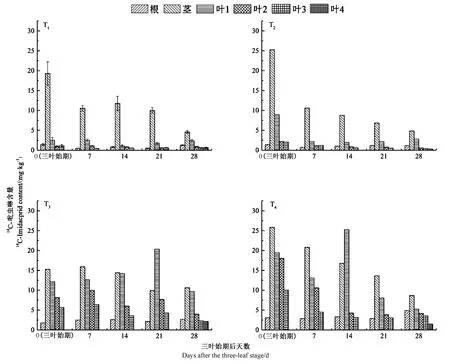
圖4 苗期油菜不同部位的14C-吡蟲啉含量及分布Fig.4 Distribution of 14C-Imidacloprid content in different parts of rape seedlings
2.3 苗期油菜不同部位14C-吡蟲啉的含量和分布的動態變化
油菜根、莖、葉中的14C-吡蟲啉含量及分布動態如圖4 所示。低藥種比T1和T2莖中的14C-吡蟲啉含量顯著高于植株其他部位(P<0.05),其最大值(T1為19.29 mg·kg-1,T2為25.26 mg·kg-1)均出現在三葉始期,最小值(T1為4.60 mg·kg-1,T2為4.84 mg·kg-1)均出現在三葉期后28 d,且下部葉(葉1)的14C-吡蟲啉含量高于其他葉。T1的莖中14C-吡蟲啉含量在三葉期后7~21 d 變化不顯著(P>0.05),隨后明顯下降,而T2的莖中14C-吡蟲啉含量在三葉期后7~14 d 變化較小,三葉期后14 d 開始顯著下降(P<0.05)。與低藥種比T1和T2相比,高藥種比T3和T4的14C-吡蟲啉含量呈現不同的分布規律,其莖和葉1 中的14C-吡蟲啉含量均顯著高于其他部位(P<0.05),在整個培養周期中的各取樣時間點,T3和T4葉片中14C-吡蟲啉含量均顯著高于T1和T2相應的葉片(P<0.05),說明高藥種比種衣劑處理時,苗期油菜吸收的吡蟲啉更易在葉片(尤其是下部葉片)積累,這有利于控制苗期油菜葉片的蟲害。除T3三葉始期外,T3和T4各取樣時間點苗期油菜根部的含量的14C-吡蟲啉含量均顯著高于T1和T2(P<0.05)。
由圖5 可知,高藥種比T3和T4的分配比與低藥種比T1和T2呈現出不同的分布特征。具體來看,T1和T2的分配比規律為:莖>葉>根,而高藥種比T3和T4的分配比規律則為:葉>莖>根(T4在三葉期后28 d除外)。各處理組根中14C-吡蟲啉的分配比均在9.81%~16.37%范圍內。
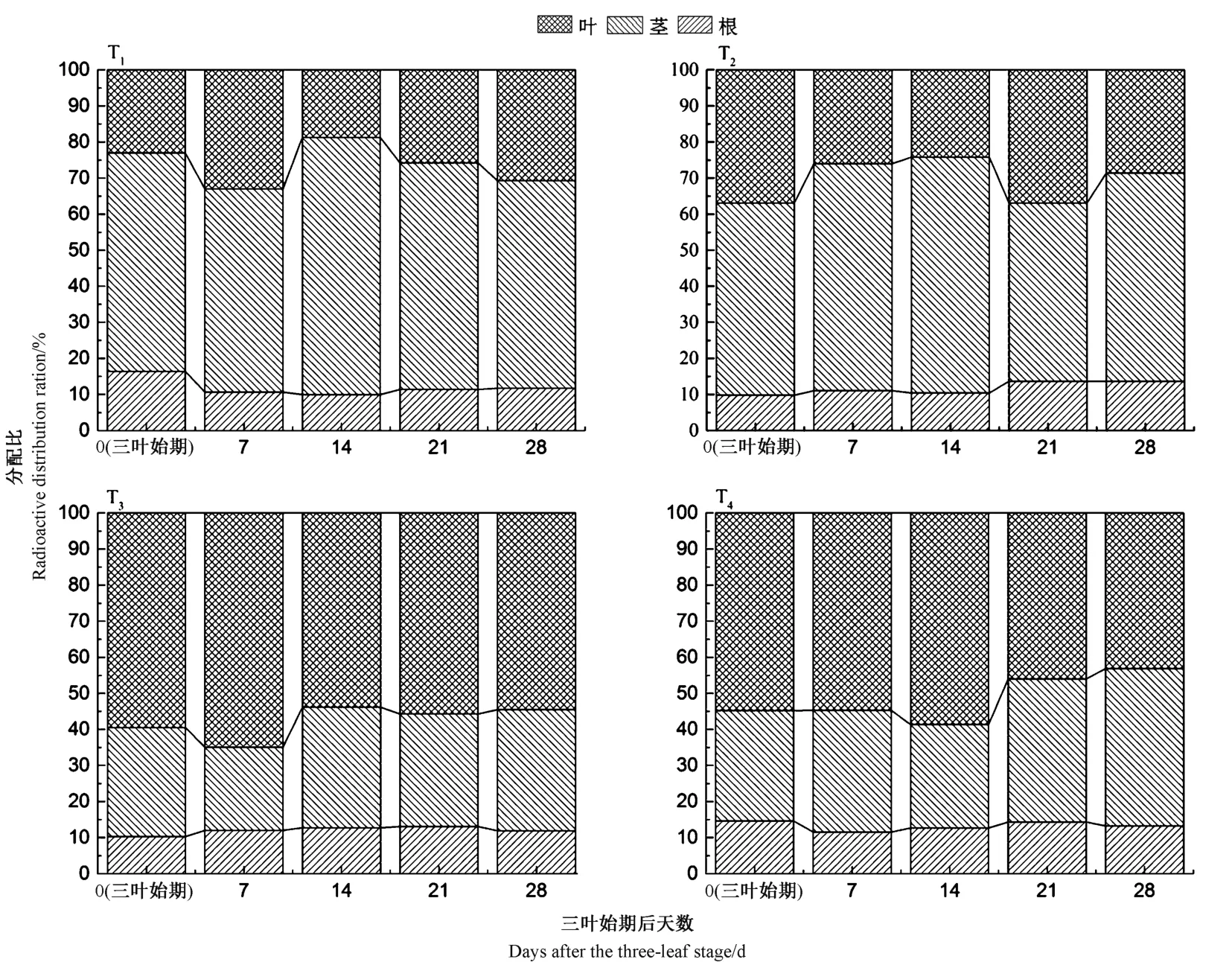
圖5 苗期油菜根、莖、葉中14C-吡蟲啉的分配比(n=6)Fig.5 Distribution ratio of 14C-Imidacloprid in roots,stems and leaves of rape seedling (n=6)
2.4 14C-吡蟲啉在苗期油菜植株中的放射自顯影
三葉始期后21 d 時,油菜植株的放射自顯影成像如圖6 所示。可見,不同藥種比處理植株體內14C-吡蟲啉及其代謝產物主要分布在莖和葉片,根中相對較少,且高藥種比處理油菜的根、莖、葉中14C-吡蟲啉含量均高于低藥種比,這與前文定量分析結果可相互印證。放射自顯影結果還表明,苗期油菜的葉緣部位的14C-吡蟲啉(包括代謝產物)含量相對于葉片其他部位更高,即14C-吡蟲啉更易在葉緣部位定向積累。
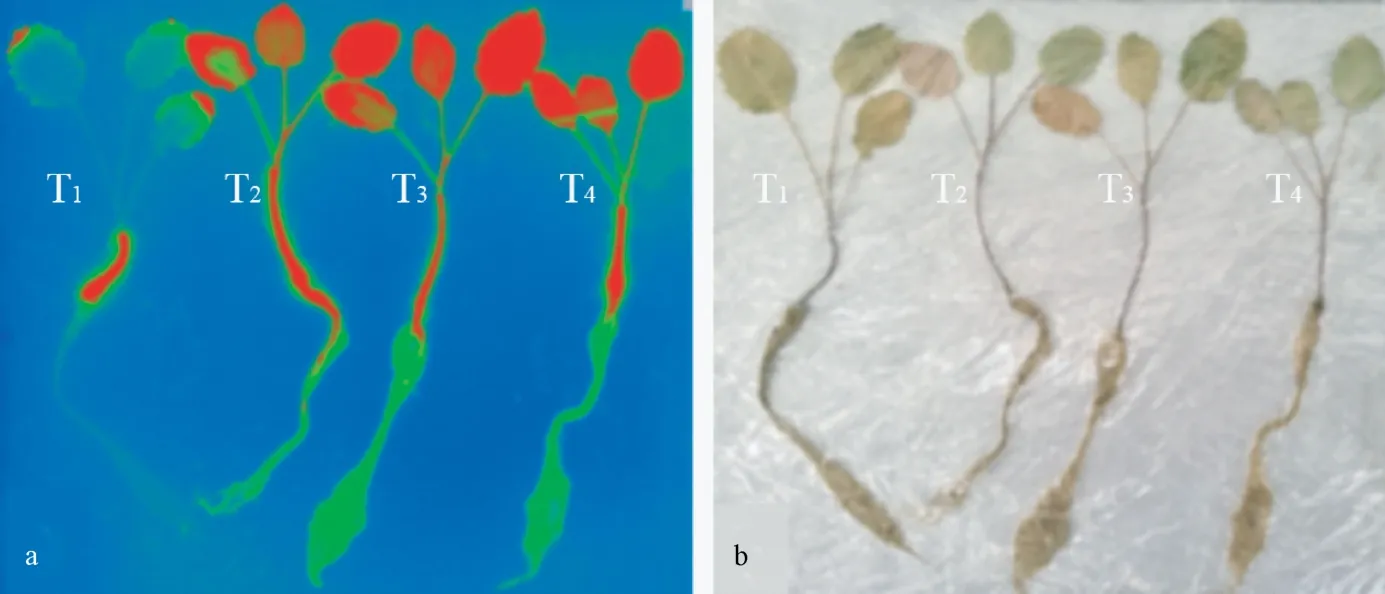
圖6 三葉期后21 d 苗期油菜的放射自顯影圖(a)和植株實物圖(b)Fig.6 Autoradiography (a) and physical map (b) of rape seedlings on the 21th day after the three-leaf stage
2.5 14C-吡蟲啉在苗期油菜中的轉運分析
14C-吡蟲啉在苗期油菜中的轉運系數動態變化如表2 所示。結果顯示,在整個試驗過程中,14C-吡蟲啉在油菜幼苗中的轉運系數均大于1,表明在種衣劑拌種處理油菜種子方式下,吡蟲啉容易通過根部向地上部轉運,且隨著培養時間延長,各處理組14C-吡蟲啉的轉運系數逐漸降低。從三葉始期至采樣終點,T1、T2、T3和T4的吡蟲啉轉運系數分別下降了63.38%、74.82%、60.84%和80.65%。在三葉始期,各處理組之間的轉運系數無顯著差異,說明三葉始期時,油菜幼苗對14C-吡蟲啉的轉運系數與藥種比之間無明顯相關性。就高藥種比T3和T4而言,在三葉始期(0 d)至三葉期后7 d,兩處理的14C-吡蟲啉轉運系數無顯著差異,而隨著培養時間延長,T3的轉運系數均顯著高于T4。

表2 14C-吡蟲啉在苗期油菜中的轉運系數動態變化Table 2 Dynamics of transport factors of 14C-Imidacloprid in rape seedlings
3 討論
3.1 吡蟲啉種衣劑處理甘藍型油菜種子的適宜藥種比
新煙堿類殺蟲劑吡蟲啉通過與昆蟲乙酰膽堿受體結合,干擾昆蟲神經系統進而達到殺蟲防害的效果[21],其對昆蟲的半致死量(lethal dose 50%,LD50)為2.0 mg·kg-1[22]。探索化學農藥的高效利用機理以及開發農藥減量增效技術是我國當前的研究熱點之一[23]。在推薦劑量范圍內,確定更適宜的藥種比以減少其用量,進而減少化學農藥污染,降低農藥用藥成本,具有重要的現實意義。
本試驗表明,吡蟲啉種衣劑拌種處理油菜種子,相較于T1、T2和T4,藥種比6.0 mg·kg-1處理(T3)的苗期油菜對吡蟲啉的吸收利用率最高。同時,苗期油菜植株體內的吡蟲啉總含量水平表現為高藥種比T3和T4顯著高于低藥種比T2和T1(除三葉始期外)(P<0.05),而高藥種比處理間無顯著性差異(P>0.05),兩者吡蟲啉含量水平分別維持在4.72 ~7.96 mg·kg-1和4.20~9.42 mg·kg-1范圍內;苗期油菜葉中高藥種比的吡蟲啉含量水平也顯著高于低藥種比(P<0.05)。植株體內的吡蟲啉含量與其蟲害防御能力有關,高藥種比處理的植株整體含量及葉中的含量均高于低藥種比,這意味著高藥種比處理有利于控制油菜植株整體尤其是葉片的蟲害。此外,在取樣后期,藥種比6.0 g·kg-1處理(T3)14C-吡蟲啉在油菜體內的轉運系數均顯著高于藥種比7.2 g·kg-1處理(T4)。綜上所述,藥種比6.0 g·kg-1為吡蟲啉種衣劑處理油菜種子的適宜藥種比。
3.2 吡蟲啉在植物體內的轉運及分布差異性的可能機理
本研究表明,油菜種子經高藥種比處理,其植株體內14C-吡蟲啉含量和放射性分配比均呈現出與低藥種比處理不同的特征。低藥種比處理,莖中吡蟲啉含量水平顯著高于植株其他部位(P<0.05),且下部葉(葉1)高于其他葉,其分配比規律為:莖>葉>根。而高藥種比處理,莖和葉1 中吡蟲啉含量均顯著高于其他部位(P<0.05),其分配比規律則為:葉>莖>根(T4三葉期后28 d 除外)。各處理組根中14C-吡蟲啉的分配比均較低,在9.81%~16.37%范圍內。Laurent 等[19]研究表明,向日葵種子經過吡蟲啉拌種后,約10%進入到種子中,下部葉片吡蟲啉含量高于上部葉片。Ge等[24]有關吡蟲啉在水稻植株中吸收與運轉的研究表明,吡蟲啉從土壤引入水稻后,葉片的吡蟲啉含量高于根部。Sun 等[25]研究也表明,水培條件下玉米植株體內吡蟲啉的分布也呈現出葉>莖>根的規律。這些研究報道與本試驗研究結果類似。
由本研究結果可知,吡蟲啉在苗期油菜中的轉運系數均大于1,表明在種衣劑拌種處理油菜種子方式下,吡蟲啉易通過根部向地上部轉運。農藥在植株體內各部位分布差異可歸因于多因素的影響。農藥分子的結構、正辛醇/水分配系數(logKow)、溶解性等理化性質對其內吸性能影響較大[26]。通常認為,當logKow小于4 時,農藥分子在植物的木質部傳導能力較強,容易向上運輸;當logKow 大于4 時,農藥分子親脂性較強,不易向上運輸[27]。吡蟲啉的logKow 為0.57[28],親水性較強,因而易通過植株木質部向地上部運輸。這可能是本研究中吡蟲啉地上部含量高于根部的原因之一。
植物的蒸騰作用是農藥分子在植物體內吸收和轉運另一個重要因素[29]。已有研究表明,內吸性殺蟲劑均能不同程度的透過角質層到達植物內部,而后隨著植物的蒸騰作用到達植株各部位[30],由于葉片邊緣布滿氣孔,植物體內的農藥分子可在蒸騰流的作用下運輸至葉片邊緣。本試驗放射自顯影結果進一步證實,14C-吡蟲啉(包括代謝產物)主要分布在莖和葉中,尤其是葉緣部位,即14C-吡蟲啉更易在葉緣部位定向積累。Tapparo 等[31]用吡蟲啉處理玉米種子時也發現,植株吐水時其葉尖端液滴中含有吡蟲啉。可見,吡蟲啉易在葉緣部位積累這一現象符合蒸騰作用影響植物對農藥分子吸收和轉運的規律。
種衣劑拌種處理油菜種子后,隨著培養時間延長,苗期油菜對吡蟲啉的轉運系數逐漸降低。楊麗旋[32]在0.5 mg·L-1吡蟲啉水培條件下處理上海青和紫金香妃青菜,發現隨著培養時間延長,其轉運系數分別從1.91 和2.77 降至1.81 和1.14,這與本試驗的研究結果類似。究其原因,可能是隨著培養時間的延長,油菜植株生物量的增加速率超過了吡蟲啉在地上部的累積速率,從而表現出轉運系數隨時間延長而下降的現象。
4 結論
本試驗研究了14C-吡蟲啉在苗期油菜中的吸收、轉運和分布規律,結果表明,吡蟲啉種衣劑處理油菜種子的適宜藥種比為6.0 g·kg-1;低藥種比4.8 和5.4 g·kg-1處理,14C 吡蟲啉在各部位的分配比規律為:莖>葉>根;高藥種比6.0 和7.2 g·kg-1處理時,其分配比規律則為:葉>莖>根;吡蟲啉更易在葉緣部位定向積累;吡蟲啉在油菜幼苗中的轉運系數均大于1,且吡蟲啉的轉運系數隨著油菜培養時間延長而降低。本研究為農業生產上科學使用吡蟲啉和客觀評價吡蟲啉安全性提供了數據支撐和理論依據。
