低鎘水稻研究進展
2021-01-28 02:23:48胡婉茵吳殿星舒小麗
核農學報 2021年1期
關鍵詞:水稻
胡婉茵 王 寅 吳殿星 舒小麗
(1浙江大學原子核農業科學研究所,浙江 杭州 310029;2浙江省農業科學院農村發展研究所,浙江 杭州 310021;3農業農村部創意農業重點實驗室,浙江 杭州 310021)
鎘(cadmium,Cd)是一種二價陽離子,通過人類活動釋放到環境中,屬于可溶性劇毒金屬,因其無法降解且具有持久性,已成為廣為人知的環境污染物。鎘廣泛存在于我國稻田土壤中,極易通過土壤-作物系統積累在植物體內,且這種積累不可逆,進而通過植物源性食物。人體內攝入的鎘與蛋白質結合生成鎘-蛋白質,不僅影響人體磷(P)、鈣(Ca)等元素的吸收代謝,還會對人體免疫系統、骨骼系統、泌尿系統和腎臟等產生毒害作用,導致高血壓、蛋白尿、葡萄糖尿和骨骼軟化等病癥,并有可能致癌和致畸[1]。20世紀50年代震驚全世界的日本“骨痛病”事件即屬最嚴重的慢性鎘中毒事件[2],引起了全球的廣泛關注。
水稻(OryzasativaL.)在我國農業生產中占據著重要地位,每年我國水稻的消耗量占谷物消耗量的55%[3]。近年來中國稻米鎘污染現象日益嚴重,王茂起等[4]在2003—2004年期間對全國16個省市及自治區的29類食品進行檢測,結果發現送檢大米的鎘超標率達到5.33%;2007年,甄燕紅等[5]隨機抽檢全國部分市場的市售大米,結果13%的樣品鎘含量超標;謝燕湘等[6]于2012年在南方某市隨機采集市售大米進行鎘含量測定,結果鎘含量超標率高達46%;覃蕓等[7]在2016—2017年期間對柳州市市售大米樣品進行質量安全檢測,結果約9%的樣品鎘含量超標。且自2013年媒體先后揭露了湖南、廣東和江西等地出現的“鎘大米”事件,引起了公眾對鎘米的恐慌。因此,減少鎘毒素在稻米中的積累,特別是減少稻田受污染地區稻米鎘積累迫在眉睫。
目前控制水稻鎘污染主要有兩種途徑:一是通過化學或者生物學方法阻礙稻田土壤中的鎘通過根系進入植物體內;二是采用適當的農藝措施與耕作制度來減少鎘在水稻體內的積累。而通過遺傳育種技術培育低鎘水稻品種是最直接有效的方式,能從源頭有效保證水稻的安全生產。本文通過對影響水稻鎘吸收和運輸的環境、遺傳因素進行闡述,并結合目前國內外有關低鎘水稻選育的最新進展,探討選育低鎘水稻的可能途徑,旨在為無鎘或低鎘水稻篩選培育提供理論和實踐依據。
1 鎘與水稻的生長
鎘是最普遍的非營養重金屬元素,其不參與構成植物體的生物結構,也不參與植物體的代謝活動,在所有有害重金屬中,鎘的危害性最大。水稻極易吸收并積累鎘,但具基因型差異。一定濃度的鎘脅迫對水稻的生理生化和形態等方面有強烈的毒害作用,嚴重影響水稻的生長并導致其減產,稻米品質劣變。
1.1 鎘對水稻生理特性的影響
水稻體內積累過量鎘會導致嚴重的毒性效應,影響植株的光合色素含量、呼吸強度、蒸騰和光化學效率。鎘能夠強烈抑制原葉綠素酸酯還原酶的活性并阻礙氨基-γ-酮戊酸的合成,從而影響葉綠素的生物合成及與蛋白質的穩定結合,破壞光系統Ⅰ與光系統Ⅱ中的光合電子傳遞,阻礙光合磷酸化過程,使得ATP合成受阻,擾亂葉綠體進行正常的光合作用[8-9],且光系統Ⅱ比光系統Ⅰ更容易受到鎘的影響[10]。鎘對光合作用的影響還可通過形成Cd-葉綠素復合物來抑制和破壞葉綠素-蛋白復合物[9]。鎘脅迫濃度增大,水稻幼苗葉片的葉綠素含量和葉片光合特性指標(凈光合率、氣孔導度、蒸騰速率、胞間CO2濃度)均呈下降趨勢[11]。此外,鎘的積累還會抑制水稻植株細胞中蘋果酸脫氫酶、琥珀酸脫氫酶、葡萄糖-6-磷酸脫氫酶和6-磷酸葡萄糖酸脫氫酶等呼吸酶的活性,造成植株呼吸代謝紊亂[12]。
1.2 鎘對水稻產量的影響
水稻產量關系國民經濟和人們的生活,鎘脅迫也會影響水稻產量。但不同的研究結果不盡相同。黃冬芬等[13]發現土壤鎘處理對盆栽水稻結實率和千粒重影響不大;李玉清等[14]也發現鎘含量不同的水灌溉對水稻結實率和千粒重的影響并不明顯;丁園等[15]發現土培不同鎘濃度對水稻的千粒重、每穗實粒數和分蘗率影響不顯著,但在高鎘土壤(20.84 mg·kg-1)中生長的水稻,經濟產量降低10.65%。而陳娟[16]研究發現,在鎘處理下,水稻產量構成因素各指標均顯著低于對照,尤其是每穗粒數,表明鎘脅迫主要通過影響水稻幼穂分化進而影響其產量;葛才林[17]研究發現鎘處理對水稻穂數的抑制效果最明顯,但對千粒重的抑制程度較小;劉春梅等[18]研究認為鎘脅迫處理濃度上升,水稻每穗粒數和粒重均隨之減少,但千粒重無明顯變化。這些不一致的研究結果可能是與使用的水稻品種不同,且試驗過程中鎘處理的方式不一致有關。因此,鎘對水稻產量的影響需要繼續深入開展,以獲得普遍性的結果。
1.3 鎘對水稻稻米品質的影響
隨著人們消費結構升級,對稻米品質的要求越來越高,然而安全、健康營養的稻米已成為最基本的需求。水稻籽粒中積累鎘,會影響稻米的加工、營養、外觀和衛生品質。其中,稻米的衛生品質指籽粒中重金屬的含量根據國家糧食衛生標準(GB 2762-2005)[19]其鎘含量必需低于0.2 mg·kg-1。據報道,受鎘污染的稻米醇溶蛋白含量較多,而清蛋白和球蛋白等含量較少,賴氨酸、直鏈淀粉含量較低,營養品質較差[13,20]。受到高濃度鎘脅迫的水稻,其稻米的淀粉粘滯譜特征值與對照組相比發生了顯著變化,崩解值和最高黏度值明顯變小,而消減值、最終粘度與熱漿黏度明顯增大,表明鎘降低了稻米的食味品質[13]。李玉清等[14]發現在水稻生長發育期間,采用低于0.012 mg·L-1鎘的水灌溉對稻米的營養成分不會產生明顯影響,但會明顯影響稻米的外觀品質。丁園等[15]發現水稻生長在鎘污染(20.84 mg·kg-1)土壤中,其糙米中直鏈淀粉和鎘含量均明顯增加,而在低濃度鎘污染(4.48 mg·kg-1)的土壤中生長的水稻,其籽粒蛋白質含量顯著降低。鎘處理濃度越高,稻米中粗蛋白、粗淀粉和賴氨酸含量越低[17]。
2 水稻對鎘吸收、轉運和積累機制研究
水稻對鎘的吸收和積累的途徑主要有3種:1)根轉運體從根際土壤吸收鎘進入根細胞;2)木質部裝載介導鎘從根轉移到莖;3)鎘經韌皮部從莖轉運到籽粒。
2.1 水稻根對鎘的吸收
根系吸收鎘被認為是植物體內鎘積累最重要的一步,水稻根系最先接觸到稻田土壤中的鎘離子并通過質外體和共質體途徑吸收土壤溶液中的鎘。其中,質外體途徑主要利用鎘離子在細胞內外產生的電勢差以自由擴散的方式通過質外體空間進入到細胞內,運輸速度快,無需消耗能量,屬于被動運輸過程。而共質體途徑主要依靠細胞膜上的載體蛋白進行選擇性跨膜運輸,運輸速度相對較慢,屬于主動運輸過程,需要消耗能量才能將鎘轉移到根系細胞內,再通過胞間連絲實現根系細胞間的運輸[21]。
2.2 鎘在水稻地上部分中的轉運
水稻根吸收鎘后,由植物短距離運輸到木質部中完成加載,而后向地上部分運輸。水稻地上部分的鎘經木質部再通過篩管轉移到韌皮部,最后通過節點和穂輸送到籽粒中。從木質部到韌皮部的轉移是鎘到達水稻籽粒的必要條件。
研究表明,鎘離子經木質部從根系到莖部的轉移是決定水稻莖鞘和籽粒鎘積累的主要因素[22-23]。鎘在水稻植株中通過篩管進行轉運,稻米中91%~100%的鎘是由韌皮部轉運而來[24-25],水稻韌皮部汁液中的鎘濃度可能是決定其籽粒鎘含量的關鍵因素[26]。Fujimaki等[27]采用正電子發射示蹤成像系統對水稻灌漿期莖中鎘轉運和分布進行示蹤研究,表明鎘從莖基部的木質部運輸至韌皮部,水稻莖節是木質部中鎘向韌皮部轉移的重要組織并在鎘向籽粒轉運過程中起著關鍵作用。在水稻營養生長期,下部葉組織是主要的鎘儲存器官,越接近地面的葉片在水稻早期生長階段,鎘含量越高,但在完全成熟階段幾乎不含鎘[28]。
2.3 鎘吸收、轉運和積累相關基因
礦質元素需要依靠轉運子才能從根部轉移到莖[29],再從莖轉運至葉片、籽粒。近年來,許多調控水稻吸收、轉運和積累鎘的基因已被克隆(表1)。
OsIRT1和OsIRT2是水稻Fe2+轉運蛋白基因,由缺鐵誘導表達,在酵母中能轉運鎘,并且OsIRT1對鎘的特異性更大,可能在鎘的吸收過程中占主導地位[30]。過表達OsIRT1水稻對鎘更敏感且積累更多的鎘,表明OsIRT1確實具有轉運鎘的活性[31]。
水稻NRAMP蛋白(natural resistance-associated macrophage protein)與金屬離子轉運具有重要關系,水稻中共有7個Nramp基因,其中OsNramp1和OsNramp5被證實參與植株體內細胞對鎘的吸收和轉運。OsNramp5位于根外皮層和根內皮層的遠端,在根部高度表達,介導鎘從外部溶液進入到根細胞內[32]。Takahashi等[33]發現OsNramp1表達水平差異是導致Habataki(高鎘積累)和Sasanishiki(低鎘積累)兩種水稻鎘積累差異的主要因素;OsNramp1是水稻鎘積累的重要蛋白,在質膜上充當鎘流入轉運體,在鎘從根到莖的轉運過程中起著關鍵作用[34]。OsNramp6蛋白能夠在酵母中轉運鎘,其定位于胞質膜上,主要在水稻幼葉中表達,可能與葉片積累鎘有一定的關系[35]。
重金屬轉運ATP酶(heavy metal transporting ATPase,HMA)蛋白被認為是重金屬穿透細胞膜的外排轉運蛋白[36],水稻中有9個HMA基因。其中OsHMA3在根中高度表達,位于根部液泡膜上,能與流入胞質溶膠中的鎘螯合,以復合物的形式暫時儲存,降低胞質鎘遷移率,限制鎘從根部向地上部位轉移從而降低植株中鎘積累,過表達OsHMA3可以顯著降低籽粒鎘含量且不影響其他微量元素[37]。OsHMA2定位在質膜上,在水稻中充當鎘的內流轉運體,其主要在根部和節間表達,參與鎘的根-莖轉移,將鎘從薄壁組織重新裝載到維管束的韌皮部中并介導鎘在水稻植株中的分配[38-40]。Lee等[41]研究發現OsHMA9定位于質膜上,主要在維管束和花藥中表達,并證實OsHMA9是一種鎘外排轉運體。
水稻低親和陽離子轉運蛋白OsLCT1是水稻韌皮部中鎘轉運的主要轉運子,屬于外排型鎘轉運體,其定位于質膜上,在水稻營養生長階段的葉片和莖中高度表達[42]。OsLCT1RNAi干擾系籽粒鎘含量大約是對照組的50%,且籽粒中礦物質營養成分以及植株生長和產量不受影響[42]。李曜魁等[43]以華占和秈稻93-11為材料,從中克隆了OsLCT2基因,該基因編碼蛋白與小麥TaLCT1蛋白的同源性超過60%,由于小麥TaLCT1對鎘具有高親和力,介導鎘流向細胞質內,因此推測水稻OsLCT2可能也有類似的鎘轉運功能。
水稻OsLCD與其他已知的鎘轉運吸收基因均不同源,其位于細胞核與細胞質中,主要在根部和葉片中表達,OsLCD的T-DNA插入突變體lcd(OsLCD的第一個內含子中插入T-DNA)籽粒鎘積累量比野生型植株顯著降低,而鋅鐵含量、干重和種子產量無明顯變化,表明OsLCD是一個低鎘水稻育種的候選基因[44]。
金屬耐受性蛋白(metal tolerance proteins,MTPs)屬于陽離子擴散促進劑轉運蛋白家族,這類蛋白能夠調節植物體內重金屬的穩定狀態[45]。水稻中已鑒定出10個MTP基因,其中OsMTP1屬于Zn-CDF子群,主要分布在質膜上,能夠有效轉運鎘,并且可以使植物體內離子維持穩定狀態[46]。Yuan等[47]通過轉基因獲得OsMTP1 dsRNAi水稻植株,該轉基因株系幼苗對鎘的耐受性下降,而OsMTP1在野生型酵母中表達增強了酵母對鎘的耐受性。劉寶秀[48]對水稻OsMTP2進行分析,結果表明該基因為MTP家族的一個膜蛋白基因,主要在葉片、莖中表達,可能在水稻營養生長期對金屬離子轉運起重要作用。
ABC轉運蛋白(ATP-binding cassette protein)也參與植株金屬離子的轉運,目前水稻2個亞種基因組中已鑒定出121個該類蛋白質[49],大部分是由膜蛋白組成的,且能夠通過消耗ATP產生的能量把有毒重金屬排出細胞。其中OsPDR5和OsPDR9對鎘可能有轉運作用。Oda等[50]研究發現鎘脅迫下水稻OsPDR5在根中大量表達,是對照的3.5倍,但在莖中的積累不受影響,說明水稻OsPDR5可能參與鎘轉運并增強水稻對鎘的耐受。Moons[51]發現鎘能夠顯著誘導OsPDR9在水稻幼苗根系中的表達,表明OsPDR9參與水稻鎘脅迫響應。此外,CAL1蛋白能夠特異性結合細胞質中的鎘,驅使鎘從原生質體中轉移到細胞外側,隨后裝載到木質部汁液中,促進鎘從根部轉運至莖節當中,調控鎘在水稻中的積累[52]。
研究發現,長非編碼核糖核酸(lncRNAs)和微小核糖核酸(miRNAs)也能夠參與植物鎘脅迫應答反應,是植物體細胞中的關鍵調節因子。Liang等[53]通過深度測序,確定了鎘誘導水稻根中全基因組的lncRNA表達譜,結果表明,lncRNAs可能對鎘脅迫起重要作用。Fei等[54]用鎘處理水稻幼苗并對其根系進行深度RNA測序,序列分析結果表明,有9個lncRNA基因與鎘脅迫反應高度相關,對這9個基因進行鑒定,發現轉錄后基因調節也可能參與鎘脅迫反應。Ding等[55-56]鑒定出19個響應鎘脅迫的miRNAs,在鎘脅迫下有18個miRNAs表達下調,而miRNA528表達上調,后續研究發現鎘脅迫顯著誘導miRNA268的表達,且在鎘脅迫下水稻體內miRNA268的靶基因(NRAMP3)表達量顯著降低,而miRNA268過表達水稻植株幼苗鎘含量比野生型高,表明miRNA268是水稻耐鎘脅迫的負調節因子。研究發現,miRNA390的過表達水稻對鎘脅迫的敏感性增強,其鎘積累量比野生型高,且在鎘脅迫下水稻體內的OsHMA2和OsNRAMP5表達水平上升,說明miRNA390也是水稻耐鎘脅迫的負調節因子[57]。現有研究證實了miRNA166在水稻木質部的鎘負載中起作用,其過表達降低了鎘在水稻中的轉運,使得鎘從根向莖的轉移減少,并且過表達植株的稻米鎘含量顯著降低,水稻體內miRNA166對鎘的積累調節作用主要是通過調節其靶基因OsHB4實現的[58]。已鑒定的鎘響應lncRNAs和miRNAs對于進一步了解水稻體內鎘代謝的分子機制具有重要意義,目前對lncRNAs和miRNAs的研究還不夠深入,大部分的lncRNAs和miRNAs生物學功能尚不清楚,未來需要加大研究力度,以便為系統了解水稻鎘代謝的機制提供有價值的參考意見。
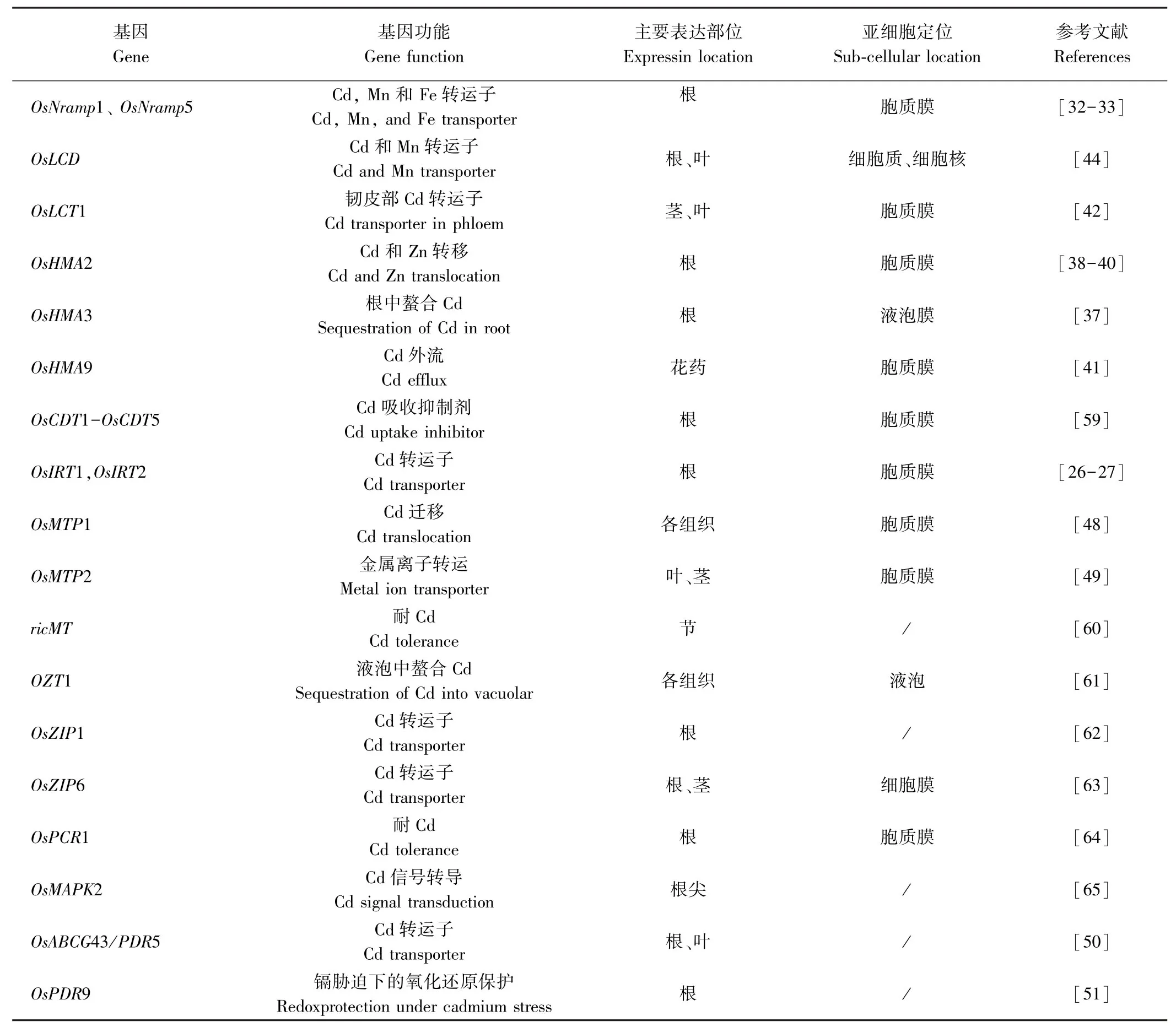
表1 水稻中已克隆的負責鎘吸收與轉運相關基因Table1 Cloned genes responsible for cadmium uptake and translocation in rice
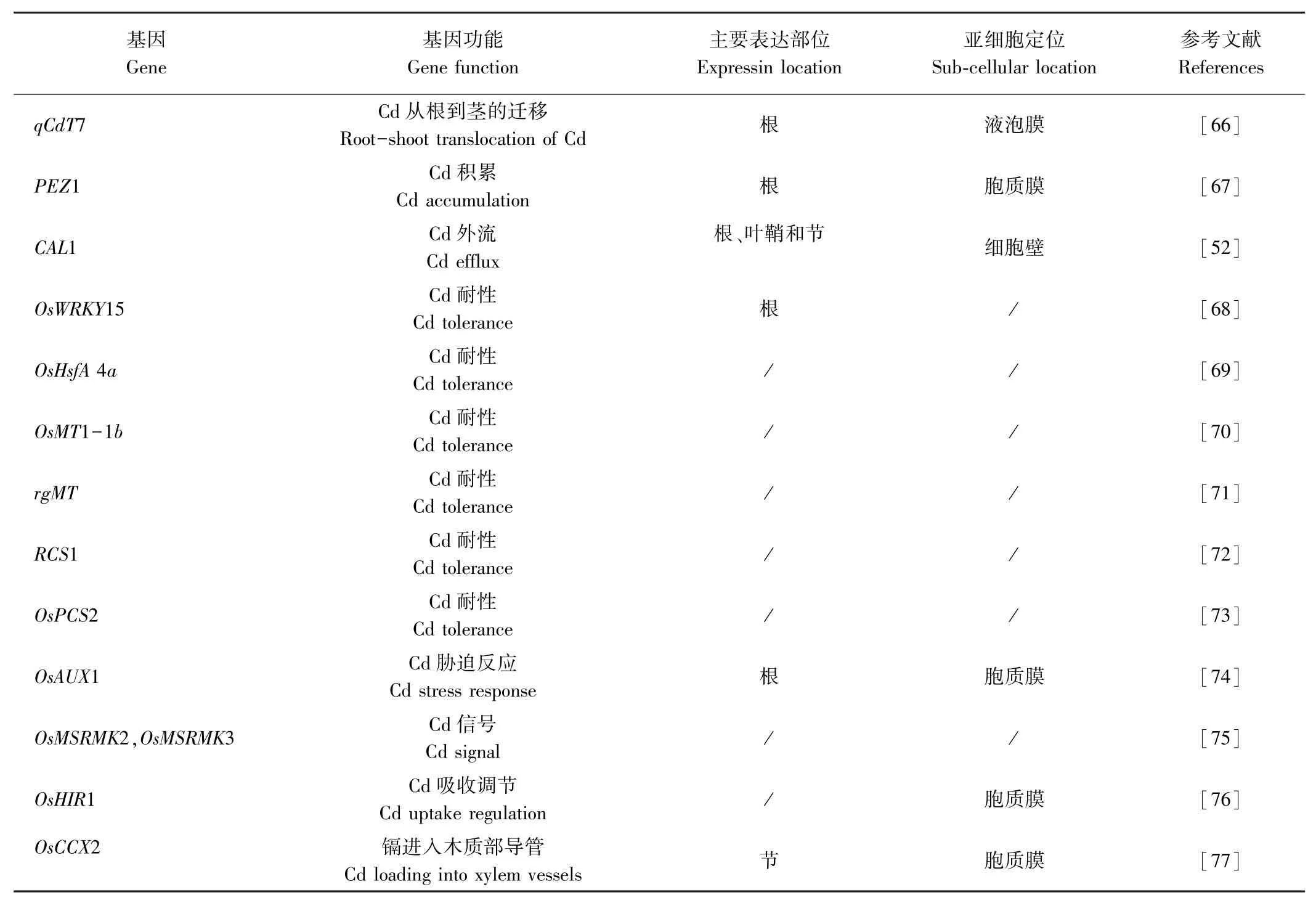
表1(續)
2.4 水稻鎘吸收、轉運和積累的影響因素
2.4.1 水稻自身遺傳特性 不同水稻品種遺傳背景不同,其生理特性和形態結構也不同,從而導致水稻品種間對鎘的吸收和積累也不盡相同。仲維功等[78]選用43個水稻品種進行田間試驗,發現粳稻根部鎘積累能力最強,秈稻精米和籽粒中鎘積累量最高,其次是雜交稻和粳稻;Ye等[3]選用9個水稻品種進行試驗,得到了與仲維功等[78]相同的試驗結果;鄧偉等[79]發現高鎘積累品種對鎘的吸收特性表現為積累速度快或者時間長,而低鎘積累品種則表現為積累速度慢或時間短;Zhong等[80]研究發現粳稻比秈稻更容易積累鎘,但是鎘使得秈稻產量明顯下降,而粳稻產量均增加。
2.4.2 環境因素 土壤環境是影響水稻鎘含量最主要的因素。水稻對鎘的吸收與鎘的總含量相關不大,而與鎘的生物有效性直接相關。稻田土壤的物質組成(質地、有機質)、化學性質(pH值、氧化還原電位、陽離子交換量)及微生物活動都會影響土壤中鎘的生物有效性[81]。黃德乾等[82]采用溫室盆栽試驗研究了3種不同類型土壤上水稻對鎘污染的響應,發現不同類型土壤上水稻中鎘的濃度大小為紅壤>紅壤性水稻土>烏柵土。
水稻對鎘的吸收取決于土壤的物理和化學條件,其中土壤酸堿度(pH值)和土壤有機物質含量是決定鎘吸收量的最主要因素[83]。土壤pH值在水稻根系對鎘的吸附以及鎘在土壤溶液中的形態、溶解度和遷移率方面起著最重要的作用[84]。研究表明稻田土壤酸堿度與水稻鎘吸收量密切相關,在一定范圍內,兩者呈負相關[85]。因此,防止稻田土壤酸化可有效降低生物可利用鎘的釋放。在受鎘污染稻田施加CaCO或Ca(H2PO4)2等堿性物質可提高稻田土壤的pH值,從而減弱水稻鎘毒害作用。土壤有機物質能夠與鎘絡合或螯合,因此,可以通過摻入有機物質,如施用有機肥(農家肥、堆肥、植物秸稈等),增加鎘與有機物質的結合,進一步減少稻田中的可吸收鎘。
此外,土壤氧化還原電位的變化也會引起鎘的溶解度發生改變。研究表明,在淹水期間,稻田土壤氧化還原電位值降低,鎘被吸附或形成硫化鎘(CdS)沉淀,溶解度降低,因此鎘不容易被吸收。然而,排水期間的稻田土壤需要氧氣,其Eh值升高,鎘將形成水溶性CdSO4,釋放出可被根系吸收的鎘離子[86]。加強稻田淹水管理也能夠有效降低稻米鎘積累。
土壤微生物在活動過程中能夠分泌出有機物質、蛋白質和多聚糖等,這些物質不僅能夠改變根際土壤的pH值,還能夠吸附鎘,對鎘具有氧化還原、絡合或沉淀等作用,從而影響根系對鎘的吸收。研究表明,水稻根際微生物群落與根系分泌活動密切相關,兩者相互促進[87]。根系分泌物包括有機配體(蘋果酸、腐殖酸、檸檬酸、黏膠、糖類、酚類和氨基酸等)和無機配體(C1-、SO42-、NH4+和PO42-),這些分泌物進入到稻田土壤后,不僅作為微生物活動的能量來源,而且有些能夠與鎘螯合,使得根際周圍土壤的pH和氧化還原電位值發生變化,改變稻田土壤中可利用鎘離子的含量[88]。
3 低鎘水稻育種
稻米鎘積累會導致嚴重的食品安全問題,對人類健康存在極大隱患。為了防止人體攝入鎘,培育和選擇低鎘水稻品種對于人類健康有深遠意義。
3.1 常規雜交和誘變育種
雜交育種是選育低鎘水稻的技術之一。可以根據水稻籽粒鎘積累的規律,選育低鎘不育系,低鎘恢復系和配組獲得低鎘雜交水稻[89]。
誘變育種具有突變頻率高、穩定時間短、操作方法簡便、可打破性狀連鎖并實現基因重組等優點,是培育新品種,實現種質資源創新的一項重要技術。高能重離子束具有誘導突變頻率高且輻射劑量低(經輻照后的植物能夠正常生長)的優勢,目前已經應用于輻照植物體以獲得非轉基因突變體。林園園等[90]利用重離子束輻照技術誘變水稻,發現重離子束能夠誘發水稻對鎘吸收能力的突變。由于誘變具有隨機均勻特性,因此通過誘變技術可以在突變體中篩選出符合要求的低鎘水稻新品種。Ishikawa等[91]也采用該輻照技術,篩選得到3個突變體,且這3個突變體的稻米鎘積累濃度均低于0.05 mg·kg-1,屬于低鎘新品種。
將雜交育種與誘變育種方法結合運用,在雜交重組的基礎上對尚未穩定的水稻雜交后代材料進行適當劑量的誘變處理,可以增加突變頻率、擴大突變譜、打斷基因連鎖,從而促進基因重組過程產生更多突變類型。據統計,1970—1983年期間,國外以誘變突變體作為雜交親本培育出的新品種占誘變育種新品種的60.5%[92]。龐伯良等[93]選用水稻作為試驗材料,并將輻射誘變與常規雜交技術相結合進行試驗,結果表明,復合處理產生的突變類型與頻率均比各種單處理要高,且能夠產生更好的育種效果。因此,誘變與雜交相結合也可以在一定程度上為選育低鎘水稻品種創造豐富的育種材料,提高低鎘水稻育種效果。
3.2 基因工程育種
參與水稻鎘代謝的基因已鑒定30多個,其中有些是調控鎘積累的關鍵基因,利用轉基因手段對這些關鍵基因進行遺傳操作可獲得低鎘水稻材料。例如,過表達OsHMA2轉基因水稻,其籽粒中鎘含量僅為野生型的一半[39],利用RNAi干擾技術,抑制OsLCT1的表達,獲得的轉基因植株籽粒鎘含量大約是對照組的50%[42]。Shimo等[44]將T-DNA插入到OsLCD中,通過篩選,鑒定出了一個新的耐鎘突變體lcd。
此外,可利用CRISPR技術在水稻中定點突變吸收鎘基因或者導入負責排鎘的基因,阻斷鎘進入到植物體內從而獲得排鎘水稻[94]。已有研究以雜交水稻的骨干親本作為受體材料利用CRISPR創制出低鎘不育系、恢復系,并采用雜交技術,配制出了高產、優質且低鎘的雜交稻新品種,使得低鎘雜交稻選育取得了突破性的進展[95]。
3.3 分子標記輔助育種
探索水稻鎘積累特性費時費力,為此提出了建立分子標記輔助育種系統,該系統可以利用追蹤水稻遺傳組成的分子標記從突變體中培育水稻新品種,縮短育種時間,達到精準育種,提高選育效率并節約篩選成本。陳彩艷等[96]開發了4個低鎘分子標記,并運用分子標記輔助選擇技術對5 769份水稻新品進行篩選,最終獲得攜帶1~3個低鎘分子標記的80份新品系,經過田間試驗證明了其中有些品種雜交后具備穩定的低鎘遺傳特性。Ishikawa等[91]研究確定降低鎘吸收的基因為OsNRAMP5,為此構建了OsNRAMP5基因的分子標記輔助選擇體系,在突變體后代群體中篩選攜帶OsNRAMP5的品種。由于離子束輻射產生的突變體是非轉基因植物,更容易被消費者接受。
4 結論與展望
在過去幾年內,國內外加大了對水稻鎘代謝分子機制方面的研究,同時也取得了重大進展,發現了一些新的水稻鎘轉運體家族,然而由于鎘與其他微量元素之間存在協同/拮抗等作用,水稻對鎘的吸收、轉運、積累機制復雜,解決稻米鎘污染問題仍任重道遠。未來低鎘水稻的研究可以圍繞以下幾個方面深入開展:1)深入挖掘與水稻鎘代謝相關的金屬轉運體,實施多基因聚合育種;2)開發與水稻低鎘基因緊密連鎖的分子標記,推動分子標記在育種中的應用并將分子育種與常規育種有機結合;3)加強各農藝措施的綜合實施,降低外源鎘的生物有效性,建立安全、高效的栽培技術體系,最終培育出穩定遺傳,安全優質的低鎘水稻新品種。
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
