紅樹林土壤中假單胞菌群的重金屬耐受活性*
2021-02-12 10:44:20黃庶識蘇芯瑩覃仙玲
廣西科學 2021年6期
李 菲,李 喆,黃庶識,蘇芯瑩,覃仙玲**
(1.廣西科學院,廣西近海海洋環境科學重點實驗室,廣西南寧 530007;2.廣西科學院,廣西海洋天然產物與組合生物合成化學重點實驗室,廣西南寧 530007)
0 引言
目前,我國耕地土壤重金屬污染形勢嚴峻,污染面積大,且污染重金屬種類多、含量高,全國重金屬污染耕地約2 000 hm2,約占全國總耕地面積的16.7%,部分污染嚴重的耕地,重金屬含量可高達安全限值的100倍[1]。重金屬污染物的生物毒性大,易與化肥、農藥以及抗生素等有機物結合,生成更具毒性的有機金屬化合物,極大影響并危害生物體正常生長。再者,重金屬污染物吸附在土壤顆粒上,既降低土壤微生物活性、抑制植物根系對營養元素吸收,也極易被農作物吸收富集[2,3]。基于重金屬污染物的遷移、富集特性,生物體中的重金屬濃度可能倍增至生境中的上萬倍,而重金屬沿食物鏈逐級進入人體內并長期富集積累后,會對人體產生巨大危害,如鉛污染會對人體的造血、神經和消化系統有較明顯傷害[4],會導致人體四肢酸痛、貧血、神經系統器質性疾病、肝/腎損傷、心血管器質性疾病、智力下降(特別是兒童)及中樞神經和造血系統的損傷等[5];鎘污染可能會導致人體的消化系統遭受破壞,腎、肝功能失調,骨質疏松,人體器官致癌致畸[6];六價鉻污染會損傷人體呼吸系統,破壞人體細胞中DNA致使細胞畸變[7];銅污染會使人體出現惡心、嘔吐、胃燒灼感等癥狀,重者會出現腹痛、吐血、溶血性黃疸、貧血、肝大、血紅蛋白尿、急性腎功能衰竭和尿毒癥[7];人類食用鈷污染的食品,輕者脫發,重者會引起血液系統疾病[8]。
修復重金屬污染的耕地土壤,能有效地將污染源遏制在食物鏈最底端,其主要途徑有去除化和固定(或穩定)化。去除化是將土壤中的重金屬污染物提取出去;固定(或穩定)化是改變土壤中重金屬的存在形態,以消除生物毒性[9]。其中,微生物修復作為固定(或穩定)化技術中的研究熱點,主要是利用微生物對重金屬的親合、吸附、轉化效應,降低土壤中重金屬的有效濃度。微生物治理法具有成本低、效率高,一般不需要停耕,可進行原位修復,能改善土壤環境、提升土壤肥力等優勢;但該方法同時具有對重金屬固定性有限、代謝能力不佳、微生物流失或被吞噬等缺點[1,10,11]。因此,尋找耐受力強、適用范圍廣的微生物類型,可對微生物在實際應用中的強化和聯合修復工作提供更好的幫助。
假單胞菌群是一類活躍在植物根際的革蘭氏陰性菌,能產生多種抗生素,改善植物營養,促進植物生長,降解土壤中有毒物質及起防病殺蟲作用等[12]。Wang等[13]研究發現,熒光銅綠假單胞菌PseudomonasaeruginosaCW-96-1在有氧培養過程中,可以通過沉淀硫化鎘去除溶液中99%以上的Cd2+。Pramanik等[14]發現一株Pseudomonassp.K32對Cd2+、Pb2+和As3+高度耐受,在Cd2+脅迫下表現出固氮和溶磷能力且能產生生長素IAA,促進水稻(Oryzasativa)幼苗生長,同時,對多種植物病原真菌有顯著抑制作用及產生大量的功能酶。Liu等[15]研究發現,內生PseudomonasputidaRE02可定植在白三葉(Trifoliumrepens)根際和根內,對鎘、鉻和鉛金屬高度耐受,且能促進植物發芽,提高土壤肥力。Mokrani等[16]研究發現,兩株固氮假單胞菌Pseudomonasazotoformans對Cd2+和Pb2+有很強耐受性,且能拮抗多種植物病原菌。此外,鑒于紅樹林處于海水與淡水交互的特殊地帶,其沉積物中富含有機質,且具有一定的厭氧性和還原性,進而可以結合更多的重金屬,使其成為重金屬污染物的“匯”[17]。故本研究從紅樹林土壤中分離假單胞菌群,并對其開展重金屬耐受活性研究,旨在獲得對多種重金屬都有高耐受活性的菌株,并檢測菌株的修復能力,為微生物修復技術研究提供數據參考。
1 材料與方法
1.1 材料
1.1.1 紅樹林土壤樣本
2021年8月初從廣西欽州茅尾海紅樹林保護區(108°36′14″ E,21°44′53″ N)中采集3處土壤樣本,待潮水退至最低潮時,從最靠近海端的紅樹林往回隨機采集(每處間隔5 m),樣品采集深度為0.5 m,將樣品均裝于密封袋,于4℃低溫保藏備用。
1.1.2 培養基
(1)分離/純化培養基:2216E固體培養基(青島高科技工業園海博生物有限公司);改良ISP2固體培養基(麥芽提取粉2.0 g,酵母提取粉2.0 g,葡萄糖2.0 g,去離子水1 000 mL,海鹽25.0 g,瓊脂14.0 g)。
(2)重金屬耐受實驗培養基:改良ISP2培養基,BE培養基(胰蛋白胨10.0 g,去離子水1 000 mL)。分別用去離子水配制6 400 μg/mL的CoCl2、ZnSO4、CdSO4、CuSO4、Pb(NO3)2和K2Cr2O7溶液,無菌過濾后即為重金屬離子標準儲備液。
1.2 方法
1.2.1 菌株分離純化
稱取2.0 g混合樣品裝于20 mL無菌海水(添加1%的吐溫20),180 r/min搖床處理30 min后,稀釋成10-2和10-3濃度的樣液備用。取200 μL稀釋的樣液涂布至2216E固體培養基中,28℃培養7 d,挑取肉眼可見菌落進行純化培養,記錄其形態特征和菌落數,以30% (V/V)甘油-ISP2混合液作為保護劑,將純化好的菌株制成凍存管保藏于-70℃冰箱。
1.2.2 16S rRNA基因測序和系統發育分析
采用chelex-100樹脂[18]快速提取細菌的DNA作為PCR模板,并根據Walsh等[19]的方法對其進行PCR擴增。擴增和測序引物均為細菌通用引物27F和1492R,PCR反應條件參照李菲等[20]的方法設定。擴增產物經1%瓊脂糖凝膠電泳檢測合格后,委托北京擎科新業生物技術有限公司進行測序分析。序列經BioEdit Sequence Alignment Editor軟件整理后,利用EzBioCloud數據庫(https://www.ezbiocloud.net/)進行在線比對[21];選取同源性最高菌株的序列作為參比對象,運用MEGA 10.0軟件,采用Neighbor-Joining法構建系統發育樹,Boostrap 1 000次檢測各分支的置信值,對各菌株的系統發育地位進行分析[22]。
1.2.3 重金屬耐受菌的初篩
以改良ISP2或BE固體培養基為基礎培養基,分別添加6種重金屬離子標準儲備液使其終濃度為100 μg/mL,將待測菌株劃線于該培養基上,28℃培養5 d,觀察其生長狀況,并記錄其結果。
1.2.4 耐受菌的重金屬離子最大耐受濃度
根據肉湯稀釋法[23]測定重金屬離子抑制待測菌株的最小抑菌濃度(Minimum Inhibitory Concentration,MIC),即細菌對重金屬最大耐受濃度。各取60 μL重金屬離子標準儲備液分別加入96孔板的第1列,加入60 μL改良ISP2或EB(Pb2+)液體培養基,混合均勻后,吸取60 μL混合液至第2列孔內,再加入60 μL改良ISP2或EB(Pb2+)液體培養基,混合均勻后,吸取60 μL混合液至第3列孔內,以此類推,稀釋至第6列孔內。第1至第6列的重金屬離子濃度依次為3 200,1 600,800,400,200及100 μg/mL,第7列加60 μL改良ISP2或EB(Pb2+)液體培養基作為陽性對照,以各濃度下的重金屬離子溶液(不加菌)作為陰性對照,每組3個平行。用麥氏比濁管將待試菌株的菌懸液濃度調至約108CFU/mL,分別加入第1至第7列96孔板中。28℃培養5 d,用酶標儀測定其OD600值。
1.2.5 菌株的重金屬耐受基因檢測
采用1.2.2節的方法提取細菌中的DNA,對重金屬耐受基因進行擴增,退火溫度均設置為58℃,參數[24,25]詳見表1。取5 μL擴增產物進行1%瓊脂糖凝膠電泳檢測,用凝膠成像儀觀察結果并拍照。
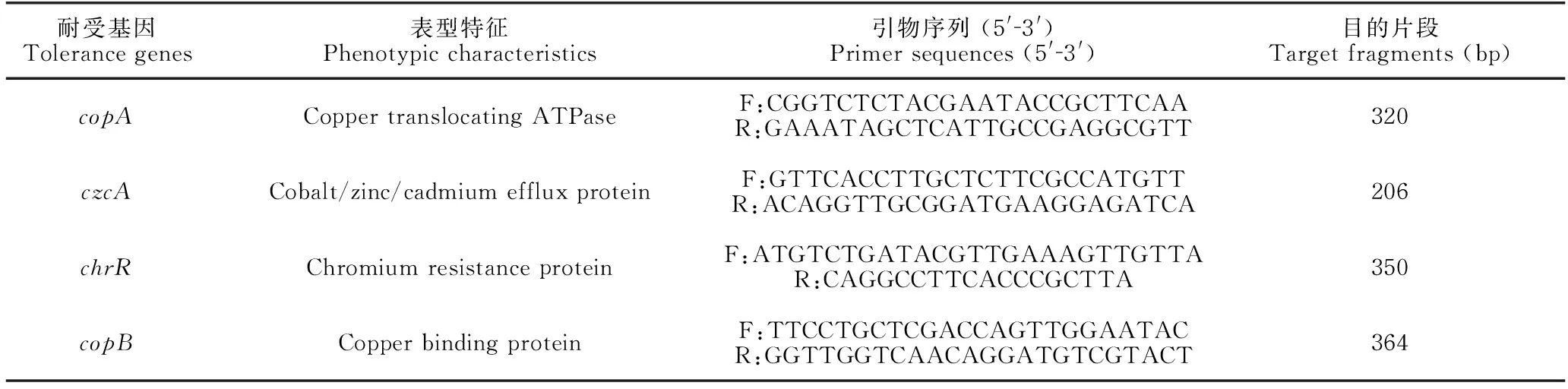
表1 假單胞菌的重金屬耐受基因檢測引物
1.2.6 菌株對重金屬離子的吸附能力
取活性顯著對數生長期菌株分別接種至100 mL改良ISP2培養基中,置于28℃、180 r/min搖床培養2-3 d,用紫外分光光度計測定培養物的OD600值,待其OD600值趨于穩定,用無菌離心管收集全部菌體,8 000 r/min離心10 min,棄上清液,收集菌體沉淀,用無菌水洗滌3次,用0.9%生理鹽水重懸菌體后置于4℃冰箱中保存14-16 h,即可得到休止細胞溶液。將休止細胞溶液置于50 mL離心管(設置3組平行),5 000 r/min離心5 min,收集菌體沉淀,稱量濕菌體重量,再用已配制好的重金屬離子溶液重懸菌體,重金屬離子終濃度為400 mg/L,28℃、180 r/min搖床中振蕩培養24 h。以接種前的重金屬離子溶液作為對照組,用原子吸收光譜儀測定重金屬離子濃度,并計算其吸附率和吸附能力,公式:吸附率=(對照組重金屬離子濃度-實驗組重金屬離子濃度)/空白組重金屬離子濃度×100%;吸附能力=溶液體積×(對照組重金屬離子濃度-實驗組重金屬離子濃度)/細胞干重,單位為mg金屬/g菌體。
2 結果與分析
2.1 紅樹林土壤中可培養假單胞菌
從廣西欽州茅尾海紅樹林保護區土壤中共獲得77株細菌,通過形態、大小、顏色等形態學特征及革蘭氏染色進行初步排重,選取42株革蘭氏陰性菌進行16S rRNA基因測序,結果共獲得23株假單胞菌,并對其構建Neighbour-Joining系統發育樹(圖1)。
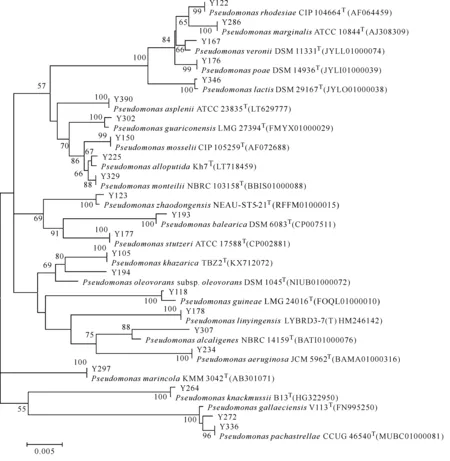
圖1 土壤中假單胞菌的16S rRNA基因序列N-J系統發育樹
2.2 假單胞菌的重金屬耐受活性初篩結果
23株假單胞菌在各重金屬培養基上生長情況如下:能在含100 μg/mL Cu2+,Pb2+,Cr6+,Cd2+,Co2+和Zn2+培養基上生長的假單胞菌分別有16,18,14,10,4和15株;總陽性率分別為69.56%,78.26%,60.87%,43.48%,17.39%和65.22%。其中,Y390,Y193和Y122均不能在各重金屬培養基上生長;Y123,Y225,Y167和Y150均在含Co2+培養基上長出少量菌落;Y286和Y336均在含Pb2+和Cr6+培養基上長出少量菌落,在其他重金屬培養基上未見菌苔。此外,重金屬培養基上生長的菌落形態發生了變化,如Cu2+培養基上,Y234的菌體顏色由淺棕色變為淺綠色,Y272的菌體顏色由微橙色變為褐色,且菌體表面較為干燥;Pb2+培養基上,Y234和Y167的菌體變為橙色,Y272的菌體變為深褐色(圖2)。
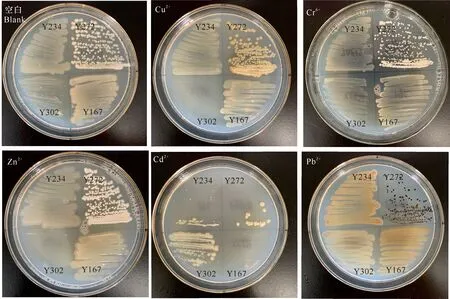
圖2 4株菌在各重金屬培養基上的生長情況
2.3 菌株的重金屬耐受性能
根據初篩結果,選出至少能在一種含有100 μg/mL重金屬培養基上生長的假單胞菌18株,進一步摸索各菌株在不同重金屬離子中最大耐受濃度。由表2可知,可耐受800 μg/mL Pb2+的菌株有7株,4株菌對Pb2+的耐受活性可達1 600 μg/mL;耐受Zn2+的假單胞菌有6株,耐受Zn2+濃度在1 600 μg/mL以上;其中,有6株菌對Pb2+和Zn2+均有顯著耐受活性,分別為Y234、Y123、Y225、Y167、Y346和Y264。除此之外,對鉛和/或鋅耐受活性顯著的11株菌對其他多種重金屬,如Cu2+、Cr6+和Cd2+,也具有一定的耐受性,這可能與其生長的紅樹林生境有關。生活在被污染土壤中的微生物往往會通過改變自身的生化和結構特征、生理特性和遺傳基因等來適應環境,從而產生各種機制應對各種復雜的環境[26],這一特性暗示這些菌株對多種重金屬復合污染的水體具有修復潛能。
對23株假單胞菌進行重金屬抗性基因檢測(表2和圖3),最常見的基因是copB(21.74%)、copA(26.09%)、czcA(13.04%)和chrR(4.35%),其中copA和copB基因與銅的抗性有關,czcA基因與鎘、鋅和鈷的抗性有關,chrR基因與鉻的抗性有關。綜合對假單胞菌在重金屬培養基上生長及攜帶重金屬抗性基因分析可知,在9株顯示copA和/或copB基因的菌株中,有7株表現出MIC (Cu2+)≥200 μg/mL。在不含copA或copB基因的14株菌中,有4株菌的MIC (Cu2+)≥200 μg/mL,3株菌的MIC(Cu2+)=100 μg/mL。23株假單胞菌中有3株檢測出czcA基因,其MIC (Zn2+)≥1 600 μg/mL;其余未檢出czcA的假單胞菌中,MIC (Zn2+)≥1 600 μg/mL的有3株,有3株菌的MIC (Zn2+)=200 μg/mL;23株假單胞菌對鎘和鈷的抗性均不顯著,MIC (Cd2+)≤400 μg/mL,MIC (Co2+)≤100 μg/mL。chrR基因僅在Y302中檢測到,其MIC (Cr6+)=100 μg/mL,其余未檢測出chrR基因的菌株中,有13株菌對Cr6+有一定的耐受性,其MIC (Cr6+)=100-200 μg/mL。
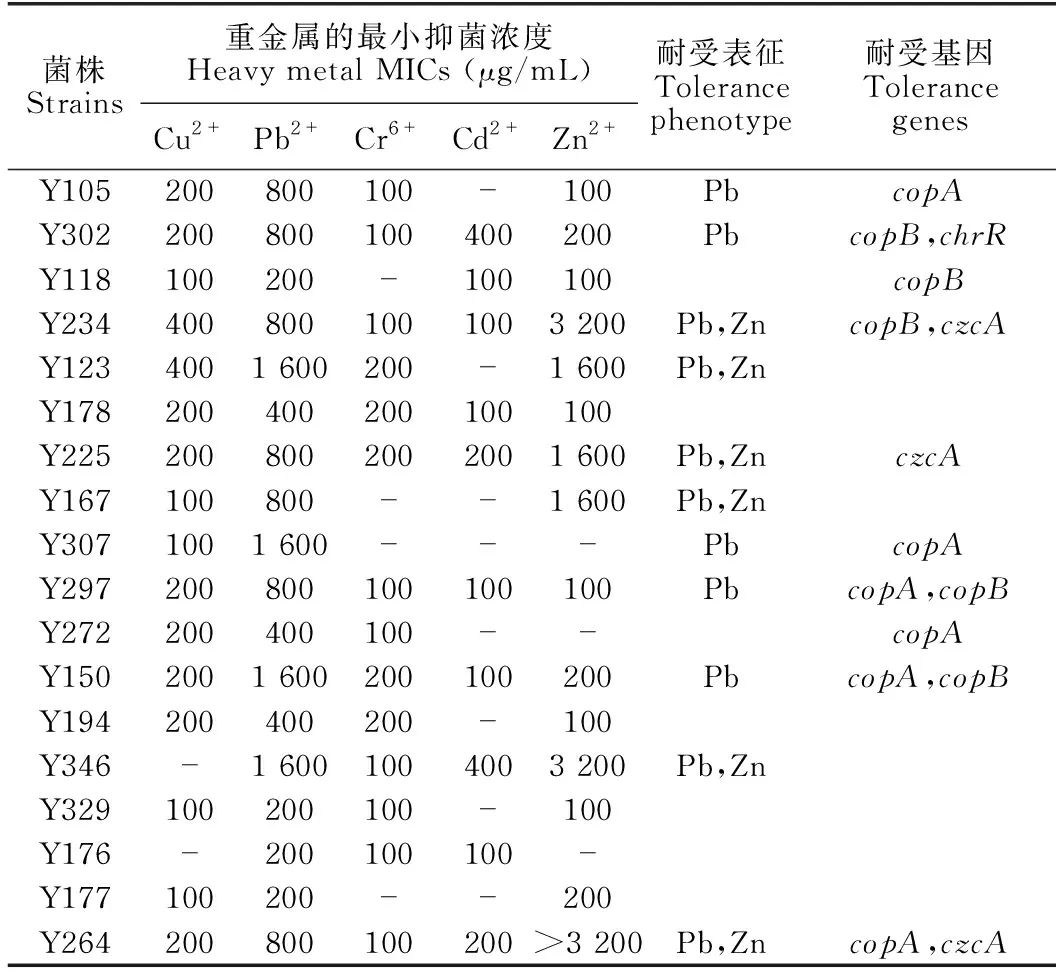
表2 18株假單胞菌的重金屬耐受活性
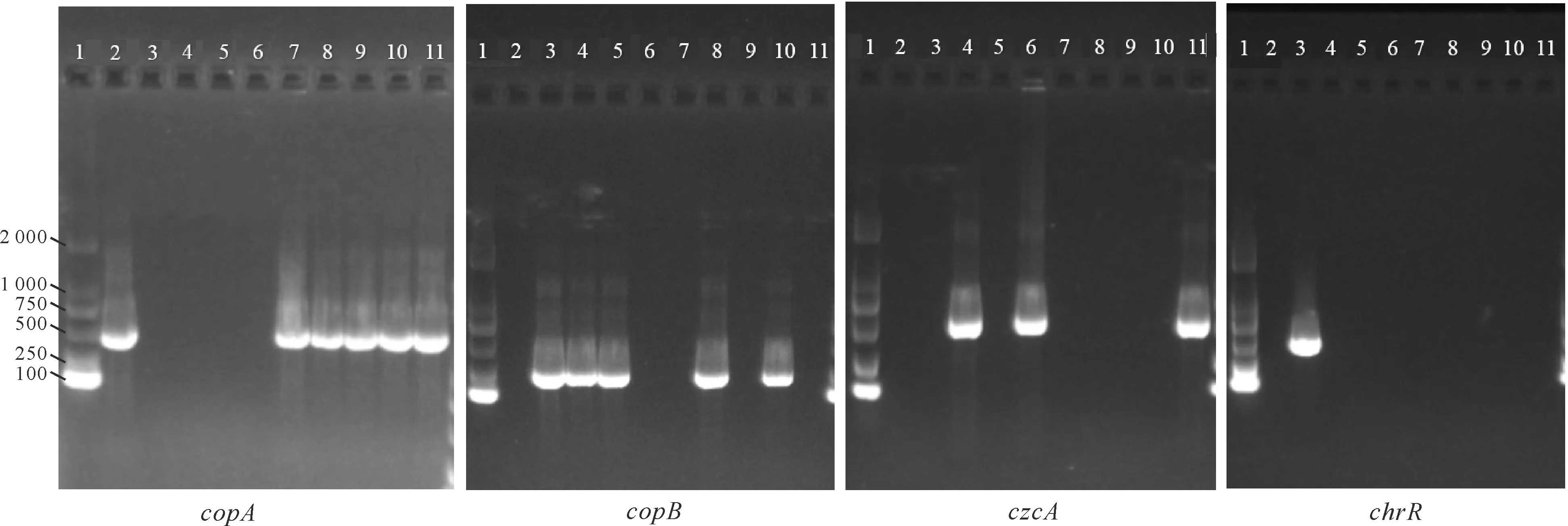
1-11號孔依次表示Marker,Y105,Y302,Y118,Y234,Y225,Y307,Y297,Y272,Y150和Y264。7條Marker指示帶從上至下表示2 000,1 000,750,500,250和100 bp
2.4 菌株對重金屬離子的吸附能力
選擇11株對鉛和/或鋅有耐受表征的假單胞菌進行吸附性能測試,在排除明顯的儀器導致的測試誤差后,獲得5株顯著吸附Pb2+和/或Zn2+的菌株(圖4)。其中,鉛的吸附率排序為Y234>Y346>Y123>Y264>Y225,鉛吸附能力排序為Y346>Y234>Y123>Y225>Y264;鋅的吸附率排序為Y123>Y264>Y234>Y225>Y346,鋅吸附能力排序為Y123>Y234>Y264>Y225>Y346。其余6株菌株對富含Pb2+和/或Zn2+的溶液基本無去除作用(吸附率均在10%以下)。吸附能力是指生物體的吸附量,在相同的菌體質量下,Y346對鉛的吸附量最大,其吸附能力最好;Y123對鋅的吸附量最大且吸附率最高,其吸附能力最強。
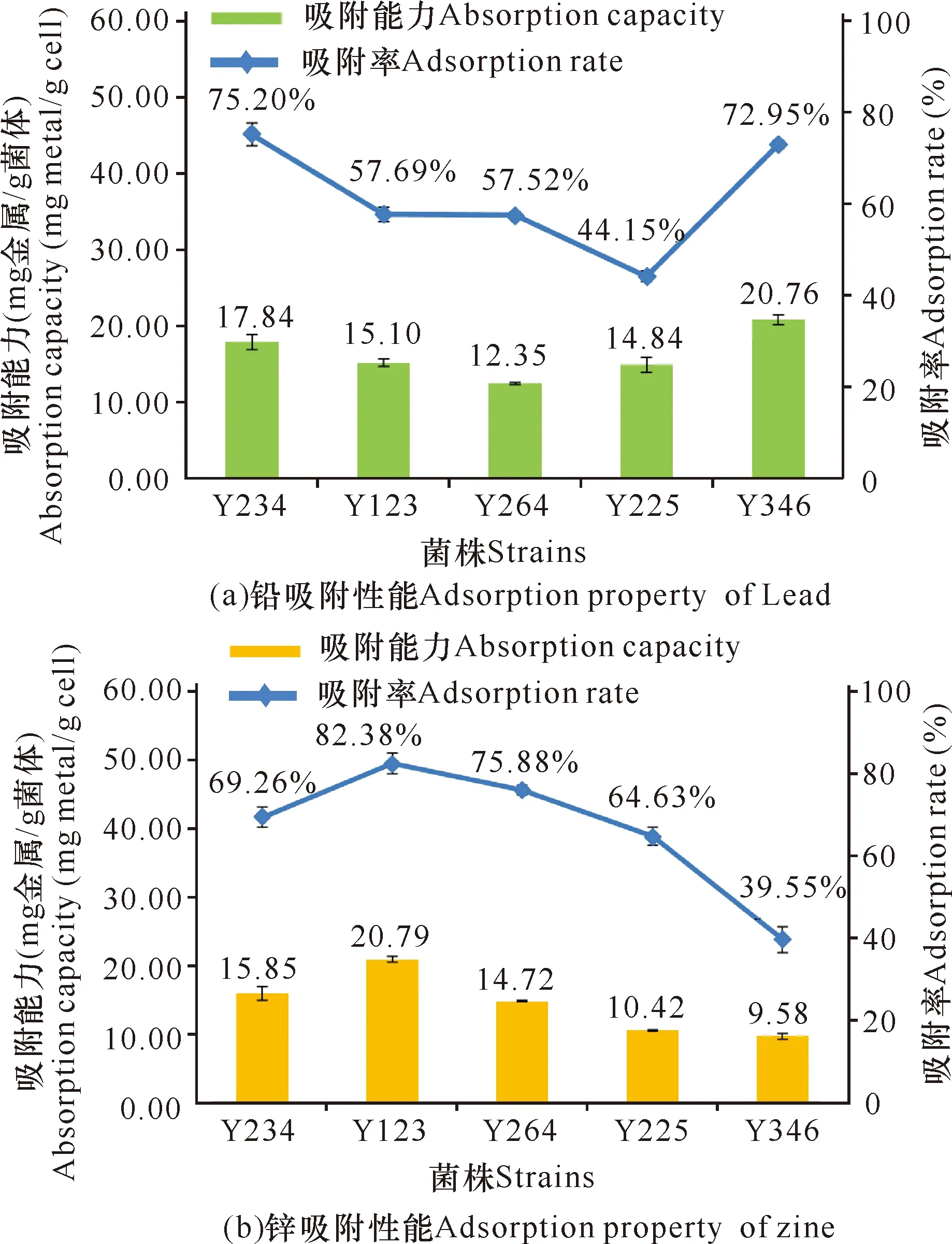
圖4 5株顯著活性菌株的吸附性能
3 討論
紅樹林微生物群落經過長時間進化,形成了特殊的生理代謝系統,在適應生態環境與自我保護的同時,通過通信、覓食等行為機制來適應特殊環境,如鹽脅迫、高礦物質組成、強還原性、強酸性、強風、強流、徑流、高溫、強輻射、頻繁的潮汐、缺氧污泥和寡營養等。獨特的生境必然會造成其微生物具有獨特的生物活性[27]。本研究選取廣西欽州茅尾海紅樹林保護區內土壤作為研究對象,從中共獲得23株假單胞菌,其中有18株菌能在多種重金屬培養基上生長,11株菌對鉛和/或鋅的耐受性顯著,5株菌對鉛和/或鋅的吸附能力突出。
耐受基因檢測中,部分菌株檢測出重金屬耐受基因,但其在高濃度重金屬培養基上卻無法生長,如檢測出copA或copB基因的菌株Y307和Y118,其對銅的耐受濃度僅為100 μg/mL;部分檢測出czcA基因的菌株對鎘和鈷的耐受性差。究其原因,可能是在發酵條件下,大部分微生物基因簇是沉默的,后續實驗將通過改變培養條件,如培養基成分、溫度、鹽度、pH、混合培養及表觀遺傳修飾劑等[28],激活微生物中的沉默基因簇,從而使之表現出較高的重金屬耐受性。然而,部分菌株雖然未檢出耐受基因,但是在重金屬培養基上仍表現出很好的耐受活性,如Y123和Y346,其耐受基因可能存在于細胞質粒中。
吸附能力實驗中,有5株假單胞菌表現出良好的鉛和/或鋅吸附性能,鉛和/或鋅的吸附率分別為44.15%-75.20%和39.55%-82.38%,吸附能力分別為12.35-20.76 mg金屬/g菌體和9.58-20.79 mg金屬/g菌體。其中,銅綠假單胞菌Y234具有很強的重金屬吸附能力,這與許多報道的結論一致[23,24,29],但因其是條件致病菌,且感染免疫力低下人群,易具有多重耐藥性及難治療等因素,因此難以廣泛應用于修復重金屬污染的環境。重金屬固定細菌是通過固定鈍化土壤/溶液中的重金屬,減少重金屬總量或者有效態含量,其作用機制主要有細胞壁吸附[30,31]、表面絡合[32]、細胞內富集[33]和胞外沉淀[34]等。后續將對4株吸附能力好的假單胞菌開展掃描電鏡-能譜觀察、傅里葉紅外光譜及X衍射分析,以研究其作用機理,為更好地開展修復重金屬污染環境工作做準備。
4 結論
廣西欽州茅尾海紅樹林保護區內土壤富含假單胞菌群資源,且部分菌株的重金屬耐受活性顯著。其中,菌株Y234、Y123、Y264、Y225和Y346對重金屬鋅和/或鉛表現出良好的吸附特性,在降解有毒物質和改善植物微環境方面具有較大潛力。