p300/p53/Smad3通路參與人心房成纖維細胞衰老相關心房纖維化*
2021-02-05 01:02:24賴穎瑜高小燕周慧珊王釗煜彭德威鄧春玉
中國病理生理雜志 2021年1期
關鍵詞:水平
賴穎瑜 , 高小燕 , 周慧珊 , 李 昕 , 王釗煜 ,彭德威 , 饒 芳 △, 鄧春玉 ,3△
(1南方醫科大學藥學院,廣東廣州510515;2廣東省心血管病研究所心內科,廣東省人民醫院醫學研究部,廣東省醫學科學院,廣東廣州510080;3華南理工大學醫學院,廣東廣州510006)
衰老是隨著壽命增長所發生的一個不可避免的過程,而心房纖顫(簡稱房顫,atrial fibrillation,AF)是臨床上最常見的心律失常之一,也是一種“老年病”,其發病率隨年齡增長而增加,但目前治療效果仍不理想,其發病機制需要深入研究。心房結構重塑是AF 的主要發病機制之一,心房纖維化是其主要特征。研究表明衰老與心房纖維化密切相關,與年齡較大者(>70歲)相比,年齡較小者(<50歲)右心耳纖維化明顯減少,年齡是影響纖維化發生的主要因素之一[1]。在快速起搏誘發AF 的動物模型中發現,與成年組相比,老年犬左心房纖維化更嚴重[2],但衰老相關心房纖維化的具體分子機制尚未完全闡明。
轉錄輔激活因子p300 是一種細胞內普遍存在的核磷酸蛋白,具有內在的乙酰轉移酶活性,在細胞增殖,凋亡和胚胎發育中都發揮關鍵作用[3]。研究發現,p300參與纖維化,如在特發性肺纖維化患者的肺成纖維細胞中,p300 明顯上調,而抑制p300 能減輕纖維化[4-5];p300 還影響細胞衰老,研究顯示在微血管內皮細胞中,高糖誘導的沉默信息調節因子1(silent information regulator 1,Sirt1)下調,p300 表達上調,導致細胞快速衰老[6]。我們的前期研究結果也顯示,p300 隨著體外培養的小鼠心臟成纖維細胞衰老而升高,伴隨有纖維化指標的增加[7]。因此,p300可能參與心臟衰老相關纖維化的病理過程。
Smads 是纖維化通路的重要信號分子[8],衰老通路中的重要因子p53 也與纖維化相關[9-10]。研究發現,在人腎小管上皮細胞株HK-2 和角化細胞株Ha-CaT中,轉化生長因子β1(transforming growth factor-β 1,TGF-β1)通過調控p53 活性進而促進 p53 與Smads相互作用,隨后結合到纖溶酶原激活物抑制因子1(plasminogen activator inhibitor-1,PAI-1)啟動子上,導致纖維化[11]。而 p300 可通過乙酰化調控 p53[12]。這些研究提示,p300 可通過調控p53,促進p53-Smad3相互作用進而參與衰老相關纖維化。
因此,本研究采用人心房成纖維細胞(human atrial fibroblasts,HAFs)作為研究對象,通過傳代建立細胞衰老模型,比較不同代數的細胞中p300、p53、Smad3 和其他衰老及纖維化相關因子的變化,并通過干預p300 的表達,觀察其對p53 和Smad3 等衰老和纖維化指標的影響,探索p300 在衰老相關纖維化中的作用及其可能機制,以期發現治療衰老相關心房纖維化的新靶點。
材 料 和 方 法
1 患者組織標本
本研究收集患者于心臟體外循環手術中或經胸腔鏡AF 外科消融手術中需切除的心耳組織,分離培養原代心房成纖維細胞并進行傳代培養。所有患者均簽署知情同意書,并獲廣東省人民醫院(廣東省醫學科學院)倫理委員會批準,批準號為No.GDREC2016128H。有肺炎或者其他感染性疾病的患者不入選。
2 試劑
特級澳洲胎牛血清和0.25% 胰蛋白酶-EDTA(Gibco);成纖維細胞基礎培養基(fibroblast basal medium,FBM)(Lonza);p300 小發卡(small-hairpin,sh)RNA 質粒和 p300 過表達質粒 pCMV p300 CHA(吉凱基因);Lipofectamine 3000 轉染試劑(Invitrogen);姜黃素(Cayman);蛋白 Marker(Fermentas);4×蛋白上樣緩沖液(BIO-RAD);RIPA 裂解液(強)(Beyotime);抗p300 抗體(Millipore);抗I 型膠原蛋白α1 鏈(collagen type I α1 chain,Col1A1)抗體和抗Ⅲ型膠原蛋白α1 鏈(collagen type Ⅲ α1 chain,Col3A1)抗體(Abcam);抗基質金屬蛋白酶2/9(matrix metalloproteinase-2/9,MMP-2/9),p21 和 p16 抗體(Santa Cruz);抗p53,Smad3,p-Smad3,GAPDH,PAI-1 和 TGF-β 抗體及衰老相關β-半乳糖苷酶(senescence-associated βgalactosidase,SA-β-Gal)染色試劑盒(Cell Signaling Technology);其他生化試劑均為進口分裝或國產分析純。
3 方法
3.1 細胞培養及質粒轉染 人心耳組織經PBS 漂洗后去除多余脂肪組織和內外膜,加入幾滴FBM 培養基(含10%胎牛血清),細剪成約1 mm ×1 mm ×1 mm 小塊,用吸管將組織塊均勻涂布于25 cm2培養瓶培養面上,靜置約15 min,輕翻培養瓶從側面加入4 mL FBM 培養基(含10%胎牛血清),培養面朝上于37 ℃,5%CO?的培養箱中靜置2 h,組織塊貼牢后,翻轉培養瓶,培養面朝下放置使培養基浸過所有組織塊,繼續培養。每隔5 d換液,待細胞游離、生長至基本融合成片達瓶底80%后,進行消化傳代。傳代至P3 和P7 的細胞接種于6 孔細胞培養板中,待細胞密度達70%~80%,使用Lipofectamine 3000 轉染試劑盒按說明操作進行質粒轉染。質粒稀釋液:125 μL Op-ti-MEM 培養基稀釋 2.5 μg 質粒和 5 μL P3000 試劑,輕輕混勻;脂質體稀釋液:125 μL Opti-MEM 培養基稀釋 6 μL Lipofectamine 3000 試劑,高速震蕩 3 s 混勻。然后將這兩份分別含有質粒和脂質體的溶液混合,輕輕混勻,室溫孵育15 min,均勻滴加到細胞新鮮更換的1 mL FBM 培養基(含10%胎牛血清)中,輕輕搖勻,于培養箱中繼續培養2~3 d。
3.2 SA-β-Gal 染色 β-半乳糖苷酶是衰老細胞的標志物。將細胞接種于6 孔細胞培養板中,待密度達到50%~70%,按試劑盒操作說明進行SA-β-Gal 染色。棄培養基后用PBS 漂洗1 次。加入固定液室溫固定15 min,去除固定液后PBS 漂洗2次。按照說明配制染色液,調節pH 值為6,每孔加1 mL 染色液,密封,于37 ℃,無CO2的干燥恒溫箱孵育過夜。在顯微鏡下觀察,核周藍染的即為衰老細胞。
3.3 Western blot實驗 細胞棄去培養基,PBS洗3次后加入適量含蛋白酶抑制劑的RIPA 裂解液(強),冰上裂解5 min,用刮刀刮細胞,用移液槍收集細胞混合物至 EP 管中,冰上繼續裂解 20 min。4 ℃,12 000 r/min 離心15 min,取上清分裝至EP 管,儲存于-80 ℃。BCA法測定蛋白濃度。加入4×蛋白上樣緩沖液稀釋30 μg 蛋白,55 ℃煮 10 min。行 8%~10% SDS-PAGE分離蛋白樣品,轉移到PVDF 膜上。蛋白面向上用5%脫脂牛奶于室溫封閉1 h。TBST 洗3 次,每次5 min。加入稀釋好的Ⅰ抗4 ℃孵育過夜,TBST 洗3次,每次5 min。加入稀釋好的Ⅱ抗室溫孵育1 h,TBST 洗3 次,每次5 min。蛋白面朝上均勻滴加ECL孵育30 s,平鋪在托盤上送入機器自動曝光。
3.4 細胞免疫熒光染色 待細胞密度達到70%~80%時,用預溫PBS 洗2 次。4%多聚甲醛固定15 min,PBS 浸洗 3 次,每次 10 min。0.2% TritonX-100通透20 min,4%BSA 封閉30 min,去除封閉液直接滴加稀釋好的Ⅰ抗放入濕盒,4 ℃孵育過夜。PBS 浸洗3 次,每次10 min。滴加稀釋好的熒光Ⅱ抗,濕盒中避光室溫孵育45 min。PBS 浸洗3 次,每次10 min。避光滴加封片劑(含DAPI)后在熒光顯微鏡下觀察。
4 統計學處理
采用統計軟件SPSS 20.0 分析處理數據。數據以均數±標準誤(mean±SEM)表示。兩組間比較用t檢驗。多組間比較用單因素方差分析,各組均數的兩兩比較采用 LSD-t或 SNK-q檢驗,以P<0.05 為差異有統計學意義。
結 果
1 不同代數人心房成纖維細胞SA-β-Gal 染色結果以及p300、p53 和Smad3 等衰老和纖維化相關蛋白水平的變化
體外培養的HAFs 連續傳代至P11 代,行Western blot 檢測不同代數(P3、P7 和P11)細胞的 p300、p53和Smad3等衰老和纖維化相關因子的蛋白水平,結果如圖1A、1B所示,隨著細胞代數增加,p300和衰老相關因子p53表達水平逐漸升高,以P11代細胞最高(P<0.05;P<0.01)。與P3 代細胞相比,P7 代細胞的p21 和p16 表達水平都明顯升高(P<0.01),且p16在P11 代達最高水平(P<0.01)。Smad3 是公認的促纖維化因子,隨著細胞傳代,總Smad3 有下降趨勢,但無統計學意義,p-Smad3 的蛋白水平在P7 和P11代細胞中均明顯增高(P<0.05),其他纖維化指標MMP-2/9 和PAI-1 表達均隨著代數逐漸增加,以P11代細胞最高(P<0.01),但Col1A1和Col3A1的表達在各組間無明顯差異,TGF-β 在P11 代細胞中表達較P3 和P7 有所下降,但無統計學差異。另外,分別取P3、P7和P11代細胞進行SA-β-Gal染色,觀察細胞衰老情況,結果如圖1C 所示,隨著傳代代數增加,核周藍染的衰老細胞逐漸增加(P<0.01)。由此可見,隨著 HAFs 衰老,p300 和 p53 表達增加,Smad3 被激活,一些纖維化相關因子表達增加。
2 姜黃素處理高代數人心房成纖維細胞對衰老和纖維化相關蛋白水平的影響
為了進一步探索p300 在HAFs 衰老以及衰老相關的纖維化中的作用,我們給予不同濃度(6、9 和12 μmol/L)的 p300 抑制劑姜黃素處理高代數(P7)HAFs,使用 Western blot 檢測細胞中 p300、p53 和Smad3 以及其他衰老和纖維化相關蛋白水平的變化。結果如圖2 所示,不同濃度姜黃素處理后,P7 代細胞的p300 水平呈現濃度依賴性下降(P<0.05),以12 μmol/L 濃度處理組達最低水平(P<0.01);衰老相關指標p53、p21 及p16 表達水平也呈逐漸下降趨勢,Smad3 及p-Smad3 的蛋白水平明顯下降,以高濃度處理組下降最為明顯(P<0.05);其他纖維化指標Col1A1、MMP-2、TGF-β 和PAI-1 在高濃度姜黃素(12 μmol/L)處理后,亦明顯下降(P<0.05;P<0.01);而Col3A1 和MMP-9 的蛋白水平也有下降趨勢,但差異無統計學顯著性。因此,p300 抑制劑姜黃素處理抑制高代數HAFs的p300,可明顯降低衰老和部分纖維化因子的蛋白水平,提示p300 在HAFs 衰老和纖維化中發揮著重要作用。
3 敲減p300 的表達對高代數人心房成纖維細胞衰老和纖維化相關蛋白水平的影響
為了確定p300 在HAFs 衰老相關的纖維化中發揮的關鍵作用,我們對高代數(P7)細胞進行p300 shRNA 質粒轉染敲減p300的表達,觀察衰老和纖維化相關因子的變化,結果如圖3 所示。質粒轉染后p300 的表達水平下降(P<0.05),伴隨p53、Smad3 及p-Smad3 的蛋白水平顯著降低(P<0.01),同時其他纖維化相關指標如Col1A1、Col3A1 和MMP-2 的蛋白水平也降低(P<0.05;P<0.05),MMP-9、TGF-β 和PAI-1 水平也有下降趨勢,但差異無統計學顯著性。由此說明,敲減HAFs 的p300表達可以抑制衰老及纖維化相關因子的分泌。
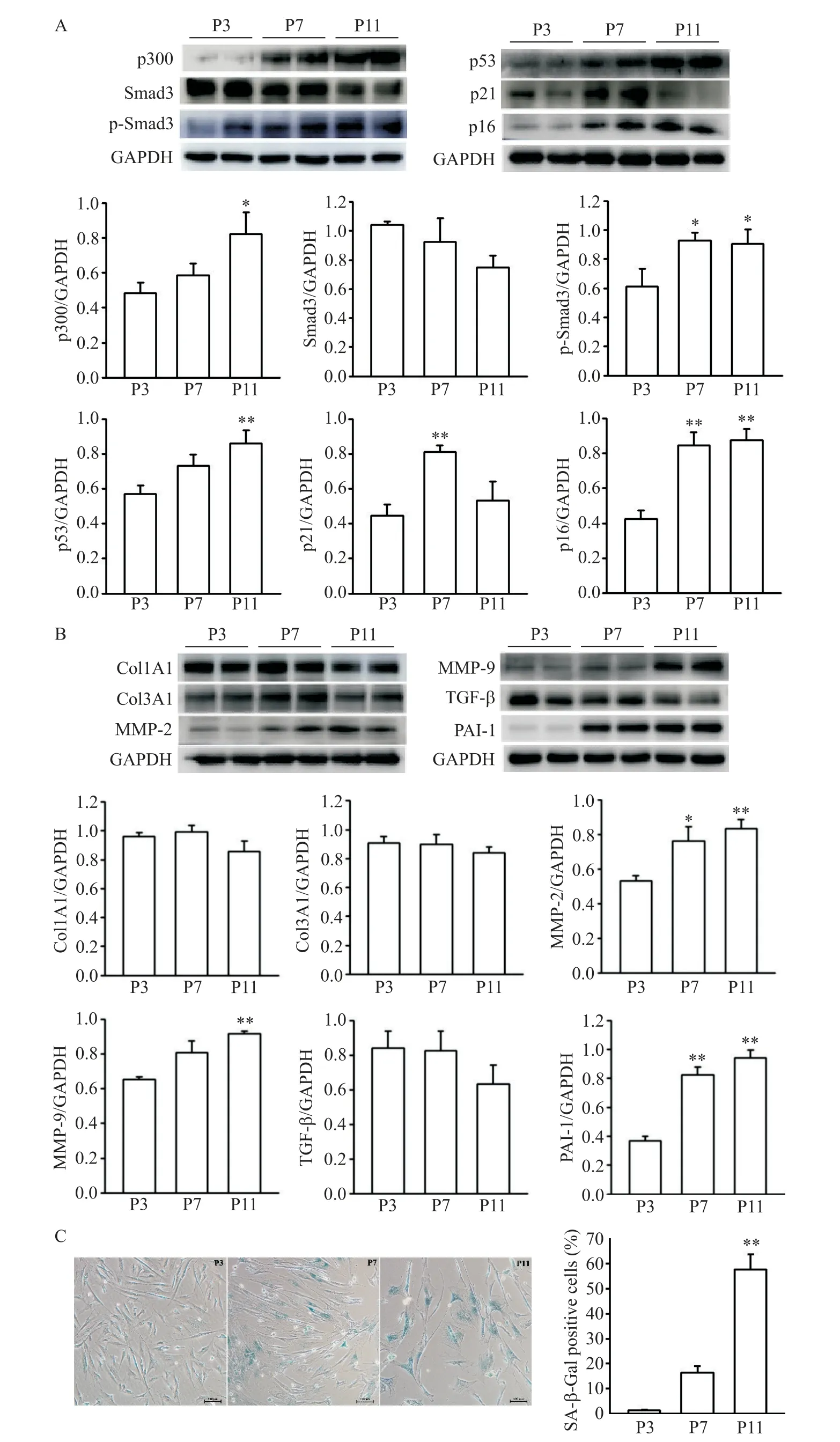
Figure 1.The results of SA-β-Gal staining and the protein expression levels of p300,senescence and fibrosis associated proteins in human atrial fibroblasts of different passages.A:the protein levels of p300,Smad3/p-Smad3,p53/p21 and p16;B:the protein expression of Col1A1/3A1,MMP-2/9,TGF-β and PAI-1;C:the images of SA-β-Gal staining(×100)and SA-β-Gal positive rate of HAFs at passage 3,passage 7 and passage 11,respectively.Mean±SEM. n=6.*P<0.05,**P<0.01 vs P3 group.圖1 不同代數人心房成纖維細胞SA-β-Gal染色結果及p300、衰老和纖維化相關蛋白的表達水平

Figure 2.Effect of curcumin on the expression of p300,senescence and fibrosis associated proteins in human atrial fibroblasts of passage 7.A:the protein levels of p300,Smad3/p-Smad3,p53/p21 and p16 before and after treated with curcumin of different concentrations(6,9 and 12 μmol/L);B:the protein expression of Col1A1/3A1,MMP-2/9,TGF-β and PAI-1 before and after treated with curcumin.Mean±SEM. n=4.*P<0.05,**P<0.01 vs DMSO group.圖2 姜黃素處理對人心房成纖維細胞p300及衰老和纖維化相關蛋白表達的影響
4 人心房成纖維細胞過表達p300 對衰老和纖維化相關蛋白水平的影響
對低代數(P3)HAFs進行質粒轉染過表達p300,觀察衰老和纖維化相關指標的變化,結果如圖4 所示,轉染后p300 的表達顯著增加(P<0.05),p53 和Smad3 的水平也明顯升高(P<0.05),纖維化指標MMP-2的水平也隨p300而增加(P<0.05),而Col1A1無明顯變化。此部分研究結果顯示,低代數的HAFs過表達p300,可促進衰老和纖維化相關因子的分泌。
討 論
以上研究結果表明,隨著HAFs 衰老,p300 及其他衰老和纖維化相關因子水平升高;姜黃素處理或p300 shRNA質粒轉染敲減p300的表達可降低p53和Smad3 的表達,抑制細胞衰老和纖維化相關因子分泌;而過表達p300 則可促進細胞衰老和纖維化相關因子分泌,說明p300 在HAFs 衰老相關纖維化中起重要作用,可能通過調控p53/Smad3通路參與其相關病理機制。
心臟纖維化的發生發展是一個復雜的過程,相關病理因素包括,遺傳因素,代謝障礙,肥胖和高血壓等。近年來研究表明,除卻病理因素,衰老亦會導致心肌細胞外基質的改變,如心臟成纖維細胞的增殖和膠原蛋白含量增加,交聯增強,導致心臟順應性下降[13]。AF 的主要病理機制包括心房肌細胞的電重塑和結構重塑,心房纖維化是結構重塑的主要表現。而進行性心房纖維化亦與心臟衰老相關[14],與年幼大鼠相比,老年大鼠的左房纖維化增加,且具有更高的 AF 誘發性[15]。因此,AF 的發生率隨年齡的增長而增加,與增齡相關的心房纖維化有關,心房組織呈現年齡依賴的間質纖維化,心肌細胞間的電通訊受阻,異位或折返活動的可能性增加,使老年人容易發生AF[15]。目前雖然衰老相關的心房纖維化研究已獲關注,但其相關機制尚未得到很好闡明。因此,本研究以HAFs 為研究對象通過傳代建立復制性細胞衰老模型,探究其相關機制。
轉錄輔激活因子p300 是體內重要的表觀遺傳調控分子,具有內的在乙酰轉移酶活性,可通過乙酰化組蛋白和非組蛋白參與調控多種基因的表達[3,16]。研究表明,p300 可調控膠原蛋白表達,促進纖維化,如在糖尿病大鼠心肌肥厚模型中,p300 通過調控 Smad2 乙酰化,增加 TGF-β 活性,促進膠原合成,導致心肌纖維化和肥厚[17];p300 HAT 特異性抑制劑—L002 在體內和體外均能有效抑制纖維化反應,如減弱體外培養的人心臟成纖維細胞分化,增殖,遷移及膠原合成能力,逆轉高血壓誘導的小鼠心肌纖維化和肥大[18-19]。雖然,直接關注p300 在細胞衰老中作用的研究不多,但在人原代包皮成纖維細胞中發現,p300 對原癌基因Ras誘導的細胞早衰至關重要,抑制p300 活性可阻止p53 活化,有助于腫瘤轉移中的衰老逃逸[20]。而在人臍帶來源的間充質基質細胞中,敲減p300的表達或抑制其乙酰轉移酶活性將誘導細胞早衰并降低增殖潛能,提示p300 通過激活p53/p21 信號途徑,在誘導細胞衰老過程中起重要作用[21]。本研究發現,p300 的表達水平隨著HAFs 傳代代數的增加而升高,同時伴隨其他衰老和纖維化相關因子增加。姜黃素處理或p300 shRNA 質粒轉染敲減p300的表達可抑制衰老信號通路和纖維化因子分泌,而過表達p300 則相反。由此證實p300 在HAFs 衰老相關纖維化中發揮關鍵作用。
本研究還對p300 參與衰老相關纖維化的分子機制進行初步探索。p53 是一種序列特異性DNA 結合蛋白,是調控細胞增殖,凋亡和衰老的重要轉錄激活子[22]。乙酰化是調控p53 功能的關鍵共價修飾,乙酰轉移酶p300 介導的p53 乙酰化不僅可增加其蛋白穩定性和轉錄活性,還可以增強其反應性基因,如細胞周期依賴性激酶抑制劑p21等的啟動子激活促進細胞周期停滯或衰老[23],這可能是p300 參與細胞衰老的重要途徑。有研究發現,高糖刺激可通過Sirt1/p300/p53/p21 途徑誘導持續的內皮細胞衰老[24]。同時,p53與Smad3都是纖維化反應的效應因子[25],在 TGF-β1 刺激腎纖維化中 p53 與 Smad3 有明顯協同作用,p53 通過與受體激活的Smads 結合而充當TGF-β1 誘導的信號傳導途徑中的輔助因子和TGF-β1 纖維化反應基因的轉錄共調節因子[26],促進腎纖維化疾病進展。本研究中也發現,p300 隨著細胞傳代而增加,且在過表達p300 時,p53 和Smad3 的活性隨之增加,并伴隨纖維化因子增加;降低p300水平后,p53 和Smad3 的水平也下降,同時纖維化因子分泌減少。因此,p300可能通過調控p53和Smad3參與衰老相關纖維化。

Figure 3.The protein levels of senescence and fibrosis associated proteins in human atrial fibroblasts of passage 7 after p300 shRNA transfection.The protein levels of p300,p53,Smad3/p-Smad3,Col1A1/3A1,MMP-2/9,TGF-β and PAI-1 in passage 7 HAFs after transfected with p300 shRNA plasmid were determined by Western blot.Mean±SEM. n=4.*P<0.05,**P<0.01 vs control group.圖3 高代數(P7)人心房成纖維細胞敲減p300的表達對衰老和纖維化相關蛋白表達的影響

Figure 4.The protein levels of senescence and fibrosis associated proteins in human atrial fibroblasts of passage 3 after p300 over-expression.The protein levels of p300,p53,Smad3,Col1A1 and MMP-2 in passage 3 HAFs after transfected with p300 over-expression plasmid were determined by Western blot.Mean±SEM.n=4.*P<0.05 vs vector control group.圖4 低代數(P3)人心房成纖維細胞過表達p300對衰老和纖維化相關因子表達的影響
此外,本研究具有以下局限性:雖然我們發現敲減p300的表達可明顯降低纖維化相關因子,如Col 1A1/3A1 和 MMP-2 的 表 達 ,同 時 p53 和 Smad3/p-Smad3 的水平亦明顯降低,而p300 過表達后p53 和Smad3 水平明顯升高,纖維化指標MMP-2 的表達亦明顯增加,但對Col 1A1 的表達無明顯影響。其原因可能是因為本研究中的HAFs來源于人體心房組織,均為接受心臟手術的患者,其心房可能均有一定程度的病理改變,在這個基礎上分離的心房成纖維細胞可能已有一定程度的纖維化,導致其膠原的表達已經比較高,故過表達p300 不能使膠原合成進一步增加。
綜上所述,本研究初步證實p300 可能通過激活p53/Smad3 通路參與人心房成纖維細胞衰老相關心房纖維化,最終導致AF的發生和發展。
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年6期)2019-10-08 08:55:48
人大建設(2019年12期)2019-05-21 02:55:32
雜文月刊(2018年21期)2019-01-05 05:55:28
人大建設(2017年6期)2017-09-26 11:50:44
學苑創造·A版(2015年11期)2016-01-14 09:03:27
俄羅斯問題研究(2012年1期)2012-03-25 09:54:45
中國火炬(2010年12期)2010-07-25 13:26:22
中國火炬(2010年8期)2010-07-25 11:34:30
