β-1,4-葡聚糖內切酶的定向進化
2021-02-25 12:28:04郭成栓楊桂玲劉曉珊
化學與生物工程 2021年2期
郭成栓,楊桂玲,劉曉珊
(廣東食品藥品職業學院生物技術學院,廣東 廣州 510520)
纖維素酶包括β-葡聚糖外切酶、β-葡聚糖內切酶和纖維二糖酶[1-3]。纖維素酶通常在酸性條件下發揮其催化作用,在堿性條件下酶活較低甚至沒有,從而限制了其在堿性環境中的應用。堿性纖維素酶是一種缺少β-葡聚糖外切酶和纖維二糖酶的非全纖維素酶組分的β-1,4-葡聚糖內切酶,不會破壞纖維素的結構,可以在堿性條件下發揮催化作用,被廣泛應用于洗滌劑、脫墨等行業[4-6]。洗滌劑中加入堿性纖維素酶,一方面可增強洗滌劑的去污能力,有效去除棉織物上的污垢;另一方面還能對棉織物起到保護作用,避免變黃、變硬,使顏色鮮艷并保持柔軟。分離堿性纖維素酶基因、構建基因工程菌是提高堿性纖維素酶產量的重要方法之一[7-9]。此外,酶分子的定點突變和定向進化也為酶分子的改造提供了新的思路[10-13]。酶分子體外定向進化模擬自然進化,通過酶編碼基因的一輪或多輪突變,構建酶突變基因文庫,經過篩選得到所需性質的酶[14-15]。定向進化的方法主要有易錯PCR、DNA shuffling等,根據待進化的酶分子特性可選擇不同的方法。作者根據陽性轉化子在IPTG誘導下可以在LB-CMC平板上產生水解圈的原理[16-17],在初篩和搖瓶復篩的基礎上,采用易錯PCR法對β-1,4-葡聚糖內切酶基因進行定向進化,從陽性轉化子中篩選酶活提高的突變菌株。
1 實驗
1.1 試劑與儀器
PCR產物純化試劑盒、IPTG,上海生工;膠回收試劑盒,Qiagen公司;Taq DNA聚合酶、DNase Ⅰ、XhoⅠ、BamHⅠ限制性內切酶、T4 DNA連接酶、RNase A、dNTPs、DNA抽提試劑,大連寶生物公司。
熱循環儀,凝膠成像系統,低溫冷凍離心機,全溫度恒溫氣浴搖床,電熱恒溫培養箱,酸度計,垂直凈化工作臺,電子天平,滅菌鍋。
1.2 菌株、載體與培養基
大腸桿菌BL21(DE3)、表達載體pET20b,中科院昆明動物研究所;pMD18T,大連寶生物公司。
LB培養基[18]。
1.3 β-1,4-葡聚糖內切酶的易錯PCR擴增
參照文獻[18],提取短小芽孢桿菌DNA,克隆編碼β-1,4-葡聚糖內切酶的基因,構建表達載體pET20b,轉化大腸桿菌感受態細胞,得到重組菌BL21(DE3)/pET20b-EglA;在PCR反應體系中添加不同量Mn2+,進行易錯PCR擴增,通過酶活比較篩選最佳的Mn2+添加量;參照TaKaRa公司的說明書進行易錯PCR擴增產物DNA的酶切及連接;按Qiagen公司試劑盒說明進行易錯PCR擴增產物DNA的回收。
1.4 易錯PCR突變文庫的構建及陽性轉化子的篩選
設計一對引物5′-ATCTGGATCCATGCACATTTTTG-3′、5′-ATCGCTCGAGTTATTTATTCGGAAG-3′,分別引入BamHⅠ和XhoⅠ進行雙酶切,對酶切片段進行鑒定[18],構建易錯PCR突變文庫。根據陽性轉化子在IPTG誘導下可以在LB-CMC平板上產生水解圈的原理進行初篩,挑選水解圈與菌落直徑比值較大的陽性轉化子進行搖瓶復篩[19],以篩選出酶活提高的突變菌株。
1.5 β-1,4-葡聚糖內切酶重組菌的誘導表達及酶活測定
挑取BL21(DE3)/pET20b-Mut-EglA及BL21(DE3)/pET20b-EglA單菌落按1%的接種量接種至含50 μg·mL-1氨卞青霉素的LB液體培養基中,37 ℃振蕩培養至OD600值在0.5~1.0之間;加入誘導物IPTG至終濃度為1 mmol·L-1,37 ℃誘導一定時間,取樣測定酶活[19]。
1.6 突變酶與野生酶的表達量分析
將BL21(DE3)/pET20b-Mut-EglA和BL21(DE3)/pET20b-EglA分別于LB培養基中37 ℃培養10 h,按1%的接種量接種至LB液體培養基中,37 ℃培養至對數期,加入誘導物IPTG后繼續培養4 h,10 000 r·min-1離心5 min,收集菌體,用2×SDS上樣緩沖液于沸水浴中加熱5 min裂解菌體,離心,取上清進行SDS-PAGE分析,利用BandScan軟件測定突變酶及野生酶的表達量。
1.7 突變酶基因DNA及氨基酸序列分析
提取酶活提高突變菌株的重組質粒,進行DNA序列測定,在GenBank數據庫中,利用BLAST軟件對突變酶基因進行DNA及氨基酸序列分析。
2 結果與討論
2.1 易錯PCR堿基錯配的引入
PCR反應體系中的Mg2+或Mn2+不僅會使Taq DNA聚合酶的保真性大大降低,還會影響酶活。Mn2+添加量對易錯PCR產物的影響如圖1所示。
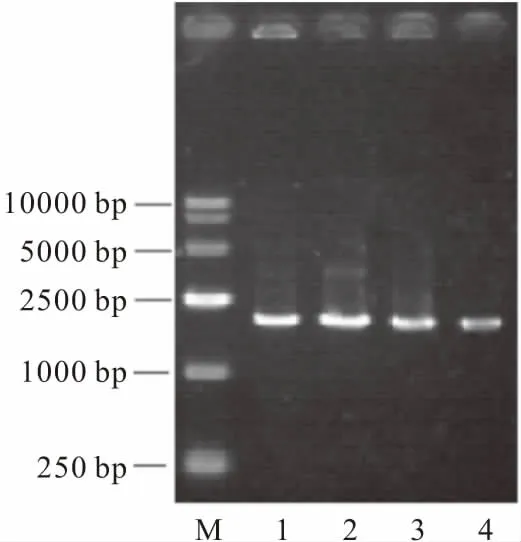
M.DL5000 DNA marker 1.Mn2+添加量1 mL 2.Mn2+添加量2 mL 3.Mn2+添加量3 mL 4.Mn2+添加量4 mL
從圖1可以看出,隨著PCR反應體系中Mn2+添加量的增加,DNA條帶亮度減弱,說明催化合成的DNA量減少,突變率隨著Mn2+添加量的增加而升高。
將添加不同量Mn2+擴增得到的PCR產物進行純化、回收、酶切、連接表達載體,轉化大腸桿菌,構建突變文庫,根據平板上陽性轉化子比率篩選合適的Mn2+添加量。結果表明,50 μL反應體系加入1 μL Mn2+,陽性轉化子比率在60%左右,對建立隨機突變體系較合適。若Mn2+添加量過多,反應體系的突變率太高,陽性轉化子太少,此時大部分為負向突變,不利于篩選到正向突變菌株。
2.2 高酶活菌株的篩選
將易錯PCR擴增得到的DNA片段構建到表達載體pET20b上,轉化大腸桿菌,在IPTG誘導下,陽性轉化子周圍可以看到清晰的水解圈(圖2a);挑選水解圈與菌落直徑比值較大的陽性轉化子進行搖瓶復篩(圖2b),可以看到,大部分的陽性轉化子都發生了負向突變,有些陽性轉化子由于發生錯配的堿基較多,從而失去了產酶活性,只有少部分的陽性轉化子發生了正向突變。

圖2 陽性轉化子在LB-CMC平板上產生的水解圈
選取5株水解圈和菌落直徑比值較大的陽性轉化子,經過1輪易錯PCR的定向進化,測得酶活(U·mL-1)分別為3.42、4.78、4.02、4.23、4.15,其中4株突變菌株所產酶的酶活得到提高,BL21(DE3)/pET20b-Mut-EglA2所產酶的酶活達到4.78 U·mL-1,為野生菌株BL21(DE3)/pET20b-EglA所產酶(3.60 U·mL-1)的1.32倍。
2.3 突變酶與野生酶的表達量分析
對BL21(DE3)/pET20b-Mut-EglA和BL21(DE3)/pET20b-EglA發酵產物進行SDS-PAGE分析,結果如圖3所示。
從圖3可以看出,誘導4 h后,菌株BL21(DE3)/pET20b-Mut-EglA和BL21(DE3)/pET20b-EglA均在73 kDa左右有β-1,4-葡聚糖內切酶目的蛋白的表達條帶。利用BandScan軟件測定蛋白條帶的相對含量,發現突變酶的表達量比野生酶提高了5%,而酶活卻提高了1.32倍,說明突變酶的催化效率得到了提高。
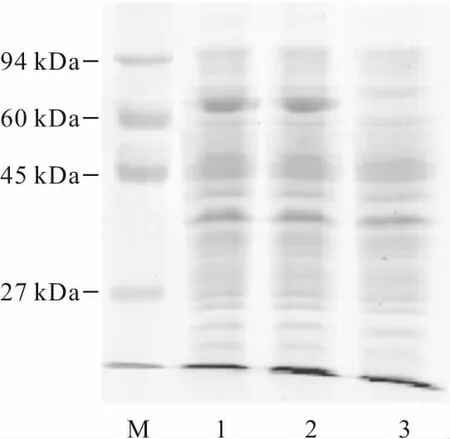
M.molecular mass marker 1.誘導4 h后的BL21(DE3)/pET20b-Mut-EglA 2.誘導4 h后的BL21(DE3)/pET20b-EglA 3.未誘導的BL21(DE3)/pET20b-EglA
2.5 突變酶與野生酶氨基酸序列的比較
基因測序結果表明,重組基因片段長度為1 980 bp。對DNA序列分析可知突變酶基因序列中有3個堿基發生了突變,其中2個為胸腺嘧啶堿基突變為胞嘧啶堿基,突變位點分別位于1 760 bp和1 962 bp處,導致酶的氨基酸序列中2個氨基酸發生了改變:其中野生酶突變位點的密碼子AAA(106 bp)突變為GAA,其編碼的氨基酸由賴氨酸變為谷氨酸;野生酶突變位點的密碼子TTT(1 760 bp)突變為TCT,其編碼的苯丙氨酸變為賴氨酸;而1 962 bp處密碼子GGT突變為GGC,但是其編碼的氨基酸沒有發生改變,為同義突變。
2.6 討論
應用PCR方法對酶分子進行改造,每一輪反應堿基突變太多會造成負向效應,一般要經過多輪反應才能將有益突變進行積累,使進化向正向突變定向進行,從而取得較好的效果。
一級序列差異很大的蛋白質能折疊成相似的三維結構,而僅有少數氨基酸差異的蛋白質卻呈現不同的三維結構,蛋白質空間結構形成的規律還遠未被認知,如何根據蛋白質的氨基酸序列折疊出其三維結構是當前研究的核心問題。突變酶氨基酸改變太少,其空間結構一般無法從三維結構上觀察到。突變酶活性的提高可能是由于N端氨基酸的改變導致酶分子空間結構發生了改變,進而提高了其催化效率;也可能是由于C端結合結構域的氨基酸改變使空間結構發生了改變,導致酶分子周圍底物濃度增大,從而使底物更容易與催化結構域結合,進而提高其催化效率[20-22]。
3 結論
根據陽性轉化子在IPTG誘導下可以在LB-CMC平板上產生水解圈的原理,在初篩和搖瓶復篩的基礎上,采用易錯PCR法對β-1,4-葡聚糖內切酶基因進行定向進化,從陽性轉化子中篩選酶活提高的突變菌株。突變酶活性是野生酶的1.32倍,催化效率約為野生酶的1.26倍。突變酶基因DNA序列中有3個堿基發生了突變,導致氨基酸序列中有2個氨基酸發生了改變:野生酶突變位點的密碼子AAA(106 bp)突變為GAA,其編碼的氨基酸由賴氨酸變為谷氨酸;野生酶突變位點的密碼子TTT(1 760 bp)突變為TCT,其編碼的苯丙氨酸變為賴氨酸;野生酶突變位點的密碼子GGT(1 962 bp)突變為GGC,但是其編碼的氨基酸沒有發生改變,為同義突變。
