耐高溫球等鞭金藻(Isochrysis galbana)藻種的選育與餌料特性初步分析*
2021-02-03 01:44:44趙東會趙麗達王寅初焦緒棟
海洋與湖沼 2021年1期
關鍵詞:生長
趙東會 趙麗達 尤 宏 秦 松, 王寅初, ① 焦緒棟, ①
(1. 哈爾濱工業大學(威海)海洋科學與技術學院 威海 264209; 2. 中國科學院煙臺海岸帶研究所 煙臺 264003;3. 中國科學院海洋大科學研究中心 青島 266071; 4. 南陽師范學院生命科學與技術學院 南陽 473061)
餌料微藻可以在生物鏈中作為魚蝦幼苗良好的開口食物, 是水產動物苗種培育階段的關鍵因素之一。與傳統的人工飼料相比, 微藻生物餌料有易被養殖對象消化、便于培養、營養全面、對水體污染小等獨特的優勢(許家輝等, 2018)。目前研究的餌料微藻主要包括三角褐指藻(Phaeodactylum tricornutum)、等鞭金藻(Isochrysis sp.)、亞心形扁藻(Platymonas subcordiformis)、中肋骨條藻(Skeletonema costatum)、威氏海鏈藻(Thalassiosira weissflogii)和海水小球藻(Chlorella sp.)等(劉皓等, 2010; 熊良偉等, 2013; 劉祚屹等, 2020)。
然而, 在夏季高溫條件下, 水溫影響了餌料生物的正常繁殖與生長(包永波等, 2004)。普通餌料微藻容易衰敗, 造成餌料微藻供應不及時或無法足量供應(鄒琰等, 2010), 為水產苗種的生產帶來諸多隱患。三角褐指藻在 25°C 下細胞生長最好, 小球藻(Chlorella vulgaris)在 5—30°C 均可正常生長, 20°C 為最適宜的生長溫度(趙婷等, 2016)。亞心形扁藻在28°C 的生長量最大, 31°C 的溫度條件下, 亞心形扁藻的生長量下降。不同溫度條件下, 等鞭金藻塔溪堤品系Isochrysis galbana (Tahitian strain)的相對生長率在25°C 條件下最高, 其最適生長溫度為 20—30°C (梁英等, 2011)。諸多研究表明, 目前的餌料微藻最適耐受溫度普遍低于30°C。因此尋找可以在30°C 及以上溫度條件下穩定生長的餌料微藻, 以保障夏季餌料微藻的供應量是比較迫切的。
單環刺螠(Urechis unicinctus)屬螠蟲動物門(Echiurioidea) 、 螠 綱 (Echiurida) 、 無 管 螠 目(Xenopneusta)、刺螠科(Urchidae)、刺螠 屬(李諾等,1998)。目前, 單環刺螠的研究主要集中在單環刺螠的繁育、池塘養殖方面的創新工作(王淑芬等, 2016; 許星鴻等, 2016), 但對單環刺螠的攝食生理學研究甚少。單環刺屬于濾食性動物, 主要以金藻、扁藻、小球藻等微藻作為餌料, 其中以金藻作為開口餌料效果較好(許星鴻等, 2016)。其夏季育苗一般自 5 月下旬開始, 至 8—9 月份結束, 育苗過程需經過夏季高溫期。目前所用的餌料微藻難以在高溫高光照條件下存活, 篩選耐高溫餌料微藻可以滿足夏季單環刺螠幼苗對活體餌料微藻的需求。
本文從夏季水溫超過 40°C、已發生衰敗的萊州某育苗場的球等鞭金藻培育池中分離出 4 株球等鞭金藻, 分別命名為PHY7001—PHY7004。分析了各藻株在33°C、40°C 的水溫下的生長狀態和藻株細胞大小、粗蛋白含量, 并對生長速率最快的一株耐高溫球等鞭金藻(PHY7004)的培養條件進行優化。通過測試單環刺螠幼苗對耐高溫藻株的攝食能力, 選育出具有較好適口性的藻株, 為后期的規模化利用提供了理論和現實依據。
1 材料與方法
1.1 微藻分離與培養
本實驗藻液樣品是在夏季從萊州市順昌水產有限公司的餌料培育池取回, 實時水溫為 43°C, 其中培育的藻類經過長期的高溫已發生嚴重衰敗。取回后立刻通過顯微操作技術分離耐受住高溫脅迫的微藻活細胞, 對分離的單細胞進行培養后得到藻株, 分別命名為: PHY7001、PHY7002、PHY7003、PHY7004。
微藻培養采用f/2 培養基(Guillard et al, 1962), 溫度為25°C, 鹽度為32, 光暗周期為 12 h/12 h, 光照強度 12 μmol/(cm2·s)。每天定時搖勻藻液 1 次, 以防細胞貼壁生長對實驗結果產生干擾。
1.2 藻株的耐高溫測試
本實驗設 A(33°C)、B(40°C)兩組溫度, 對分離藻株的耐高溫性能進行測試。每組每藻株設三個平行。將分離培養后達到對數生長期的藻株分別取100 mL藻液至250 mL 三角瓶。將A、B 組分別置于溫度為33°C、40°C 的恒溫光照培養箱中培養, 光暗周期為24 h/0 h, A、B 兩組有效光強均為 16 μmol/(cm2·s)。采用681 nm 吸光光度值表征其細胞數目。
1.3 藻株PHY7004 培養基的優化
PHY7004 是33°C 條件下生長速度最快的藻株。本實驗對 PHY7004 進行培養基的優化, 優化的指標為 pH 值和氮磷比(NaNO3∶Na2HPO4), 其中 NaNO3的濃度為 75 g/L; Na2HPO4·2H2O 的濃度為 5.65 g/L。pH 值設置三個梯度: pH=7, pH=8, pH=9, N/P 比設置三個梯度: 12∶1, 13∶1, 14∶1, 對 pH 值和 N/P 進行交叉實驗, 共9 個梯度。
將PHY7004 置于33°C 的恒溫振蕩培養箱中培養,光暗周期為 24 h/0 h, 光強為 12 μmol/(cm2·s)。
1.4 藻株鮮重總蛋白含量的測定
用BCA 總蛋白濃度檢測試劑盒(上海碧云天生物技術有限公司)對所篩選出的耐高溫餌料微藻進行鮮重下蛋白質含量的測定。用Origin 軟件線性擬合得到蛋白質的標準曲線: A=13.50C+0.1170, R2=0.9937。其中: A 為 562 nm 的吸光度值; C 為總蛋白質濃度(mg/mL)。
1.5 單環刺螠幼苗飼喂實驗
選取 4 月齡, 體重為 0.32—0.90 g, 體長為2.1—3.9 cm 的單環刺螠幼苗200 尾, 從中挑選出生長狀態良好的150 尾幼苗, 隨機分為5 組, 1 組30 尾單環刺幼苗, 每組設 3 個平行重復。分別置于 5 0 L養殖容器中, 底部鋪設15 cm 干凈海沙; 采用過濾海水每天6:00am 換水1 次, 充氣暫養7—10 d。分別用PHY7001、PHY7002、PHY7003、PHY7004 的新鮮培養液, 于每天 5:30pm 定時飼喂單環刺螠幼苗, 投喂的餌料微藻的初始濃度為2×106—3.5×106CFU/mL。不放單環刺螠幼苗為對照組。飼喂實驗持續 5 d, 于實驗開始后第3 天開始取樣。飼喂后2 h 在養殖池的不同位置分別取3 次海水樣品, 測試海水濁度。根據公式計算單環刺螠幼苗對所篩選出耐高溫餌料微藻的攝食強度(Fi)(孫濤等, 2017)。
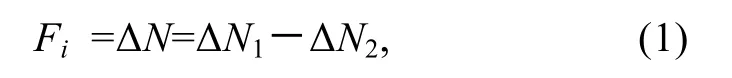
式中, ΔN1表示試驗組投餌1 h 后因單環刺螠攝食和沉降造成濁度的改變值(NTU); ΔN2表示對照組投餌1 h后因沉降造成濁度的改變值(NTU)。
2 結果與討論
2.1 藻株的分離
通過顯微操作技術分離出4 株球等鞭金藻。分別命名為: PHY7001、PHY7002、PHY7003、PHY7004。單細胞培養48 h 后, 觀察各藻株顏色為綠色, 顏色純正, 鏡檢細胞大小一致, 無原生動物污染。
利用倒置顯微鏡拍攝各藻株的細胞照片后, 用Digimizer 軟件測量細胞直徑(圖1)。計算各藻株細胞的平均直徑為 5.5—7.5 μm, 與常規的球等鞭金藻(5—7 μm)相比, 大小差別不明顯。
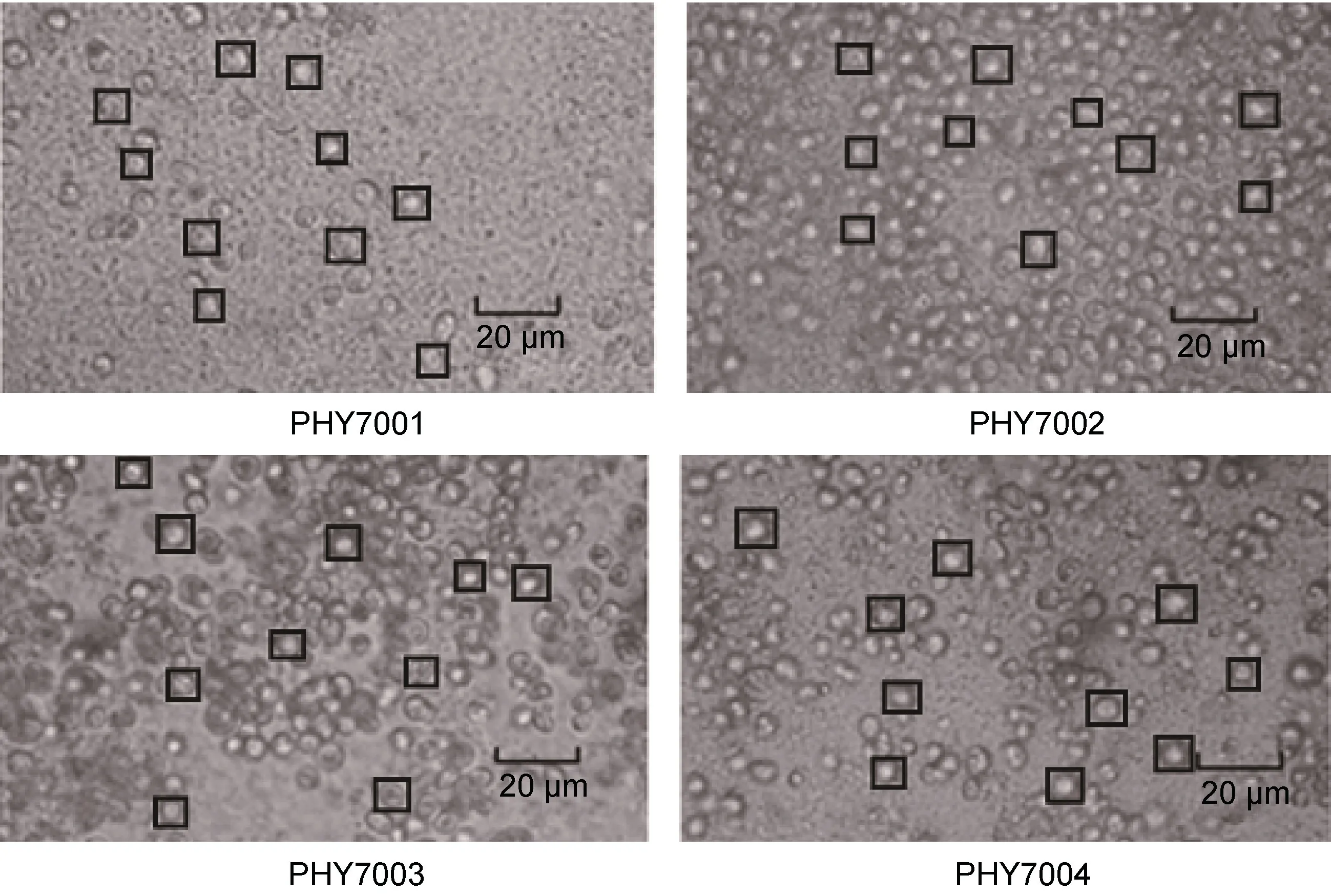
圖1 分離的4 株耐高溫球等鞭金藻Fig.1 Four isolated strains of I. galbana resistant to high temperature
在培養微藻過程中, 與控制溫度相比, 光照條件易控制且控制成本較低, 因此本實驗目的為篩選耐高溫藻種, 對光照強度的設置條件為不抑制球等鞭金藻正常生長即可。本實驗微藻培養與耐高溫測試實驗的光照條件分別為 12 和 16 μmol/(cm2·s), 均在球等鞭金藻可正常生長的光照強度范圍內(郭兵等, 2011)。
2.2 耐高溫性能測試
將分離的藻株在 33°C 水溫中進行培養, 發現分離的藻株均可在33°C 水溫下存活。其中從接種開始到第7 天, 各藻株的細胞數目均有增加; 與第7 天相比, 第 9 天的 PHY7001、CHY7004 的細胞數量繼續增加, PHY7002、CHY7003 的細胞數量沒有大的變化;根據生長曲線的走勢, PHY7001 和PHY7004 的細胞數目可能繼續增長。在培養周期內, PHY7004 的細胞數目幾乎呈線性增長, 生長狀態最優, 推測該藻株對高水溫的適應能力較好。其他藻株也展示出了較好的生長能力, 存在不同的生長滯后過程, 提示其對高水溫的適應能力需要進一步馴化(圖2)。
將分離的藻株在 40°C 水溫中進行培養時, 藻株的生長狀態與在 33°C 時完全不同。由圖 3 所示,PHY7001、PHY7002、PHY7003、PHY7004 均不能耐受40°C。與第1 天相比, 第3 天的4 株球等鞭金藻的細胞數量大幅減少, 第3 天的細胞數目約為第1 天的1/3—1/6 倍。在培育的第3—9 天, 雖然細胞數目中間有小幅度的上升, 但是最終的細胞數目仍普遍呈下降趨勢, 這可能是由于球等鞭金藻最終無法適應高溫造成的。

圖2 球等鞭金藻的存活及生長性能(33°C)Fig.2 Analysis of the survival and growth capability of I.galbana (33°C)
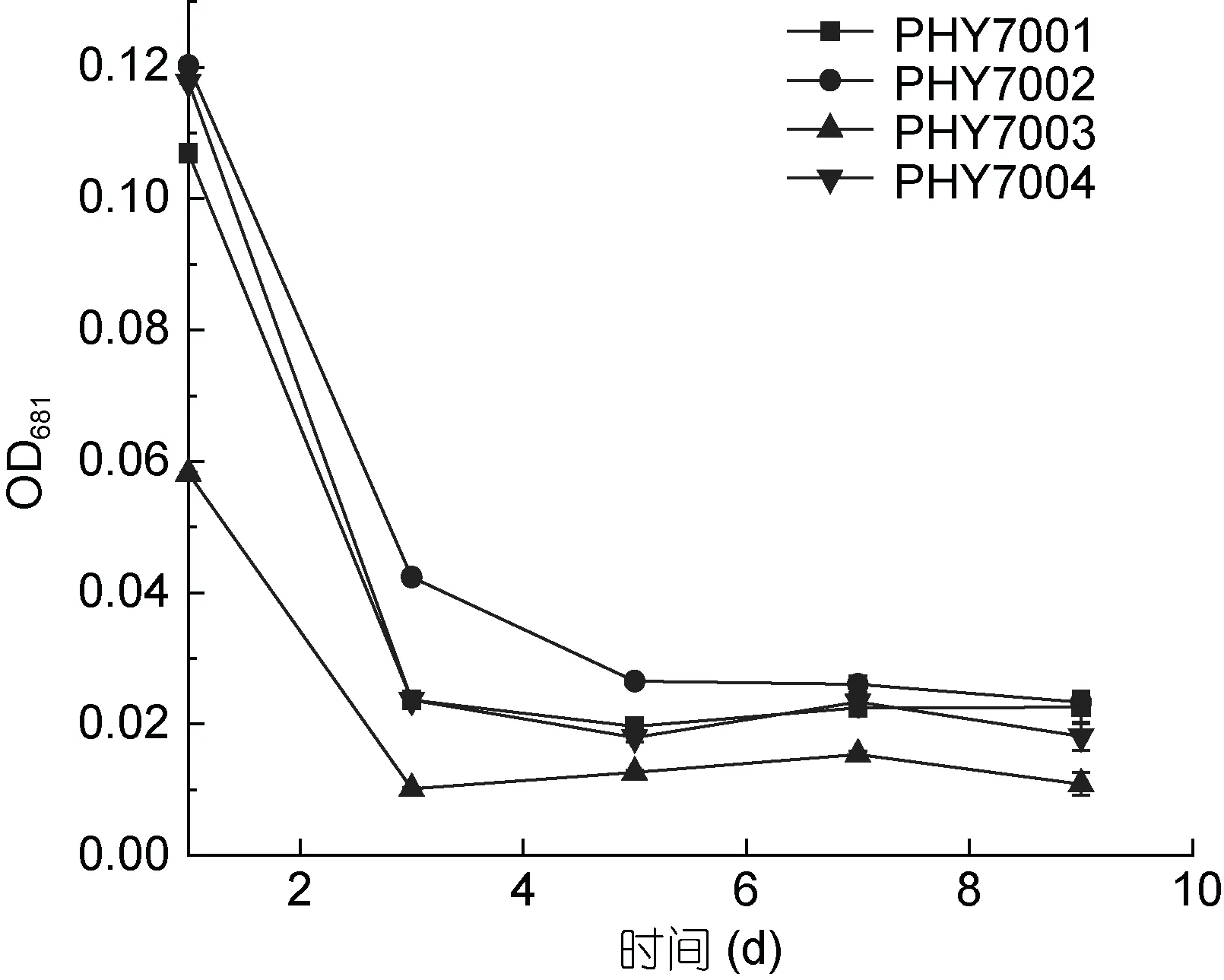
圖3 球等鞭金藻的存活及生長性能(40°C)Fig.3 Survival and growth of I. galbana (40°C)
2.3 球等鞭金藻的優化
對可耐受33°C 且生長速率最快的PHY7004 繪制其在33°C 下的生長曲線。用Origin 軟件對數據進行線性擬合, 擬合出的方程為: A=0.1097-0.0015T+0.0048T2-0.0003T3, R2=0.9920。由圖 4 可見, 當PHY7004 起始接種量 OD681值為 0.1007 時, 第 1—2天為 PHY7004 的適應期; 第 2—9 天為 PHY7004 的對數生長期; 第 9—11 天為 PHY7004 的平穩期; 第11—12 天為 PHY7004 的衰退期。培育第 9 天時PHY7004 的OD681值為0.2491, 細胞濃度達到起始濃度的1.37 倍。梁英等(2011)對球等鞭金藻進行生長曲線的繪制, 第 10 天生長達到平穩期, 與本研究結果基本一致。叢超等(2017)研究的5 株球等鞭金藻均在培養第 4—5 天后便進入穩定期, 與本研究中的PHY7004 相比, 其生長周期要短得多。這可能與培養條件和株系不同有關。
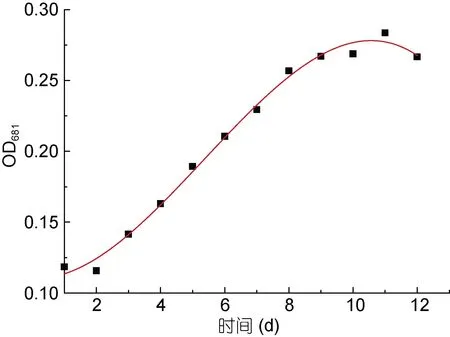
圖4 球等鞭金藻PHY7004 的生長曲線(33°C)Fig.4 Growth curve of I. galbana PHY7004 (33°C)
目前微藻培養的生長速度普遍較快, 微藻的生長速度與接種濃度、培養方法和培養條件等有很大關系(Moheimani et al, 2011; 孫春曉等, 2014)。本實驗數據僅限于實驗室條件下測出, 藻株應用于大規模生產時的繁殖速度還有待進一步研究。
從圖5 和圖6 可以看出, pH 值范圍為7—9, 氮、磷比例分別為12:1、13:1、14:1 時, PHY7004 的細胞數量和生長速率均呈增長趨勢, 這可能是因為 pH 值和氮、磷比例均在球等鞭金藻耐受范圍內。球等鞭金藻在pH 值為3—9 的條件下均能生長, 研究發現金藻生長的最適pH 范圍為7.0—9.0, 當偏離這個范圍時,藻體生長和體內代謝活動都將會受到抑制(王珺等,2008)。在氮、磷比等于 16:1 的條件下, 球等鞭金藻的生長速度最快(劉東艷等, 2002)。
在培育第2 天時, 各條件下的藻生長速率均增長最快; 與第2 天相比, 培育第3 天各條件下的藻生長速率明顯降低; 培育第 3—8 天藻生長速率均穩步上升。據圖 5 可知實驗期為球等鞭金藻的對數生長期,后期還會增長。
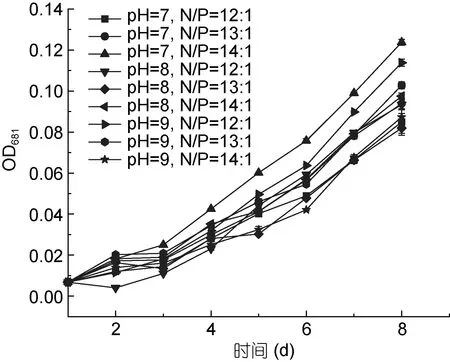
圖5 球等鞭金藻PHY7004 在不同優化條件的生長曲線(40°C)Fig.5 The growth curve of I. galbana PHY7004 in different optimum conditions (40°C)
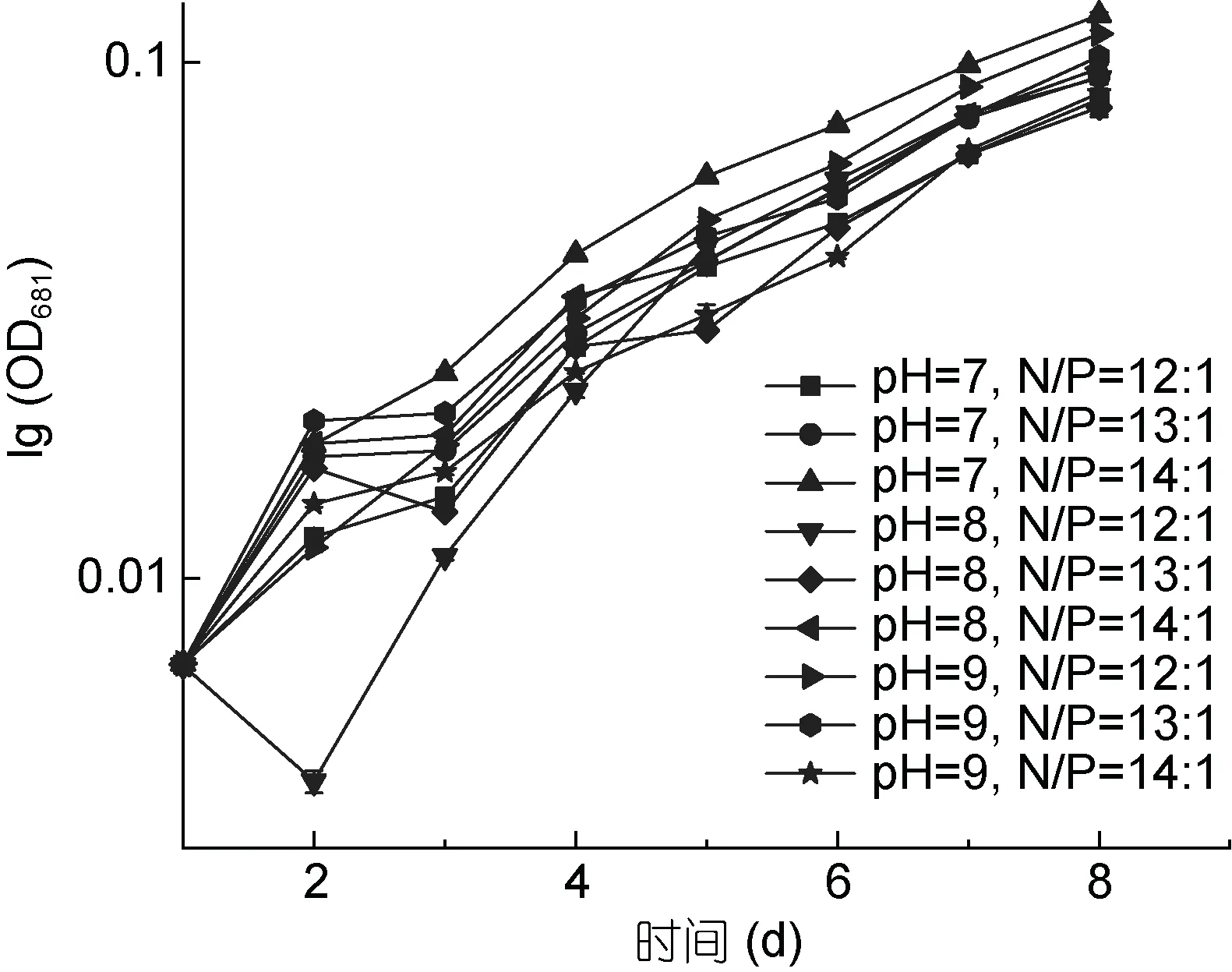
圖6 PHY7004 在不同優化條件下的生長速率(40°C)Fig.6 The growth rate of PHY7004 in different optimum conditions (40°C)
通常海水中的 pH 比較穩定(pH≈8.2), 但受自然因素影響, 海水pH 值會產生波動, pH 會影響脂質生產率, 對細胞代謝具有廣泛影響(Ren et al, 2013)。氮磷營養鹽在藻類生長過程中是重要的影響因子。磷對于生物質生產至關重要。因此本實驗對培養基中的氮磷比和 pH 值進行優化, 結果表明, 在 N/P=14:1,pH=7 的狀態下, 藻細胞的生長速率最快, 細胞數量最高, 因此, PHY7004 在 33°C 下最適的培養基N/P=14:1, pH=7。在 33°C 下, 當培養基 N/P=12:1 時PHY7004 較適pH=9; 當培養基N/P=13:1 時PHY7004較適 pH=9; 當培養基 N/P=14:1 時 PHY7004 較適pH=7; 當培養基 pH=7 時 PHY7004 較適 N/P=14:1; 當培養基 pH=8 時 PHY7004 較適 N/P=12:1; 當培養基pH=9 時 PHY7004 較適 N/P=12:1。
2.4 4 株球等鞭金藻蛋白質的含量
球等鞭金藻平均蛋白質含量為 34.29 mg/L, 分離的4 株球等鞭金藻蛋白質含量差異較大, PHY7003蛋白質含量最高, 見表1。飼料蛋白質含量對動物消化酶影響顯著, 在魚蝦等水產動物中均有報道(黃凱等, 2003; 趙東海, 2004)。宋萍萍等(2015)研究的球等鞭金藻的蛋白質含量可達到 110 mg/L, 其蛋白含量遠高于本實驗藻株。蛋白質的含量與微藻的品系、培養條件和培養方式有關(Wang et al, 2004; 張躍群等, 2009)。
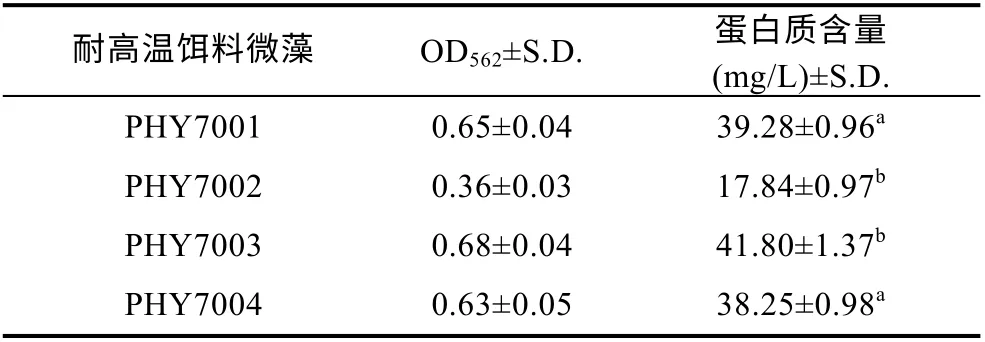
表1 耐高溫餌料微藻蛋白質含量Tab.1 Protein content of high-temperature-resistant microalgae bait
總蛋白含量是評價微藻營養價值的重要指標之一, 但不是唯一標準。對于餌料微藻而言, 脂肪含量,特別是所含不飽和脂肪酸的總量和組成也是評價微藻是否適合海水養殖生物應用的重要指標。微藻飼養動物的效果不因微藻蛋白質水平而異, 微藻所含的必需脂肪酸能否滿足動物的營養需求才是評定微藻營養價值的重要指標(Brown et al, 1997)。高蛋白飼料或餌料對養殖動物體內的消化道、酶系、微生態平衡都會有顯著影響, 有時也會損害養殖動物的健康。
藻類蛋白的研發正在迅速發展, 小球藻、柵藻、螺旋藻等蛋白產品開發已吸引了眾多的目光, 在保健品和飼料行業小有市場, 然而其他72000 種微藻資源還未得到研究和開發。本文研究的球等鞭金藻藻株,為微藻蛋白的開發提供了一個新的原料。球等鞭金藻蛋白質含量在已經研究的微藻中含量較高。以這幾株藻為基礎進行微藻工業開發, 在生物能源、微藻餌料、單細胞蛋白等方面均有巨大的應用潛力, 值得對其進行更多的研究和應用發掘。
2.5 單環刺螠的攝食強度
在篩選出的耐33°C 的球等鞭金藻中, 4 月齡單環刺螠幼苗(體重0.32—0.90 g, 體長2.10—3.90 cm)對篩選出的四株耐高溫球等鞭金藻的攝食強度無顯著性差異(P>0.05), 單環刺螠幼苗對PHY7002 的攝食強度最大, 見表 2 。一般認為餌料顆粒的大小是判斷其是否適合育苗生產的關鍵要素。由表 2 可知,PHY7002 的細胞大小跟濾食性動物 4 月齡單環刺幼苗的口徑相一致。值得注意的是, 本實驗中的單環刺螠幼苗對蛋白含量最少的PHY7002 攝食強度最大,對蛋白含量最多的 P HY7003 攝食強度最小。這說明藻株的蛋白含量與單環刺螠對其攝食強度不一定成正比, 這與前人研究結果一致, 飼料蛋白含量與增重率并不呈正比(朱偉等, 2005)。
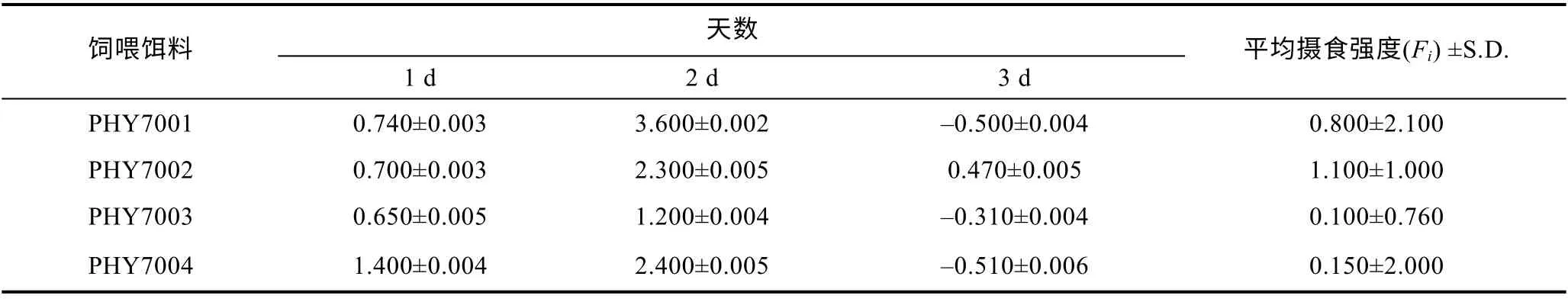
表2 單環刺螠幼苗對球等鞭金藻的平均攝食強度Tab.2 Feeding intensity of larval U. unicinctus on Days 1—3
最新研究表明, 在長期的進化過程中, 許多海洋生物幼體擁有選擇適合自身生長所需的藻類的“特異功能”, 可利用其尚未明確的“嗅覺”等方式選擇自己喜歡的餌料(麥康森, 2020)。球等鞭金藻(I. galbana Parke)是一種金黃色的金藻, 個體小、繁殖速度快、營養豐富, 并且沒有細胞壁, 容易被動物幼蟲消化吸收(王珺等, 2002), 可作為優質餌料。已有研究(Kain et al, 1958; 陳椒芬等, 1985; 周汝倫等, 1994)報道,球等鞭金藻(I. galbana Parke)是貽貝、牡蠣、泥蚶等雙殼類貝苗和對蝦幼體的優良餌料。但目前還沒有用球等鞭金藻飼喂單環刺螠的報道。單環刺螠人工養殖正處于摸索起步階段, 目前用餌料微藻飼養中國對蝦, 牙鲆等水產動物較多, 對于單環刺螠攝食餌料微藻的研究甚少。有研究表明, 使用微綠擬球藻和三角褐指藻的混合餌料投喂單環刺螠幼苗會取得比較明顯的促生長效果(常林瑞等, 2018)。因此, 在今后單環刺螠人工配合飼料的研發中可適當添加球等鞭金藻。
3 結論
通過顯微操作技術分離出4 株球等鞭金藻。經耐高溫測試, 分離出的 4 株球等鞭金藻均可耐受 33°C,且 PHY7004 生長速率最快, 分離出的各藻株細胞的平均直徑為 5.5—7.5 μm; 經培養基優化實驗得, 可耐受 33°C 且生長速率最快的一株球等鞭金藻(PHY7004)在 33°C 下較適的培養基條件為 N/P=14:1,pH=7。通過耐高溫藻的單環刺螠幼苗飼喂實驗, 發現單環刺螠幼苗對PHY7002 攝食強度較高, 這兩株藻后期可以作為單環刺螠育苗期間的餌料微藻備選。人們對單環刺螠的攝食生理學還未開展系統的研究, 本實驗可為單環刺螠的池塘養殖技術提供一定的參考。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
