不同播深環境下玉米耐深播性狀雜種優勢及遺傳效應解析
2021-02-22 08:48:02趙小強李文麗周文期
核農學報 2021年3期
關鍵詞:效應
鐘 源 趙小強 李文麗 張 丹 周文期
(1甘肅農業大學,甘肅省干旱生境作物學重點實驗室, 甘肅 蘭州 730070;2甘肅省農業科學院作物研究所, 甘肅 蘭州 730070)
我國三分之二的玉米(Zea mays)主要種植分布于東北、華北及西北等依靠自然降水的旱作區[1],玉米播種時這些區域常年0 ~5 cm 表土蒸發量大,土壤含水量和降水量低,90%的年份因墑情不足嚴重影響了玉米適時播種及種子正常出苗生長[2-3]。目前,一般的玉米品種因其頂土能力和破土出苗能力差,在深播下即使發芽也不能把幼芽送出地表,造成玉米缺苗斷壟達40%~50%[2,4],嚴重制約玉米穩產增產。因此,培育耐深播玉米品種并及時趕墑深播,是一項拓寬玉米耐深播抗旱育種的新策略和節本高效的輕簡栽培措施。
玉米耐深播性屬于典型的數量遺傳性狀,受復雜遺傳體系控制[5-7],深入理解玉米耐深播性狀的遺傳機理,對指導玉米耐深播抗旱育種和提高玉米產量具有重要意義。目前已在玉米Bin7.04、 Bin9.03、Bin10.03 處找到調控中胚軸長的GRMZM2G133836[4]、GRMZM2G092174[8]、GRMZM2G149580[9]基因。Troyer[6]最早采用易位測交材料在玉米3、6 和9 號染色體上找到了3 個調控中胚軸長的顯性位點;后來Zhang 等[10]用F2∶3群體在10 和20 cm 播深下共檢測到40 個調控中胚軸長、胚芽鞘長、苗長及出苗率的加性數量性狀基因座(quantitative trait loci, QTLs)位點;Liu 等[4]還用IBM Syn10 DH(doubled haploid)群體在2 cm 和12.5 cm 播深下共定位到55 個調控發芽率、苗長、中胚軸長及胚芽鞘長的加性QTLs 位點。此外,曹立勇等[11]用DH 系在2 種低溫環境下共識別到7 個調控水稻(Oryza sativa)中胚軸長的加性QTLs 和2 對加性×加性上位性QTLs 位 點。Spielmeyer 等[12]還 用RILs(recombinant inbred lines)群體分析了小麥(Triticum aestivum)6A 染色體上調控胚芽鞘長的加性主效QTLs位點。從這些研究可以看出,玉米耐深播性狀QTL 定位,特別是QTL 與環境互作及上位性互作位點的深入分析還不夠深入。
在Cockerham 遺傳模型和MINQUE 法基礎上,Zhu[13]提出了研究動植物基因型與環境互作的數量性狀加性-顯性-母體遺傳模型(additive-dominantmaternal effect genetic model, ADM),為調控玉米耐深播性遺傳效應的系統解析提供了可行方法。基于以上考慮,本研究以耐深播性不同的22 份玉米親本及組配的22 份F1為試材,采用ADM 模型,分析3 種播深下玉米13 個耐深播性狀的遺傳效應;根據各性狀的雜種優勢表型及遺傳效應預測值,選用遺傳主效應表現良好并在不同播深下互作效應表現穩定的優良親本有目的地組配F1,旨在提高育種選擇效率,為玉米耐深播抗旱育種奠定理論依據。
1 材料與方法
1.1 試驗材料
以耐深播性不同的黃早四(P1)、昌7-2(P2)、H21(P3)、JW183(P4)、A413(P5)、K12(P6)、N192(P7)、W182BW(P8)、 掖478 (P9)、65232 (P10)、 Ca317(P11)、8723(P12)、TS141(P13)、TS161(P14)、廊H(P15)、A654(P16)、齊319(P17)、A154(P18)、XZX227(P19)、Q1723(P20)、8802A(P21)、綜31(P22)共22 份親本,及按不完全雙列雜交設計組配的22 份F1為試驗材料。其中P1~P8為Tangsipingtou(TSPT)種質,P15~P16為Lancaster (Lan)種質,P21~P22為Luda-Red -Cob(LRC)種質,P17~P20為P 群種質,其余為Reid 種質[3,5,14-16]。
1.2 試驗方法
采用本研究設計的種子深播試驗裝置(http:/ /epub.cnipa.gov.cn/patentoutline.action)進行深播試驗。試驗時先往該種子深播試驗裝置(高50 cm,內徑15 cm)中分層裝入滅菌蛭石,再將提前浸種12 h 的親本及F1種子各15 粒播于滅菌蛭石上,最后分別蓋3、15和20 cm 滅菌蛭石使該種子深播試驗裝置蛭石達50 cm,每隔3 d 澆20 mL 超純水,3 次重復。待種子在人工氣候室(每天光照12 h,光照強度600 μmol.s-1.m-2, 恒溫20±0.5℃)萌發10 d 后統計出苗率(seeding emergence rate,RAT)。每處理選擇長勢整體一致的5 株幼苗測定中胚軸長(mesocotyl length,MESL)、胚芽鞘長(coleoptile length, COLL)、苗長(seeding length, SDL)、根長(root length, RL)、中胚軸粗(mesocotyl coarse, MESC)、胚芽鞘粗(coleoptile coarse,COLC)、苗重(seeding weight,SDW)、中胚軸重(mesocotyl weight, MESW)、胚芽鞘重(coleoptile weight, COLW)和根重(root weight, RW),并計算中胚軸與胚芽鞘和(mesocotyl and coleoptile total length,MESL+COLL)及中胚軸與胚芽鞘比(mesocotyl length/coleoptile length ratio, MESL/COLL)。
1.3 數據統計與分析
采用IBM SPSS 19.0 (https:/ /www. ibm. com/products/spss-statistics) 中 的 GLM、 Reduction 及Correlate 模塊對3、15 和20 cm 播深下親本及F1的上述13 個性狀進行ANOVA、Pearson 及PCA(principal component analysis)分析。參照趙小強等[17]的方法估算13 個性狀的雜種優勢,即:

式中,F1為雜交種值,MP 為雙親均值,PH為高值親本值。采用隸屬函數法[5]計算3 種播深下親本及F1的13 個性狀耐深播隸屬值,并進行耐深播性綜合評價,即:

式中,Uijk為第i播深下第j材料第k性狀的耐深播隸屬值,Tijk為第i播深下第j材料第k性狀值,Tikmax為第i播深下第k性狀最大值,Tikmin為第i播深下第k性狀最小值。若所測性狀與玉米耐深播性呈正相關,則用式(5),反之用式(6)。計算各播深下所有性狀的算術耐深播隸屬值進行比較,值越大玉米耐深播性越強。參照ADM 模型分析玉米13 個性狀的遺傳效應。采用QGA Station(http:/ /ibi.zju.edu.cn/software/qga)無偏估算各性狀的方差分量占表型方差的比率及遺傳力。采用AUP(Adjusted-Unbiased-Prediction)估算各性狀的遺傳效應值[18];采用基因型為抽樣單位的Jackknife 抽樣方法[18]估算各性狀統計量的標準誤,并檢驗各遺傳參數的顯著性。
2 結果與分析
2.1 播深對玉米耐深播性狀的響應
由表1 可知,隨著播深增加,22 份親本及22 份F1的RAT、SDL、RL、MESC 和RW 整體呈降低趨勢,其余性狀整體呈升高趨勢。但不同親本及F1各性狀間的變化幅度不同,與3 cm 播深相比,15 cm 播深下親本及F1的RAT、MESL、COLL、MESL + COLL、MESL/COLL、SDL、RL、MESC、COLC、SDW、MESW、COLW 及RW 分別平均變化了22.94%、-114.12%、-48.02%、-81.32%、 - 56.00%、 14.71%、 16.26%、 8.99%、 -8.89%、-20.17%、-132.56%、-172.11%和10.02%,20 cm 播深下親本及F1的這13 個性狀分別平均變化了44.93%、 - 139.47%、 - 61.27%、 - 100.47%、 -58.94%、 15.65%、 16.73%、 6.14%、 - 15.37%、 -40.28%、-131.93%、-94.00%和16.46%。

表1 3 種播深下親本及F1 耐深播性狀比較Table 1 Comparison of deep-seeding traits of parents and F1 at three depths
2.2 玉米耐深播性狀差異分析
3 種播深下所有親本及F1的13 個性狀聯合方差分析表明(表2),親本及F1的13 個性狀在校正模型間均達到極顯著差異水平(P<0.01),說明這13 個性狀整體方差分析有效。此外,親本及F1所有性狀在基因型間及播深間均存在顯著或極顯著差異,而除SDL、RL、MESW、COLW 外,其余性狀均受基因型×播深互作的顯著或極顯著影響。說明這13 個性狀總體顯著受玉米基因型自身遺傳基礎、播深及基因型×播深互作的影響,可能這些性狀間的相互作用決定玉米耐深播性的強弱。

表2 3 種播深下親本及F1 耐深播性狀聯合方差分析(F 值)Table 2 Combine variance analysis of deep-seeding traits of parents and F1 at three depths (F value)
2.3 玉米耐深播性狀間的關系
3 種播深下所有親本和F1的13 個性狀的Pearson相關分析表明(圖1),1 個單-性狀至少與其余6 ~12性狀呈顯著或極顯著正/負相關。表明玉米這13 個性狀間的相互協同或拮抗作用共同決定了玉米的耐深播性強弱。此外,除SDL 外,其余性狀均與播深呈極顯著或顯著正/負相關(圖1)。說明玉米的這些性狀受播深影響顯著。
進一步對3 種播深下所有親本及F1的13 個性狀進行PCA 分析,結果表明(圖2),13 個主成分中前3個主成分的特征值均大于1,總體方差累計貢獻率達77.738%。表明前3 個主成分能代表玉米耐深播特性的絕大部分信息。其中決定第1 主成分的主要是MESL+COLL、MESL、MESW、SDW、RW 及RAT,可概括為玉米中胚軸、幼苗及根系綜合生長因子,解釋47.550%的貢獻率。決定第2 主成分的主要是COLL、COLC 及COLW,可概括為玉米胚芽鞘生長因子,解釋19.057%的貢獻率。決定第3 主成分的主要是SDL 和RL,可概括為玉米幼苗及根系綜合生長因子,解釋11.131%的貢獻率。
2.4 玉米耐深播相關性狀的雜種優勢分析
3 種播深下玉米13 個性狀雜種優勢分析表明(圖3),13 個性狀的平均F1雜種優勢指數介于90.97%~175.64%,CV 為16.67%;平均相對雜種優勢介于-16.49%~34.58%,CV 為99.77%;平均中親優勢介于-9.03%~75.65%,CV 為85.74%;平均超親優勢介于-17.78%~53.48%,CV 為212.96%。說明玉米這13 個耐深播性狀間的雜種優勢差異較大,今后育種應充分利用玉米這些耐深播性狀間的雜種優勢表現,盡可能地發揮相應性狀的正向雜種優勢,以便組配出強耐深播雜交種。
2.5 玉米耐深播性狀的遺傳方差分量和遺傳力分析
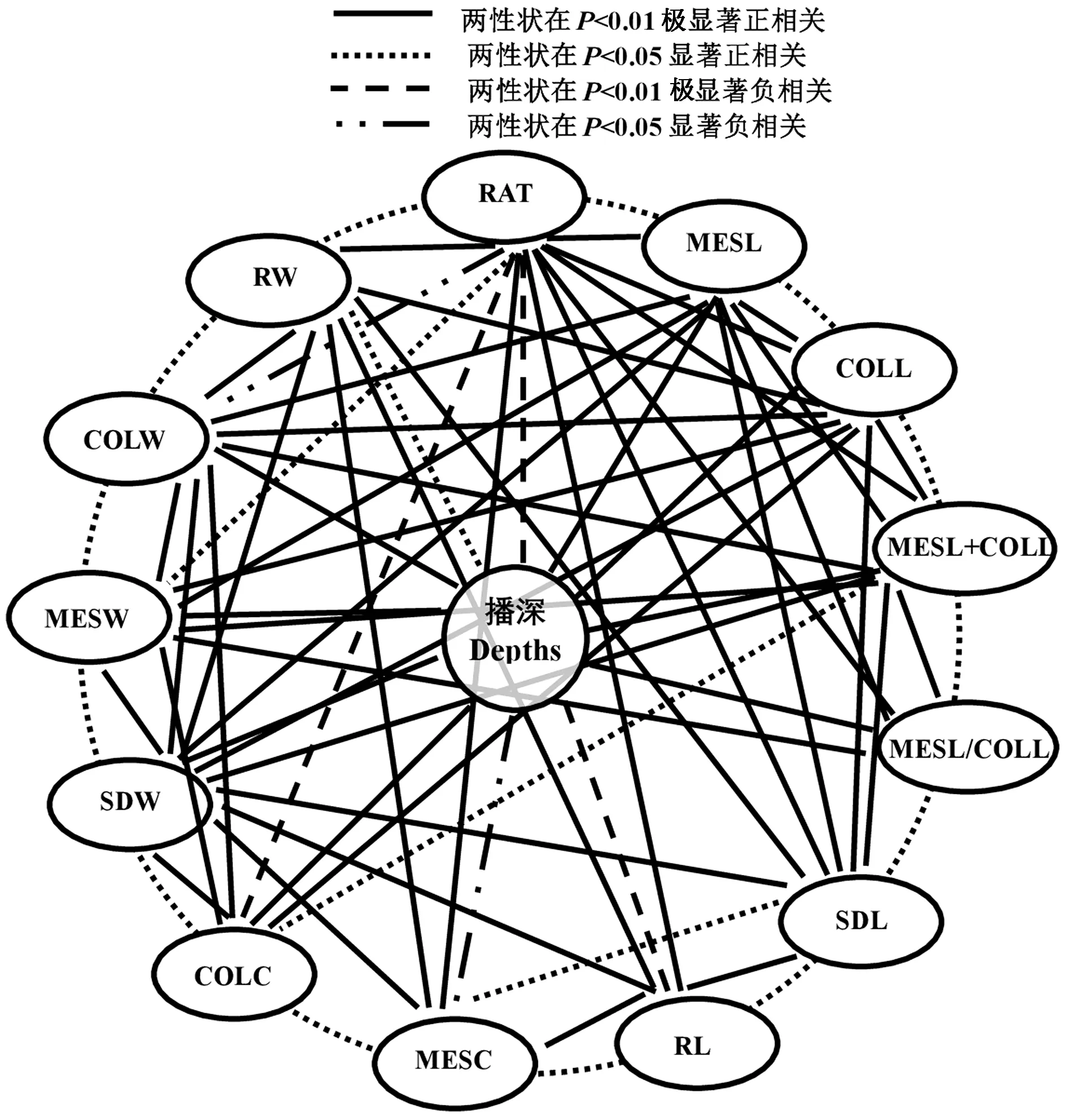
圖1 玉米耐深播性狀Pearson 分析Fig.1 Pearson correlation analysis of deep-seeding traits in maize
13 個性狀各遺傳方差分量分析表明(圖4-A),除RAT 的母體方差比率及母體×環境互作方差比率不顯著外,其余各遺傳效應方差分量比率均在P<0.01/0.05 水平下顯著,但所占比重不同。說明玉米這13個耐深播性狀同時受多種遺傳效應影響。其中MESL、COLL、MESL+COLL、MESL/COLL、RL 的加性遺傳效應占主導地位,其加性方差比率均高于顯性方差比率。此外,這些性狀的遺傳力分析表明(圖4-B),所有性狀的普通廣義遺傳力介于57.4%~85.9%,明顯高于其互作廣義遺傳力;這些性狀的普通狹義遺傳力介于10.8%~35.7%,明顯低于其普通廣義遺傳力;除RAT、SDW、MESW、COLW、RW 外,其余性狀的互作狹義遺傳力均低于其狹義遺傳力。說明玉米13 個性狀均顯著受基因加性與非加性效應及其與環境互作效應調控。
2.6 玉米耐深播性狀的配合力分析
13 個性狀的GCA 和SCA 方差分析表明(圖5),這13 個性狀在F1組合間均達到差異極顯著(P<0.01)。說明玉米這些耐深播性狀在不同F1組合間的遺傳差異真實存在。此外,所有性狀的父/母本GCA及F1組合SCA 間均差異顯著或極顯著。表明玉米這些性狀的GCA 和SCA 在親本及F1間差異真實存在,可進一步預測親本相關性狀的加性效應及F1的顯性效應。

圖2 玉米耐深播性狀PCA 分析Fig.2 Principal component analysis (PCA) of deep-seeding traits in maize
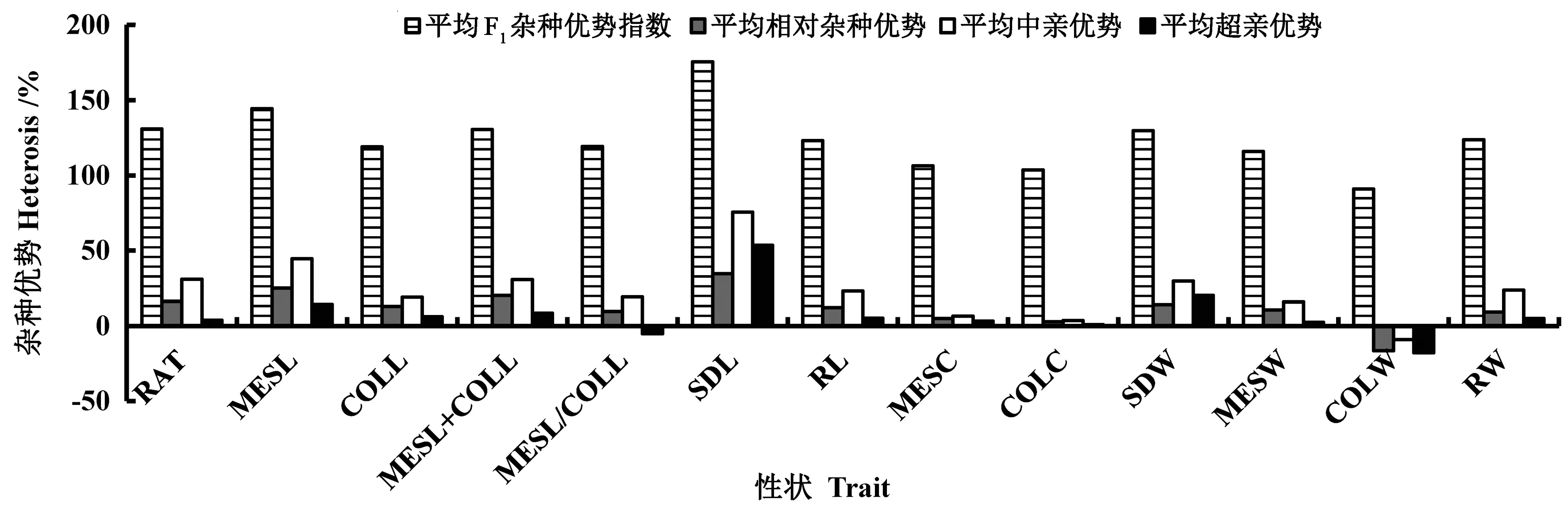
圖3 玉米耐深播性狀雜種優勢分析Fig.3 Heterosis analysis of deep-seeding traits in maize

圖4 玉米耐深播性狀的遺傳方差分量比率和遺傳力估計值Fig.4 Estimates proportions of genetic variance components and heritability of deep-seeding traits in maize

圖5 玉米耐深播性狀配合力方差分析(F 值)Fig.5 Variance analysis of combining ability of deep-seeding traits in maize (F value)
2.7 玉米親本及F1 耐深播性狀的加性和顯性效應分析
由于61.5%的玉米耐深播性狀的加性效應高于加性×環境互作效應(圖4-A),因此可對22 份親本的加性主效應進行分析。總體而言,22 份親本13 個性狀間的加性效應值差異較大(表3)。說明不同親本耐深播性狀的GCA 不同。其中8802A 的COLL、SDL、RL、MESC、SDW、MESW、COLW、RW 的加性效應表現均最高,其F1這8 個性狀分別增加13.48%、1.08 cm、14.05 cm、0.26 mm、5.90 cm、0.74 g、0.13 g、0.13 g 和0.70 g。由于玉米這13 個性狀均顯著受顯性效應及顯性×環境互作效應的調控(圖4-A),因此分析這些性狀在特定F1中的顯性效應十分必要。整體而言,22份F1這13 個性狀的顯性效應差異較大(表4),說明,特定環境下不同F1的SCA 不同。其中A413 ×K12RAT(26.51%)的正向顯性效應最大,且MESL+CoLL、MESL/CoLL、SDL 和RL 呈顯著的正向顯性效應。A413 × A154RAT 的正向顯性效應次之,為13.18%,且CoLL、MESL+COLL、CoLW 和RW 的正向顯性效應顯著。
3 討論
深播下玉米種子頂土出苗過程往往涉及諸多復雜生理代謝反應,并通過不同組織的協同或拮抗作用,最終形成玉米的耐深播響應機制。彭云玲等[3,5]和Wilhelm 等[19]研究表明,中胚軸與胚芽鞘協同伸長是深播下玉米種子頂土出苗的關鍵因素;而趙小強等[7]及Zhao 等[20]研究表明,深播下各種植物激素能調控玉米中胚軸與胚芽鞘細胞的伸長與擴大或細胞數目的增減。本研究也得到一致的結果,即播深從3 cm 增加到15 或20 cm 時,玉米親本和雜交種的中胚軸和胚芽鞘均顯著伸長,且中胚軸(126.80%)伸長幅度遠高于胚芽鞘(54.64%)(表1),致使玉米的中胚軸與胚芽鞘和(90.89%)、中胚軸與胚芽鞘比(57.22%)也協同升高;而玉米的中胚軸粗(7.56%)則略微減小,胚芽鞘粗(15.96%)略微增大。本研究還發現,不同播深對玉米幼苗、根系及其干物質積累量等也存在一定的影響,播深增加時玉米的苗長、根長及根重呈不同幅度的降低,而苗重、中胚軸重及胚芽鞘重呈不同幅度的增加。這與Sanusan 等[21]和李明月等[22]的研究結果一致。表明玉米的這些性狀可能與其耐深播性存在著十分密切的關系。本研究在3 種播深下相關性狀相關分析表明,13 個性狀彼此間及其與播深間均呈顯著正或負相關(圖1)。進一步對3 種播深下相關性狀的PCA 分析表明,前3 個主成分的特征值均大于1,累計貢獻率達77.738%。因此,前3 個主成分代表了玉米耐深播特性的絕大部分信息,且中胚軸長、胚芽鞘長、中胚軸與胚芽鞘和、苗長、根長、胚芽鞘粗、中胚軸重、胚芽鞘重、苗重、根重、出苗率是造成不同玉米耐深播性差異的主要因素。因此這13 個性狀可作物玉米耐深播性評價的主要參考指標,解析這些性狀在后代中的雜種優勢表現、遺傳效應大小及配合力強弱,為耐深播優良種質遺傳改良及耐深播品種選育奠定基礎。
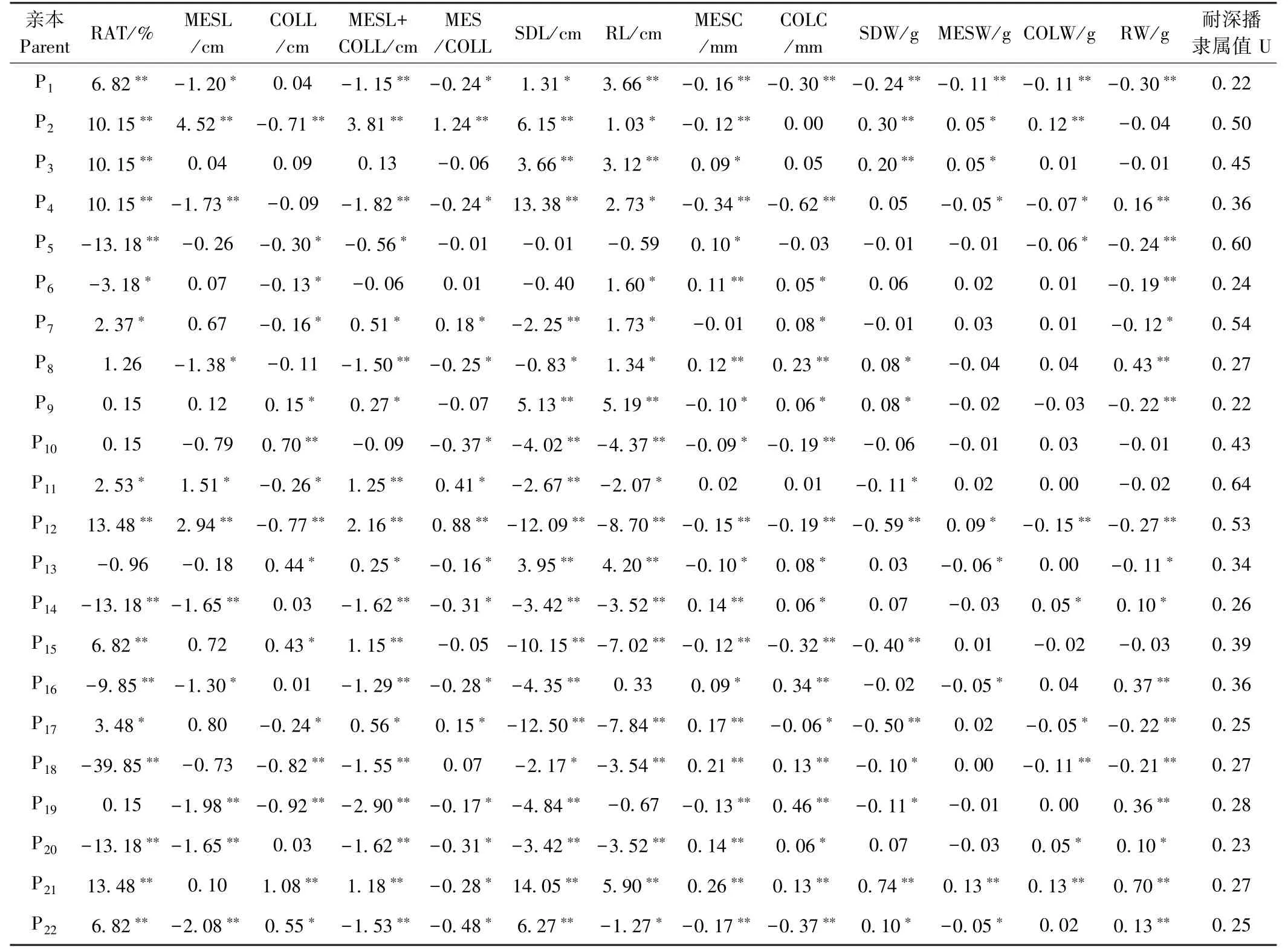
表3 玉米親本耐深播性狀加性效應及親本耐深播性綜合評價Table 3 Additive effects of deep-sowing traits in parents and comprehensive evaluation of deep-seeding tolerance in parents

表4 玉米F1 耐深播性狀顯性效應及F1 耐深播性綜合評價Table 4 Dominance effects of deep-sowing traits in F1 and comprehensive evaluation of deep-seeding tolerance in F1
作物性狀間遺傳性的差異造成其雜種優勢表現的方向和程度也不盡相同[23]。因此明確玉米耐深播性狀的雜種優勢對優良耐深播抗旱新品種培育具有重要意義。王云生等[24]研究表明,玉米親本整體耐深播性在其雜交種中的雜種優勢指數平均為197%,雜交優勢表現明顯。Zhang 等[10]分析發現,玉米中胚軸長、胚芽鞘長、苗長及出苗率的平均相對雜種優勢分別為115.74%、92.83%、92.05%和111.63%。而本研究表明,13 個性狀的雜交優勢均表現明顯,平均F1雜種優勢指數介于90.97%~175.64%;除胚芽鞘重的平均相對雜種優勢為負值(-16.49%),其余性狀的平均相對雜種優勢均為正值;除F1的平均胚芽鞘重顯著低于父母本、F1的平均中胚軸與胚芽鞘比低于高值親本,F1的平均其余性狀均顯著高于雙親,這些性狀均表現出較高的正向中親和超親優勢。李月明等[22]研究也揭示,在3 種播深下玉米品種的平均出苗率均高于父母本,表現出明顯的超親優勢,且這些玉米品種的出苗率還與其父母本呈顯著或極顯著正相關。說明玉米這13 個性狀的F1雜種優勢指數、相對雜種優勢、中親優勢及超親優勢間的差異可能是由父母本的遺傳不同引起的。因此培育綜合耐深播性強的玉米新品種時,不僅需要注重對高親、中親及母本等的選擇,還需兼顧雜種優勢的影響,以提高育種選擇效率。
作物性狀的雜種優勢是由不同遺傳效應共同作用構成的基因網絡系統形成的[25],因此系統解析玉米耐深播性狀的遺傳效應可為玉米耐深播雜種優勢遺傳機理的闡明奠定基礎。Zhang 等[10]檢測到了50.0%、33.3%、40.0%和50.0%調控中胚軸長、胚芽鞘長、苗長及出苗率的加性QTLs,檢測到了33.4%、66.7%、60.0%和50.0%調控這些性狀的部分顯性QTLs,檢測到了8.3%和8.3%調控中胚軸長的顯性和超顯性QTLs。而本研究采用ADM 模型對13 個耐深播性狀的遺傳效應分析表明,這13 個性狀同時受加性、顯性、母體、加性×環境互作、顯性×環境互作及母體×環境互作效應的影響,這些效應在不同性狀間所占比重不同。其中,中胚軸長、胚芽鞘長、中胚軸與胚芽鞘和、中胚軸與胚芽鞘比及根長的加性方差比率均高于顯性方差比率。因此這5 個性狀的加性遺傳效應占主導地位,育種過程中可通過簡單回交法或單交重組法在早代對其遺傳改良。加性×環境互作方差比率、顯性×環境互作方差比率在13 個性狀間表現不同。說明環境互作效應對玉米這些性狀的影響不同,今后應重點檢測調控這些性狀的QEIs(QTL-by-environment interactions)位點,遺傳改良這些性狀也最好在特定環境下進行,并充分發揮這些性狀在特定環境下的雜種優勢。研究還指出上位性互作效應是復雜數量性狀遺傳和雜種優勢形成的重要遺傳基礎[26-27]。之前曹立勇等[11]已在水稻上定位到2 對調控中胚軸長的加性×加性上位性QTLs位點。推測調控耐深播性狀的上位性QTLs 位點在玉米上可能普遍存在,今后應在更為復雜的遺傳背景下有效地檢測和利用玉米耐深播性狀的上位性QTLs 位點,為玉米耐深播特性雜種優勢利用提供有益參考。
親本性狀的加性效應值決定GCA 的大小,與性狀的遺傳力呈正比[28]。因此,親本加性效應值可反映親本的育種利用價值。而親本的非加性效應值決定親本組配F1的SCA,雖不能在雜交世代間穩定遺傳,但可以指導雜種優勢利用及雜交種培育[28]。本研究配合力分析發現,13 個性狀的父/母本GCA 及雜交組合SCA 間均差異顯著(圖5)。表明玉米這些性狀的GCA 和SCA 在親本及F1間差異顯著,因此預測親本這些性狀的加性效應及F1的顯性效應十分必要。就親本的加性主效應分析可知(表3),作為LRC 種質的8802A,除中胚軸與胚芽鞘比外,其余性狀均表現較高的正向加性效應,但由于其耐深播性較弱,因此可利用Reid(Ca317、8723)和TSPT(A413、N192)中的強耐深播種質改良8802A。其次,作為TSPT 種質的昌7-2和H21 其分別有9 和11 個性狀表現較高的正向加性效應,這2 個親本還表現出較強的配合力、抗旱性和耐深播性[29],因此可利用這2 份材料作為耐深播種質改良及育種的主體親本。進一步對22 份F1的13 個性狀的顯性效應進行分析(表4),結果表明A413×K12 出苗率的正向顯性效應最大,為26.51%,其中胚軸與胚芽鞘和、中胚軸與胚芽鞘比、苗長及根長還表現出顯著的正向顯性效應和較強的耐深播性,預測其貢獻主要來源于母本A413。A413×A154 出苗率的正向顯性效應次之,為13.18%,此F1其余5 個性狀的正向顯性效應顯著,而耐深播性較弱,可能受弱耐深播父本A154 的影響。JW183×綜31 中胚軸長(2.08 cm)、中胚軸與胚芽鞘和(1.53 cm)、中胚軸與胚芽鞘比(0.48)、根長(1.27 cm)、中胚軸粗(0.17 mm)、胚芽鞘粗(0.38 mm)、中胚軸重(0.05 g)的正向顯性效應顯著,此F1的耐深播性顯著強于雙親(表3 和4)。說明JW183×綜31 的耐深播性狀間GCA 較強。此外,H21×8802A 耐深播性最強,但其出苗率(-10.15%)、苗長(-3.66 cm)、根長(-3.12 cm)、中胚軸粗(-0.09 mm)、胚芽鞘粗(-0.05 mm)、苗重(-0.20 g)、中胚軸重(-0.05 g)均呈顯著負向顯性效應。因此今后在玉米耐深播新品種培育中可優先考慮綜合加性效應優良的8802A 作父本,并可在H21×8802A特定組合后代中改良創制一些優良耐深播資源。
4 結論
本研究表明,深播下玉米出苗狀況、中胚軸、胚芽鞘、幼苗及根的生長形態與有機物積累通過彼此間協同或拮抗作用將玉米幼芽送出地表;13 個耐深播性狀的雜交優勢均表現明顯,F1雜種優勢指數均大于90%,除胚芽鞘重、中胚軸與胚芽鞘比外,其余性狀均呈不同程度的正向中親和超親優勢;中胚軸長、胚芽鞘長、中胚軸與胚芽鞘和、中胚軸與胚芽鞘比及根長的加性遺傳效應占主導地位;其余性狀同時受加性與顯性遺傳主效應及其與環境互作效應的影響,13 個耐深播性狀間的雙親GCA 及F1的SCA 均差異顯著,本研究為更系統地揭示這些性狀雜種優勢形成的遺傳基礎及改良創制耐深播資源提供了參考。
猜你喜歡
核科學與工程(2021年4期)2022-01-12 06:30:26
今日農業(2020年19期)2020-12-14 14:16:52
小學生必讀(中年級版)(2020年9期)2020-12-04 02:07:22
科學大眾(2020年17期)2020-10-27 02:49:10
紅土地(2018年11期)2018-12-19 05:10:56
意林·全彩Color(2018年9期)2018-11-13 22:49:38
中學物理·高中(2016年12期)2017-04-22 11:53:03
中國衛生(2016年4期)2016-11-12 13:24:14
中國衛生(2014年4期)2014-12-06 05:57:14
小櫻桃·童年閱讀(2014年11期)2014-12-01 22:21:30
