高氮磷比與酸化共同作用對微小亞歷山大藻生長和產毒的影響
2021-03-11 05:43:54劉麗君陳軍輝何秀平連子如
海洋科學 2021年2期
郝 爽 , 劉麗君 陳軍輝, 何秀平, 連子如
(1. 山東大學 海洋學院, 山東 威海 264209; 2. 自然資源部第一海洋研究所, 自然資源部海洋生態(tài)環(huán)境科學與技術重點實驗室, 山東 青島 266061)
近年來, 人類活動導致大氣中CO2含量迅速上升,從工業(yè)革命開始時的280 ppm 增至當今的411 ppm[1]。大氣中CO2含量上升不僅會造成氣候變暖, 還會導致海洋中溶解CO2含量增加和海水pH 值降低。目前,表層海水的平均pH 值約為8.10[2], 預計到2100 年將下降至7.8 或7.7, 到2300 年將下降至7.4~7.6, 將對海洋生態(tài)系統(tǒng)造成嚴重影響[3]。富營養(yǎng)化是一種由于水體中氮和磷含量、相對比例或化學形態(tài)發(fā)生變化而導致水體中生產力, 尤其是有機質不斷增加的過程[4], 是水質退化的主要原因之一, 對世界上許多水生態(tài)系統(tǒng)造成重大破壞[5], 還會對人類健康和社會經濟發(fā)展構成威脅[6]。藻類作為最重要的初級生產者之一, 是海洋食物網的基礎, 海水酸化和富營養(yǎng)化引起的海水環(huán)境變化勢必也會影響到海洋藻類的生理機能。
隨著全球氣候問題的加劇, 海洋產毒藻在全球海域的分布范圍越來越大[7-8], 尤其是麻痹性貝毒(PST)毒素產毒藻[9-10]。海洋環(huán)境中能夠產生PST 的微藻主要有微小亞歷山大藻、鏈狀亞歷山大藻[11-12]、鏈狀裸甲藻[13]和巴哈馬梨甲藻[14]等, 這些產毒甲藻在全球近岸海域廣泛分布, 嚴重威脅近海養(yǎng)殖環(huán)境安全。其中, 文獻報道我國多起PST 中毒事件是由亞歷山大藻屬和裸甲藻屬的部分有毒藻種造成的[15-16]。
氮、磷作為產毒藻進行生理活動必需的營養(yǎng)元素[3], 在富營養(yǎng)化的產生和赤潮的暴發(fā)中起著重要的驅動作用。有研究表明, 亞歷山大藻細胞的毒性可能同時受到N 和P 濃度的影響, 并且隨著N/P 的變化而變化[17]。從PST 的化學結構式可以看出, N 元素含量約占PST 分子的30%, N 元素與亞歷山大藻產毒有著十分密切的聯(lián)系。關于酸化研究, 在室內培養(yǎng)實驗中, 最初用化學方法調控培養(yǎng)液的pH 值, 使用強酸模擬酸化環(huán)境, 探究pH 變化對水生生物的影響。如用鹽酸調節(jié)pH 對云紋石斑魚胚胎發(fā)育和仔魚活力以及文蛤的呼吸與排泄進行研究[18-19], 王越等[20]通過用鹽酸探究了pH 對米氏凱倫藻生理代謝的影響。然而, 有毒甲藻在強酸調節(jié)的酸化海水中產毒響應的相關研究目前鮮有報道。目前在產毒藻對環(huán)境因子的響應方面的研究多是針對溫度、鹽度、海水酸化或營養(yǎng)條件等單因素, 而兩個及兩個以上因素的聯(lián)合作用對有毒甲藻的影響研究較少, 關于所釋放的胞外毒素的濃度、組成等也沒有明確報道。那么, 在多種環(huán)境因子的聯(lián)合作用下有毒甲藻產毒的規(guī)律是怎樣, 胞內外毒素組成、含量是否有明顯差異, 這些問題值得深入研究探討。
基于以上論述, 本工作選取我國近海典型產毒甲藻微小亞歷山大藻(A. minutum)作為研究對象, 通過強酸酸化方法, 聯(lián)合一定條件下的富營養(yǎng)化模式,構建基于氮濃度變化的不同氮磷比條件下的酸化海水環(huán)境, 分析測定了胞內外毒素的含量組分以及藻密度、葉綠素含量等相關數(shù)據(jù), 初步探討了在較高氮磷比條件下的酸化海水對A. minutum產毒的影響。
1 材料與方法
1.1 主要儀器與材料
G1969A 型飛行時間質譜儀和1200 型高效液相色譜儀(美國Agilent 公司); ZIC-HILIC 色譜柱(德國MERCK 公司); GTX1、GTX2、GTX3、GTX4 標準品(NRC-IMB), 用0.1 mol/L 乙酸溶液稀釋后濃度分別為497 μg/L、903 μg/L、343 μg/L、162 μg/L; 產毒微小亞歷山大藻分離自臺灣海峽, 株號AM-1。
1.2 藻的培養(yǎng)與收集
取天然海水, 經0.22 μm 濾膜過濾后高溫高壓滅菌, 冷卻至室溫。以NaNO3作為氮源, NaH2PO4作為磷源, 微量元素和維生素的初始濃度按照f/2 培養(yǎng)基配方添加, NaH2PO4初始濃度為9.7±0.3 μmol/L, 通過改變NaNO3濃度改變培養(yǎng)液的氮磷比。向500 mL的錐形瓶中添加300 mL 培養(yǎng)液,A. minutum初始密度為1×103個/mL, 設置明暗比(L︰D)為12 h︰12 h、培養(yǎng)溫度20℃、光照強度3 000 lx, 培養(yǎng)過程中定時搖動錐形瓶。每天用1 mol/L 的鹽酸和1 mol/L 的NaOH 溶液調控培養(yǎng)液的pH 值。實驗設置兩個pH值(8.0, 7.5)和兩個氮磷比(80︰1, 40︰1)共4 個組合,如表1 所示。
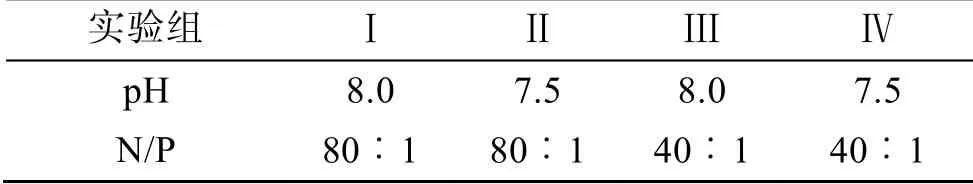
表1 實驗組的設置Tab. 1 Setting of the experimental group
培養(yǎng)周期為34 天, 每隔3 天收集3 瓶藻液。從每個錐形瓶的300 mL 藻液中各取1 mL 混勻的培養(yǎng)液, 置于2 mL 離心管中, 加入1 滴Lugol’s 碘液, 用于藻細胞的計數(shù); 各取2 mL 混勻藻液抽濾至直徑25 mm 的GF/F 膜上, 用于葉綠素a的測定; 將剩余藻液中藻細胞收集至GF/A 膜上, 實現(xiàn)藻細胞與培養(yǎng)液的分離; 用孔徑為0.22 μm 的混合纖維濾膜對培養(yǎng)液進一步過濾。
1.3 測定方法
藻密度、葉綠素a及營養(yǎng)鹽(NO3-N、NO2-N、NH4-N 和PO4-P)的測定以及A. minutum胞內胞外PST的提取、富集和儀器檢測方法參照文獻[21-22]。溶解無機氮(DIN)為NO3-N、NO2-N、NH4-N 濃度之和。
1.4 數(shù)據(jù)分析
數(shù)據(jù)采用 SPSS 26.0 的One-way ANOVA 和LSD (Least Significant Difference, 最小顯著性差異法)多重比較分析,P<0.05 為差異顯著,P<0.01 為差異極顯著; 使用 Origin 2018 軟件繪圖。
2 結果
2.1 對生長的影響
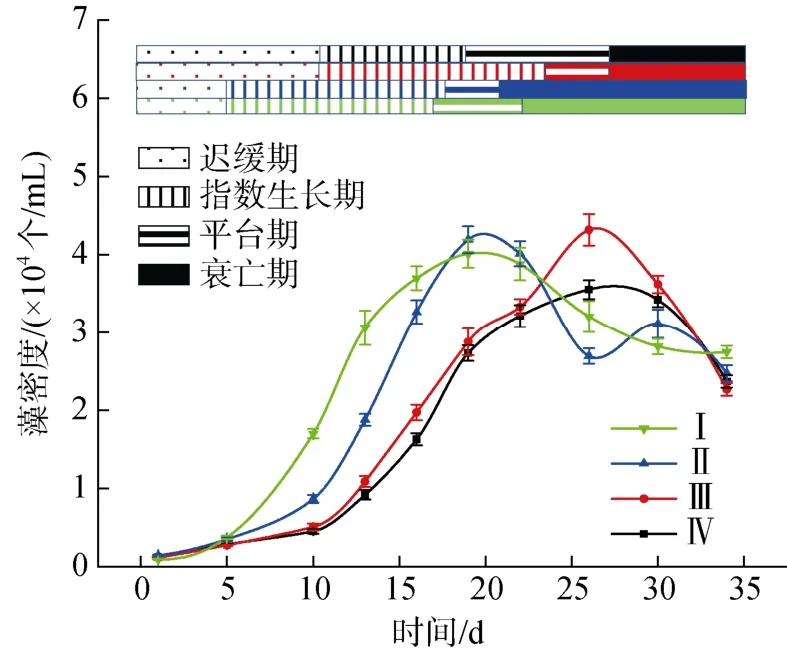
圖1 不同氮磷比條件下酸化環(huán)境對微小亞歷山大藻密度的影響Fig. 1 Effect of acidification on the density of A. minutum under different N/P ratios
不同氮磷比條件下培養(yǎng)液酸化對A. minutum生長狀況的影響如圖1 所示。各實驗組A. minutum生長可分為四個階段: 遲緩期、指數(shù)生長期、平臺期和衰亡期。N/P=80︰1 時,A. minutum進入指數(shù)生長期和平臺期的時間比N/P=40︰1 條件下提前5 天左右, 適當降低培養(yǎng)液的pH 值可以提高A. minutum的最大細胞數(shù)。N/P=40︰1 時, pH=8.0 條件下A. minutum的最大細胞數(shù)反而顯著高于pH=7.5 條件。
2.2 對葉綠素a 的影響
如圖2a 所示,A. minutum葉綠素a含量在培養(yǎng)周期中先增加后減少, 與圖1 中微小亞歷山大藻密度變化趨勢基本一致。每個細胞內葉綠素a含量(pg/細胞)的變化情況與單位體積含量(μg/L)在不同實驗條件下的相對關系基本一致, 除實驗組Ⅳ中每個細胞內葉綠素a含量隨生長周期不斷減少之外, 其余三組的細胞葉綠素a含量在整個生長周期中變化不大(見圖2b)。從表2 可以看出微小亞歷山大藻密度與葉綠素a含量成正相關關系, 氮磷比的改變和酸化環(huán)境沒有改變這種關系。在A. minutum的培養(yǎng)周期中, 葉綠素a含量隨培養(yǎng)液pH 值的變化而發(fā)生改變:生長初期, 葉綠素a含量隨培養(yǎng)液pH 值的降低而降低; 進入指數(shù)生長期后, 培養(yǎng)液pH 降低時A. minutum葉綠素a含量增加極為顯著 (P<0.01)。
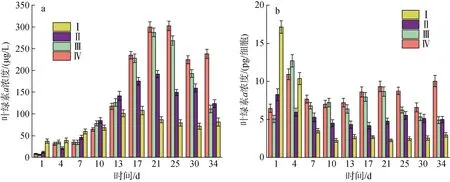
圖2 不同氮磷比條件下的酸化環(huán)境對微小亞歷山大藻葉綠素a 含量的影響Fig. 2 Effect of acidification on the chlorophyll a content of A. minutum under different N/P ratios
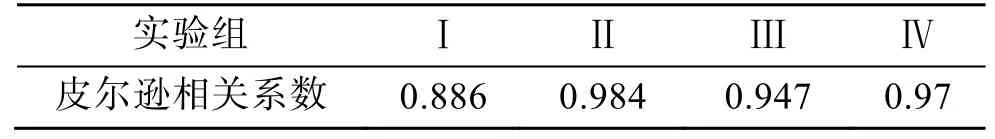
表2 葉綠素a 與藻密度的相關性Tab. 2 Correlation between chlorophyll a and algal density
2.3 對營養(yǎng)鹽(DIN 和PO4-P)的測定
培養(yǎng)液中DIN 和PO4-P 濃度變化趨勢不同(圖3)。不同實驗條件下的微小亞歷山大藻在前五天快速吸收PO4-P, 四組實驗分別吸收培養(yǎng)液中PO4-P 的64.03%(Ⅰ)、52.19%(Ⅱ) 、73.65%(Ⅲ)和62.58%(Ⅳ)。之后培養(yǎng)液中PO4-P 的含量沒有明顯變化。N/P=80︰1實驗組培養(yǎng)周期內DIN 的濃度大致呈現(xiàn)先減少后增加的趨勢, 而N/P=40︰1 的兩組培養(yǎng)液中DIN 的濃度則隨培養(yǎng)時間而減少。其中NO3-N 作為培養(yǎng)基中添加的唯一氮源, 與DIN 的變化趨勢基本一致; NH4-N濃度則隨著A. minutum的生長呈現(xiàn)波動上升的趨勢;NO2-N 占DIN 的比例不足1%, 且對A. minutum的生長影響不大, 在此不予討論。
2.4 對毒素的影響
為了更直觀地比較不同氮磷比條件下的酸化培養(yǎng)液對A. minutum生長和產毒的影響, 根據(jù)4 組實驗的藻密度曲線選取五個點進行分析: 遲緩期(樣品1),指數(shù)前期(樣品2), 指數(shù)后期(樣品3), 最大生物量所在的樣品(樣品4), 衰亡期(樣品5)。
2.4.1 對A. minutum 胞內毒素的影響
不同氮磷比條件下酸化培養(yǎng)液對A. minutum單位細胞毒素總量的影響見圖4。N/P=40︰1 時, pH 的改變對指數(shù)期至衰亡期A. minutum單位細胞總毒素含量的影響并不明顯(P>0.05); 而當?shù)妆容^高(N/P=80︰1)時, 與A. minutum藻細胞生長趨勢不同,pH 的降低反而顯著降低了A. minutum單位細胞總毒素含量(P<0.01)。相同培養(yǎng)液pH 條件下, N/P=80︰1條件下的A. minutum細胞在衰亡期積累的毒素迅速增加, 含量比N/P=40︰1 條件下高出57.8%(pH=8.0)和42.5%(pH=7.5)。
在不同生長時期A. minutum胞內4 種PST 總量及毒素組成如圖5 所示, 可以看出胞內PST 總量隨培養(yǎng)時間基本呈現(xiàn)增加的趨勢, 尤其在N/P=80︰1 時,毒素含量增加顯著。胞內GTX1 和GTX4 隨培養(yǎng)時間基本呈現(xiàn)增高的趨勢, GTX2 和GTX3 含量變化不大甚至減少。N/P=80︰1 時, 4 種PST 在遲緩期的含量隨pH 降低而增加, 而在指數(shù)期至衰亡期, 各毒素反而隨pH 的降低而降低。N/P=40: 1 條件下, 在pH 降低時各毒素含量除遲緩期外增加了5.04% ~ 62.38%。
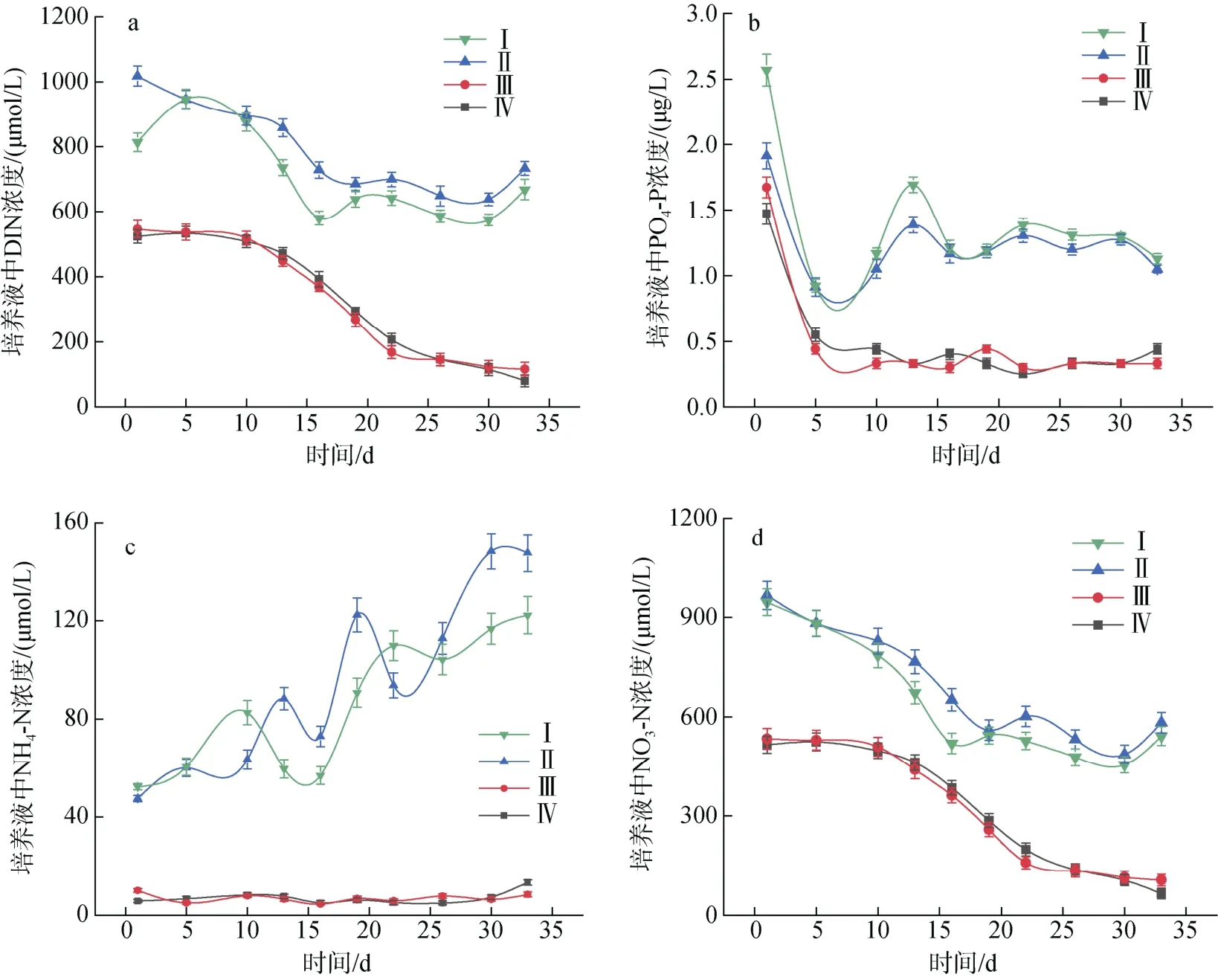
圖3 不同氮磷比條件下酸化環(huán)境對微小亞歷山大藻培養(yǎng)液中營養(yǎng)鹽含量的影響Fig. 3 Effects of acidification on nutrient content in A. minutum culture medium under different N/P ratios
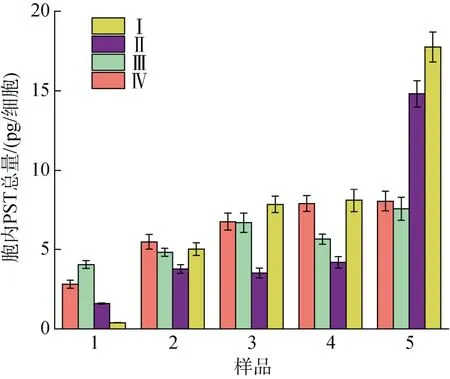
圖4 不同氮磷比條件下酸化環(huán)境對微小亞歷山大藻單位細胞毒素總量的影響Fig. 4 Effect of acidification on the total amount of PST of an A. minutum cell under different N/P ratios
2.4.2 對A. minutum 胞外毒素的影響
不同氮磷比條件下酸化培養(yǎng)液對A. minutum胞外培養(yǎng)液中PST 總量的影響如圖6 所示。相同pH條件下, N/P=80︰1 時衰亡期之前胞外培養(yǎng)液的PST含量明顯高于 N/P=40︰1 條件, 而到衰亡期時,N/P=40︰1 條件下培養(yǎng)液中PST 迅速增加, 40/7.5 實驗組中毒素含量超過80/7.5 實驗組。不同氮磷比條件下pH 對衰亡期之前的A. minutum胞外培養(yǎng)液中PST 總量無明顯影響, N/P=80︰1 條件下的A. minu-tum到達衰亡期時, pH=8.0 條件下的胞外PST 含量迅速增加, 超過了較低pH 條件下毒素含量71.05%。
在不同生長時期A. minutum胞外培養(yǎng)液中PST含量以及毒素組成如圖7 所示(圖中數(shù)據(jù)為三組平行實驗結果的平均值), 胞外培養(yǎng)液中PST 總量隨培養(yǎng)時間均呈現(xiàn)增加的趨勢。氮磷比較高(N/P=80︰1)時,pH 降低并未顯著促進PST 總量的增加(P>0.05), 但在衰亡期之前胞外培養(yǎng)液中GTX2 和GTX3 占比隨pH 降低而提高, GTX1 和GTX4 與之相反, 在pH=7.5時其濃度較低; N/P=40︰1 時, pH 變化對胞外培養(yǎng)液中PST 的含量及組成無明顯影響(P>0.05)。
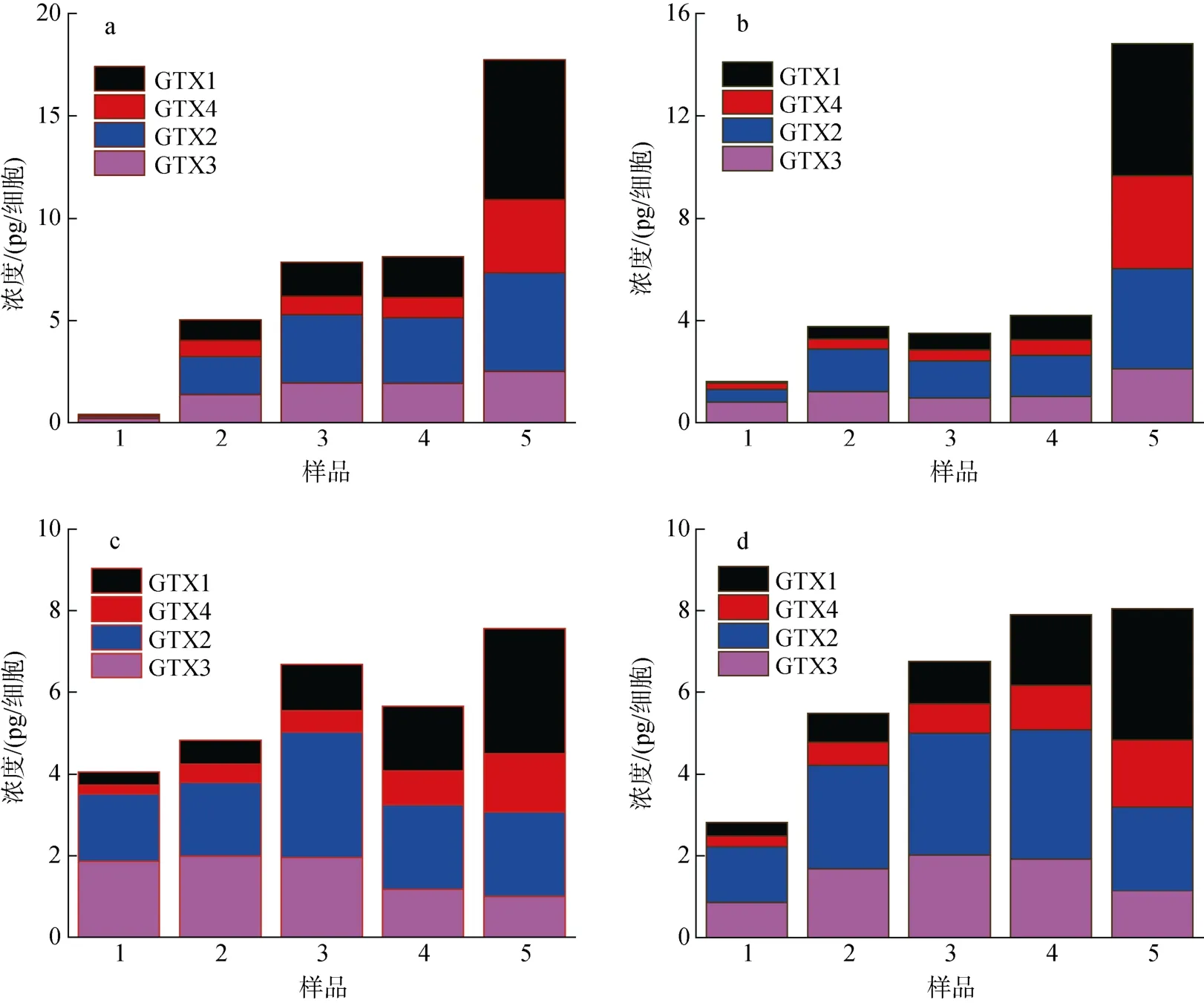
圖5 4 組實驗微小亞歷山大藻胞內PST 總量的變化Fig. 5 Changes in the total amount of intracellular PST in A. minutum in four experimental groups
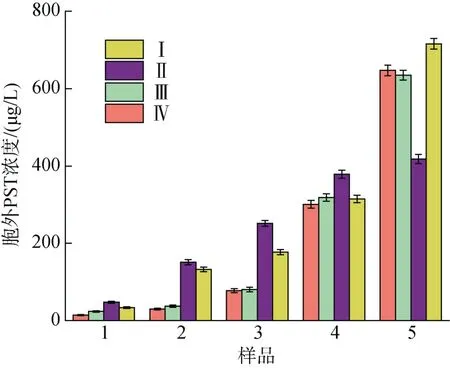
圖6 不同氮磷比條件下酸化環(huán)境對微小亞歷山大藻胞外培養(yǎng)液中總毒素的影響Fig. 6 Effects of acidification on the total amount of extracellular PST in A. minutum under different N/P ratios
2.4.3 對A. minutum胞外毒素與胞內毒素比值的影響
本實驗將A. minutum胞外培養(yǎng)液中4 種PST 與胞內進行比較, 得到單位體積中胞外培養(yǎng)液和胞內4 種PST 物質的量(nmol)以及毒素總量(nmol)的平均值的比值(圖8、圖9)。PST 總量的比值以及GTX1、GTX4、GTX2、GTX3 各自比值的變化規(guī)律相似: 衰亡期之前,N/P=80︰1 條件下二者比值高于N/P=40︰1 條件下的比值, 在遲緩期尤為明顯(P<0.01), 且遲緩期時隨著pH 的降低二者比值減小59.2%。指數(shù)生長期胞外與胞內毒素的比值隨著pH 的降低而增加。衰亡期時,N/P=40︰1 時的胞外/胞內毒素比值高出N/P=80︰1條件下比值的52.4%。所以, 較高氮磷比條件下, 特別是在遲緩期和指數(shù)期, 酸化環(huán)境對A. minutum胞外培養(yǎng)液中4 種PST 的含量與胞內的比值會產生影響(P<0.05)。
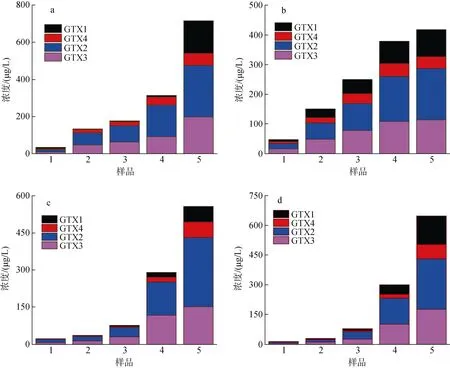
圖7 4 組實驗微小亞歷山大藻胞外培養(yǎng)液中PST 總量的變化Fig. 7 Changes in the total amount of extracellular PST of A. minutum in four experimental groups
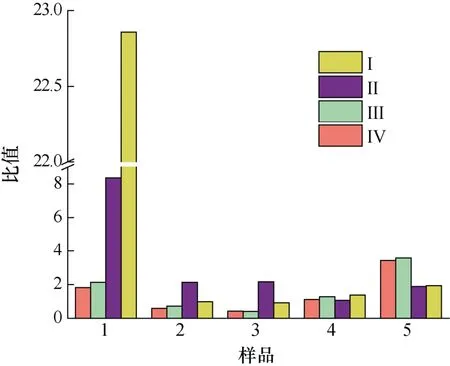
圖8 不同氮磷比條件下酸化環(huán)境對微小亞歷山大藻胞外毒素與胞內毒素總量比值的影響Fig.8 Effects of acidification on the ratio of total extracellular PST to total intracellular PST in A. minutum under different N/P ratios
3 討論
現(xiàn)有的大多數(shù)關于酸化環(huán)境對亞歷山大藻生長及產毒影響的研究是通過CO2加富進行酸化實驗。有研究顯示, 由于碳濃縮機制的存在, CO2濃度的在一定范圍內升高并不會對亞歷山大藻的生長產生明顯影響, 但會使與次級代謝產物生物合成、運輸和分解代謝以及氨基酸運輸相關的下調基因數(shù)量增加,從而降低塔瑪亞歷山大藻株毒素含量[23]。與通過CO2加富模擬酸化環(huán)境不同, 本實驗采用鹽酸來調節(jié)培養(yǎng)液的pH, 探究高氮磷比條件下酸化環(huán)境對A.minutum生長和產毒的影響。
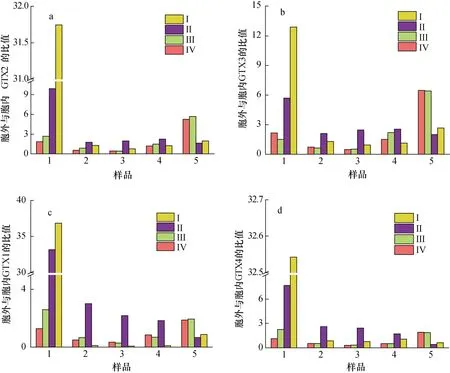
圖9 不同氮磷比條件下酸化環(huán)境對微小亞歷山大藻胞外與胞內不同毒素的比值的影響Fig. 9 Effects of acidification on the ratios of different extracellular to intracellular toxins in A. minutum under different N/P ratios
3.1 對A. minutum 生長的影響
雖然氮是蛋白質和核酸等生命物質的重要組成元素, 但是在本實驗中, 氮磷比較高(N/P=80︰1)時并沒有對產毒微小亞歷山大藻的生長產生促進作用,可能是過量氮在藻體內蓄積轉變成有害物質, 從而對藻的生長造成不利影響。但pH 值降低會促進有毒甲藻的生長, 增長速度快且最大生物量高, 進而引發(fā)危害程度更高的有毒赤潮。
A. minutum產生的PST 是一種富含氮的生物堿[17],而磷元素是A. minutum遺傳物質核酸的重要組成成分之一[24],A. minutum細胞內PST 含量與培養(yǎng)液營養(yǎng)狀況密切相關。本研究顯示A. minutum在培養(yǎng)的前五天快速吸收培養(yǎng)液中的PO4-P, 對P 采取儲存策略;在培養(yǎng)周期的中后段,A. minutum幾乎不再從培養(yǎng)液中吸收P; 而A. minutum在整個生長過程中持續(xù)消耗氮, 對氮營養(yǎng)鹽沒有明顯的儲存策略。當細胞死亡破裂時, 胞內未吸收利用的NO3-N 可能會重新釋放到培養(yǎng)液中, 導致培養(yǎng)后期培養(yǎng)液中DIN 升高。N 在生物體內會發(fā)生轉化, 部分NO3-N 可能在A. minutum中轉化成NH4-N, 并隨著的藻細胞生長周期被利用或釋放。酸化環(huán)境下, 氮磷比的升高可能會降低A.minutum的磷儲存能力而對藻的氮吸收能力沒有明顯的影響。
3.2 對A. minutum 產毒的影響
氮磷比增加有利于A. minutum胞內GTX1 和GTX4 的占比的增加, 使A. minutum產生的PST 向毒性更高的化合物轉變, 其中衰亡期毒素含量更高。Mohamed 的觀點可以解釋這一現(xiàn)象: 亞歷山大藻進入衰亡期后, 細胞分裂間期延長, 細胞分裂的速度減緩甚至停滯, 而PST 合成酶的編碼基因仍持續(xù)表達, PST 可持續(xù)合成[25], 使得衰亡期的A. minutum胞內PST 高于其他生長時期。衰亡期的A. minutum胞外培養(yǎng)液中PST 總量也比其他生長時期要高, 可能原因有二: (1) 衰亡期微小亞歷山大藻胞內PST 含量高于其他生長時期, 所以釋放到胞外培養(yǎng)液中的PST 也會更多; (2) 藻細胞進入到衰亡期, 死亡、裂解速度加快,A. minutum胞內的毒素直接進入到培養(yǎng)基質, 導致胞外PST 含量升高。本實驗的結果與Lim 的研究結果基本一致, 認為亞歷山大藻細胞的毒性隨著N/P 的變化而變化, 在不同的N/P 處理下A. minutum的毒素組成相對穩(wěn)定, 在較高的N/P 條件下培養(yǎng)的藻細胞的毒素含量也比較多[17]。此外Han 還發(fā)現(xiàn), 在高氮條件下的PST 種類更加豐富[26], 這點在本實驗中并未體現(xiàn),可能存在實驗藻株之間的差異。
較高氮磷比條件下的酸化環(huán)境使胞外培養(yǎng)液中PST 含量比胞內增加的程度高。在指數(shù)前期之前,A. minutum胞外培養(yǎng)液中PST 的總量較胞內可高出20 倍以上, 這可能是處于酸化環(huán)境中的微小亞歷山大藻為了提高在惡劣環(huán)境下的生存能力而向環(huán)境中釋放更多的毒素來提高自身競爭力; 也可能是酸化環(huán)境會提高藻細胞內相關酶的含量或活性[27], 進而促進藻細胞的生理代謝以及PST 的合成與釋放。綜上所述, 在富營養(yǎng)化條件下, 日益嚴重的酸化情況可能會導致海水中PST 含量與藻胞內的比值顯著增加, 嚴重威脅到海水環(huán)境。
4 結論
微小亞歷山大藻對氮磷比和pH 的變化較為敏感, 在本實驗中, 酸化環(huán)境對A. minutum的生長、毒素含量和毒性有一定促進作用, 可以推測日益酸化的海水環(huán)境或許在一定程度上會加劇A. minutum引起的有毒赤潮。但也有其他研究表明酸化對亞歷山大藻生長和產毒有一定的抑制作用, 可見酸化對亞歷山大藻的影響十分復雜。酸化環(huán)境如何影響亞歷山大藻的生長和產毒?不同酸化模式對亞歷山大藻的作用機制是否一致?尚未有明確統(tǒng)一的定論。酸化環(huán)境下氮磷比增加并沒有促進A. minutum的生長,且降低了其葉綠素a含量和對P 的儲存能力, 但幾乎沒有影響到對硝氮的吸收能力。氮磷比的增加還促進了A. minutum胞內和胞外毒素含量和毒性的增加,這更加增大了A. minutum赤潮產生的環(huán)境危害。本研究通過探究無CO2參與的酸化環(huán)境和氮加富的共同作用對微小亞歷山大藻生長產毒的影響, 以期為酸化和富營養(yǎng)化條件下的微藻產毒提供更多研究基礎。
猜你喜歡
中學生數(shù)理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
媽媽寶寶(2017年3期)2017-02-21 01:22:28
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
中國塑料(2016年3期)2016-06-15 20:30:00
