NKG2D配體在 NK細胞殺傷三氧化二砷誘導的KG1a細胞中的作用①
2021-03-01 03:27:28王艷婕周彥生牛新清
中國免疫學雜志 2021年3期
王艷婕 曹 威 周彥生 楊 瑞 牛新清
(新鄉醫學院第二附屬醫院,新鄉 453003)
化療是治療白血病的有效方法,但仍有許多白血病患者不能完全緩解或緩解后復發。白血病患者復發的主要原因是抗癌藥物不能完全消除所有的白血病細胞。細胞在治療期間或治療后留在患者體內的情況被稱為微小殘留病變(minimal residual disease,MRD),這些細胞通常是白血病干細胞(leukemia stem cells,LSCs)。LSCs的細胞表面標記表型為CD34+/CD38-,與造血干細胞(hematopoietic stem cells,HSCs)具有相同的特性,且難以消除。許多研究證明LSCs在化療誘導后對化療藥物的耐受性較正常的白血病細胞更強,因此更容易導致患者復發[1-2]。臨床上高劑量阿糖胞苷、米托蒽酮、氯丙嗪、氟達拉濱等藥物聯合使用用于單純復發治療,這種再誘導減少了腫瘤的爆發,但不能長期維持[3]。現代細胞治療已廣泛應用在許多血液病惡性腫瘤中,特別是復發或難治性的血液病。供體淋巴細胞輸注或二次造血干細胞移植是一種標準的再誘導后免疫治療,但一直沒有取得較好效果[4]。NK細胞是先天免疫應答的重要組成部分,能夠殺傷腫瘤和病毒感染細胞,因此體外活化NK細胞輸注已經成為一種有希望的骨髓治療方法。
ATO作為砷劑的主要成分,是治療急性早幼粒細胞白血病(acute promyelocytic leukemia,APL)的有效藥物。大量文獻證明,ATO是一個多靶點、多通路的抗腫瘤藥物,其治療白血病的機制與誘導細胞分化、凋亡有關,具有有效的抗腫瘤作用[5]。故本研究探討聯合低濃度的三氧化二砷(arsenic trioxide,ATO)與NK細胞,對急性髓系白血病KG1a細胞株細胞增殖的影響及可能的作用機制。
1 材料與方法
1.1材料 人類急性髓系白血病 KG1a 細胞株為本實驗室留存細胞。ATO購自哈爾濱伊達藥業有限公司。LDH殺傷活性檢測試劑盒購自Promega公司。RPMI1640為Gibco公司產品。胎牛血清購自杭州四季青生物公司。抗 MICA/B單抗、抗ULBP1單抗、抗ULBP2單抗、抗ULBP3單抗購自R&D公司。甲基纖維素、XTT、硫酸酚嗪甲脂(PMS)為Sigma公司產品。淋巴細胞分離液為上海試劑二廠產品。
1.2方法
1.2.1細胞培養 KG1a 細胞培養于RPMI1640細胞培養液(含10%胎牛血清、1%青鏈霉素),37℃、5%CO2飽和濕度培養箱中培養。24 h細胞定期換液,取生長狀態良好的生長對數期細胞進行實驗。
1.2.2XTT檢測不同濃度ATO 對KG1a細胞24 h、48 h的細胞生長抑制作用 選取生長狀態良好的對數期KG1a細胞,調整細胞濃度5.0×104個/ml,96孔板每孔接種80 μl,加入不同終濃度為0、1.0、2.5、5.0、10.0、20.0、40.0 μmol/L的ATO,設3個復孔。細胞培養箱培養24、48 h后,每孔加入XTT(終濃度0.2 mg/ml)及PMS(終濃度25 μmol/ml)混合液20 μl,避光孵育2 h后,酶標儀雙波長450 nm、630 nm檢測OD值,每組吸光度取3孔均值,計算不同ATO濃度下的KG1a細胞生長抑制率。細胞生長抑制率=(OD對照組-OD加藥組)/OD對照組×100%。
1.2.3LDH檢測試劑盒檢測定NK細胞對ATO作用前后KG1a細胞的殺傷作用 3組實驗組分別檢測NK細胞對ATO處理前、后KG1a細胞的殺傷作用以及NKG2D單克隆抗體預處理后的NK細胞對ATO處理后KG1a細胞的殺傷作用。實驗前抽取健康志愿者外周血,在無菌條件下淋巴細胞分離液梯度密度離心法分離外周血單個核細胞,PBS洗滌后用免疫磁珠分選NK細胞[6]。NKG2D阻滯實驗中將免疫磁珠分選出的NK細胞與抗NKG2D 抗體(10 μg/ml)37℃共孵育30 min,然后加入到靶細胞中。細胞殺傷實驗按照LDH檢測試劑盒操作說明進行實驗,靶細胞分別是工作濃度為IC30的ATO 作用前后的KG1a細胞,效應細胞分別為NKG2D單克隆抗體預處理前后的NK細胞,3個實驗組依次按照效靶比5∶1、10∶1、20∶1 加入 96 孔板,每組設3個復孔。37℃、5%CO2飽和濕度細胞培養箱培養4 h,容積糾正組及靶細胞最大裂解釋放組均加入10 μl裂解液繼續孵育,1 h后吸取上清轉到另外一個96孔板,加顯色混合液常溫避光反應30 min,酶標儀檢測波長490 nm的吸光度值。計算NK細胞殺傷率:NK細胞殺傷率(%)=(實驗組OD平均值-靶細胞自然裂解釋放組OD 平均值-效應細胞自然裂解釋放組OD平均值)/(靶細胞最大裂解釋放組OD平均值-靶細胞自然裂解釋放組OD平均值)×100%。
1.2.4甲基纖維素集落形成實驗檢測不同實驗組集落數量 實驗設置單純對照組和3組實驗組。對照組為生長狀態良好的KG1a細胞,實驗組分別是:ATO作用24 h后的KG1a細胞組、NK細胞與ATO作用24 h后的KG1a細胞組、NKG2D單克隆抗體預處理后的NK細胞與ATO作用24 h后的KG1a細胞組。ATO作用KG1a細胞24 h后用PBS清洗細胞3次,按照效靶比20∶1于含有20%的胎牛血清、0.9 %的甲基纖維素的克隆體系中培養14 d,觀察集落形成,計數集落數量。實驗重復3次。
1.2.5流式細胞術檢測ATO作用前后KG1a細胞表面NKG2D 配體表達 該實驗分兩組,實驗組為生長狀態良好的KG1a細胞,對照組為2.0 mol/L的ATO處理24 h后的KG1a細胞。收集細胞并調整細胞濃度為1×106個/ml,100 μl待標記細胞與ABMO-1、M295、M310 和M551 一抗4 ℃避光孵育30 min,PBS洗滌2遍后細胞重懸,加熒光素標記的山羊抗鼠IgG1二抗,4℃條件下孵育30 min,陰性對照為同型IgG1 抗體。流式細胞儀分析1×104個細胞中陽性細胞數量并計算其百分率。為減少誤差,實驗重復3次。
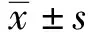
2 結果
2.1ATO對KG1a細胞生長抑制作用 分別用0、1.0、2.5、5.0、10.0、20.0、40.0 μmol/L濃度的ATO處理KG1a細胞24 h、48 h,結果顯示ATO對KG1a 細胞具有明顯的殺傷作用,其細胞生長抑制率在一定濃度范圍內呈明顯的時間濃度依賴性,差異有統計學意義(P<0.05)。根據SPSS23.0軟件計算ATO抑制30%KG1a細胞24 h的濃度,即IC30值為2.0 μmol/L。選取該濃度作為與 NK細胞聯合殺傷KG1a細胞的后續工作濃度。見圖1。

圖1 ATO對KG1a 細胞增殖抑制作用
2.2NK細胞對ATO 作用前后KG1a細胞的殺傷率 效靶比分別為5∶1、10∶1、20∶1 時,單純對照組,NK細胞對ATO 作用前的KG1a細胞的殺傷率分別為(1.74±0.91)%、(2.49±0.87)%、(7.05±2.41)%;實驗組1,NKG2D單克隆抗體預處理后的NK細胞對工作濃度為2.0 μmol/L的ATO作用后的KG1a細胞的殺傷率分別為(1.86±0.94)%、(2.56±0.88)%、(7.19±2.54)%;實驗組2,NK 細胞對2.0 μmol/L的ATO作用后的KG1a細胞的殺傷率分別為(4.95±0.65)%、(7.32±1.12)%、(19.87±1.87)%。結果顯示,NK 細胞對 ATO 作用前后的KG1a 細胞殺傷率之間的差異具有顯著統計學意義(P<0.05),NKG2D單克隆抗體預處理前后的NK 細胞對ATO作用后的KG1a 細胞殺傷率之間的差異有顯著統計學意義(P<0.05)。這提示NK細胞對ATO作用后的KG1a 細胞殺傷作用明顯,且與NKG2D配體表達密切相關。見圖2。

圖2 NK細胞對ATO作用前后KG1a細胞的殺傷敏感性
2.3ATO單用及聯合NKG2D單克隆抗體預處理前后NK細胞對KG1a細胞集落形成的影響 不同處理組于甲基纖維素集落系中培養KG1a細胞14 d后用倒置顯微鏡觀察細胞集落形成。結果顯示,組1:對照組KG1a集落數174.3±8.6,組2:ATO 處理組集落數為112.5±5.1,組3:ATO處理24 h后與NK細胞聯用組集落數48.6±7.2,組4:ATO處理24 h后與NKG2D單克隆抗體預處理后NK細胞聯用組集落數為108.5±4.9。組間兩兩比較,組1與組2、組2與組3、組3與組4組間集落數差異有統計學意義(P<0.05)。見圖3。

圖3 組間集落數比較
2.4ATO作用前后對KG1a細胞 NKG2D 配體表達的作用 單純對照組與實驗組(2.0 μmol/L的LATO處理KG1a細胞24 h后)KG1a細胞表面NKG2D 配體MICA/B、ULBP2、ULBP的表達未見明顯改變;而實驗組ATO處理KG1a細胞24 h后KG1a細胞表面NKG2D 配體ULBP1 表達增高,且差異有統計學意義(P<0.05)。見表1,圖4、5。

表1 ATO作用KG1a細胞24 h前后KG1a細胞NKG2D配體表達率

圖4 ATO作用前KG1a細胞的MICA/B、ULBPI-3表達

圖5 ATO作用24 h后KG1a細胞的MICA/B、ULBPI-3表達
3 討論
目前,白血病主要的治療方法包括化療、放療、細胞免疫治療以及這些方法的結合。傳統中藥及其成分也顯示出較強的抗腫瘤作用。如果能更廣泛地探索這些藥物在癌癥治療中的作用機制,其將有可能用于臨床靶向治療。因此,探索其分子機制對化療治療具有極其重要的意義。
NK細胞作為先天免疫應答的一部分,可以通過一系列受體來識別和殺傷病理細胞。這些受體可分為活化受體和抑制受體兩大類。其中活化受體主要包括C型凝集素受體(NKG2D/E/F)、天然細胞毒性受體(NKp46、NKp30和NKp44)和活化殺傷細胞免疫球蛋白樣受體(KIRs-aKIRs);抑制受體主要包括抑制殺傷細胞免疫球蛋白樣受體(KIRS-ikKIRs)和C型凝集素受體(如NKG2A/B)[7-9]。這些受體之間的良好平衡對調節NK細胞功能至關重要,抑制受體通過SHP-1和SHP-2磷酸酶使得激活性受體(DAP10、DAP12)去磷酸化,進而直接抑制受體的激活[10-11]。報道顯示KG1a細胞主要為具有LSCs特性的CD34+/CD38-細胞,LSC的數量超過50%,本課題組先前的研究也表明KG1a細胞具有LSCs的特征,包括分化、增殖和自我更新[12]。NKG2DLs(MICA/B、ULBP2、ULBP3)的表達缺失可能是一種普遍的LSC標志物,利用這種功能特性逃避NK細胞免疫監視的能力[13-15]。
本研究結果表明ATO對 KG1a 細胞的增殖抑制作用與時間濃度呈正相關,考慮到臨床用藥的可能性,選擇ATO作用濃度IC30用于后續的實驗。結果發現,聯合低濃度的ATO 與NK細胞能有效抑制 KG1a 細胞的增殖,降低KG1a 細胞的集落形成能力,且效果好于單用ATO組和ATO聯合NKG2D單克隆抗體預處理NK細胞組。這證明ATO可提高NK細胞殺傷KG1a細胞的能力,且與KG1a細胞表面NKG2D配體表達密切相關。我們接下來檢測ATO對 KG1a 細胞作用前后細胞表面NKG2D配體表達的變化,流式細胞檢測發現KG1a細胞表面NKG2D 配體MICA/B、ULBP2、ULBP3的平均熒光強度(MFI)在ATO作用前后未見明顯改變,而ULBP1 表達在ATO處理后明顯增高(P<0.05)。綜上所述,NK細胞對ATO處理后的KG1a細胞具有明顯殺傷作用,聯合ATO降低了 KG1a 細胞集落形成能力。這與低濃度的ATO誘導KG1a細胞表面NKG2D配體ULBP1的表達密切相關,深層次的機制仍需進一步研究探討。
