CoCl2脅迫下青稞苯丙氨酸解氨酶基因的克隆與表達分析
2021-03-20 02:12:28耿貴工謝惠春
中國農業大學學報 2021年3期
關鍵詞:植物
喬 楓 張 麗 耿貴工 陳 志 曾 陽 謝惠春
(1.青海師范大學 青海省青藏高原藥用動植物資源重點實驗室,西寧 810008;2.青海大學 農林科學院,西寧 810016)
青稞(Hordeumvulgarevar nudum)是青藏高原地區種植的一種重要農作物,可以作為西藏自治區和青海省農牧民的主食。與其他植物相比青稞抗逆能力較強,如抗旱、抗鹽堿、抗冷、抗寒和抗病等[1-3]。青稞基因組中蘊藏著大量的抗逆基因,其獨特的遺傳基礎、生理功能和次生代謝產物逐漸成為學者研究極端逆境的首選植物或農作物材料[1-3]。青稞的抗逆作用是受到多方面調控的,在逆境下可以通過生理代謝調節,如滲透調節、生長發育調節,來抵抗不良環境。也可以通過基因表達調節,如通過轉錄因子、抗逆基因的誘導表達對逆境作出響應。前人已從植物生理生化和分子生物學方面探討青稞抗逆境機制[4-6]。本研究探索在鈷脅迫下青稞幼苗相關抗逆基因的表達變化,為青稞的抗重金屬機制提供線索和科學依據。
鈷易被植物根系吸收,能夠在植物的不同組織中如根、莖中積累,影響植物的生長發育,超過一定濃度后,鈷會抑制植物的生長甚至引起植物死亡[7]。我國大部分地區的土壤鈷含量為5~40 mg/kg,鈷含量平均背景值為11.6 mg/kg[8]。當植物體內的Co2+過高時,嚴重損傷植物細胞的膜結構和代謝酶如超氧化物歧化酶(SOD)、過氧化物酶(POD)、谷胱甘肽合成酶(GSH)等活性,植物會因毒害而死亡[9-10]。小麥遭受Co2+脅迫后,施用生物炭可調節細胞內抗氧化酶活性和抗壞血酸-谷胱甘肽循環,可以最大限度地減少Co2+誘導的氧化損傷[11]。通過轉錄組學及microRNA測序研究鈷脅迫下垂柳的基因表達和生理生化指標的變化,結果發現, 100 μmol/L 的鈷脅迫植株的葉片枯黃、根系生長受阻、POD和SOD活性升高,過氧化氫酶(CAT)活性降低;轉錄組序列分析發現鈷脅迫的植物根部有1 165 個差異基因和62個差異表達microRNA,地上部分組織中有836個差異基因和80個差異表達microRNA[12]。植物如何降低或減弱Co2+的毒害作用,是農作物抗逆生理研究的熱點之一。目前,青稞幼苗響應鈷脅迫分子機制的研究尚未見報道。本研究設置不同濃度Co2+處理,分析青稞‘昆侖15號’幼苗中抗逆相關的過氧化物酶(POD)、苯丙氨酸解氨酶(PAL)、谷胱甘肽合成酶(GSH)、Δ1-吡咯啉-5-羧酸合成酶(P5CS)和超氧化物歧化酶(SOD)的基因表達差異,旨在闡明青稞植株抗氧化酶基因對Co2+脅迫的生理響應機制,以期為青稞的抗Co2+栽培和育種提供參考。
1 材料與方法
1.1 材料
供試青稞品種為‘昆侖15號’,由青海大學農林科學院作物所提供。
挑選無病蟲害的種子,先用水沖洗3次,再用2%的次氯酸鈉消毒3 min,后用去離子水洗2~3次。在直徑為20 cm的培養皿中鋪放2層濾紙,將滅過菌的青稞種子均勻地分散在培養皿的濾紙上,每個培養皿放50粒種子,置于光照培養箱中,溫度為28 ℃,每天定時定量用去離子水處理,保持一定的濕度。持續處理8 d,待青稞幼苗長出2片子葉,選取長勢一致的幼苗用于氯化鈷溶液處理。
將氯化鈷(分析純)配置成30、70、100、150、200和250 mg/L的6個Co2+處理濃度,用6種處理液對青稞幼苗繼續處理培養,對照用去離子水處理,每天定時定量處理。持續處理7 d,取樣用于分析幼苗相關逆境基因的表達變化。
1.2 青稞總RNA的提取及cDNA的合成
青稞幼苗葉片總RNA的提取選用Tiangen生化公司(北京)公司的植物總RNA提取試劑盒,cDNA的第一鏈的合成選用TaKaRa公司的PrimeScriptTM1st strand cDNA synthesis,瓊脂糖PCR產物的測序選用TaKaRa公司的瓊脂糖凝膠回收試劑盒,PCR產物連接到克隆載體pGEM-T easy(美國Promega公司) 及測序、引物合成由上海桑尼生物科技有限公司完成。在線Primer 3.0 軟件設計引物,引物序列,見表1。
1.3 序列的生物信息學分析
采用 DNAstar 軟件拼接基因測序結果,得到青稞PAL基因的全長cDNA序列。序列Blast相似性比對在NCBI(http:∥www.ncbi.nlm.nih.gov/)網站上分析,ORF 的查找和核苷酸的翻譯在軟件 ORF Finder (http:∥www.ncbi.nlm.nih.gov/gorf/gorf.html) 上進行,PAL基因編碼的蛋白質的結構特點和結構性質等在ExPASy(http:∥www.expasy.org)上分析,多種植物PAL的氨基酸序列的多重比對由Mega 3.1軟件Clustal X (1.83)完成,系統發育樹采用最小進化法(Minimum Evolution)構建。

表1 引物序列Table 1 Primer sequences
1.4 引物設計
選用28SrRNA作為參比基因,同時設置滅菌超純水為陰性對照。根據NCBI網站里大麥(Hordeumvulgare)的超氧化物歧化酶基因(SOD)、Δ1-吡咯啉-5-羧酸合成酶基因(P5CS)、苯丙氨酸解氨酶基因(PAL)、過氧化物酶基因(POD)、谷胱甘肽合成酶基因(GSH)、28SrRNA基因序列,設計引物在Primer 3.0完成,合成引物由上海桑尼生物科技有限公司完成,引物序列見表1。
1.5 實時熒光定量PCR技術
實時熒光定量 PCR 方法采用 SYBR?Premix Ex TaqTM(TaKaRa 公司)試劑盒。PCR 擴增程序為:95 ℃ 預變性 40 s;95 ℃ 變性 10 s,62~64 ℃延伸 30 s,40 個循環,熒光定量 PCR 反應結束后確認反應的融解曲線、擴增曲線和產物定量。基因的相對表達量通過2-ΔΔCt方法計算[13]。
1.6 統計分析
數據的統計學分析采用Excel 2016處理,在α=0.05水平上分析差異顯著性,作圖選用Sigmal Plot 14.0軟件完成。
2 結果與分析
2.1 青稞PAL基因的克隆
采用電子同源克隆法,以青稞幼苗葉片RNA為模板,使用不同引物組合擴增青稞PAL基因序列,并且在NCBI網站比對分析,結果顯示青稞測序結果與大麥PAL基因序列有較高的相似性。其中經PALF1/PALR1引物組合擴增、測序青稞PAL基因片段長度為1 200 bp,經PALF2/PALR2擴增的產物測序為900 bp,經PALF2/PALR3擴增的產物測序為830 bp,與預期結果一致。將獲得的3個片段用DNAstar軟件拼接成一個連續的序列。在此序列上設計引物組合PALF3/PALR4擴增全長序列后,經測序獲得 2 142 bp 的序列(圖1)。
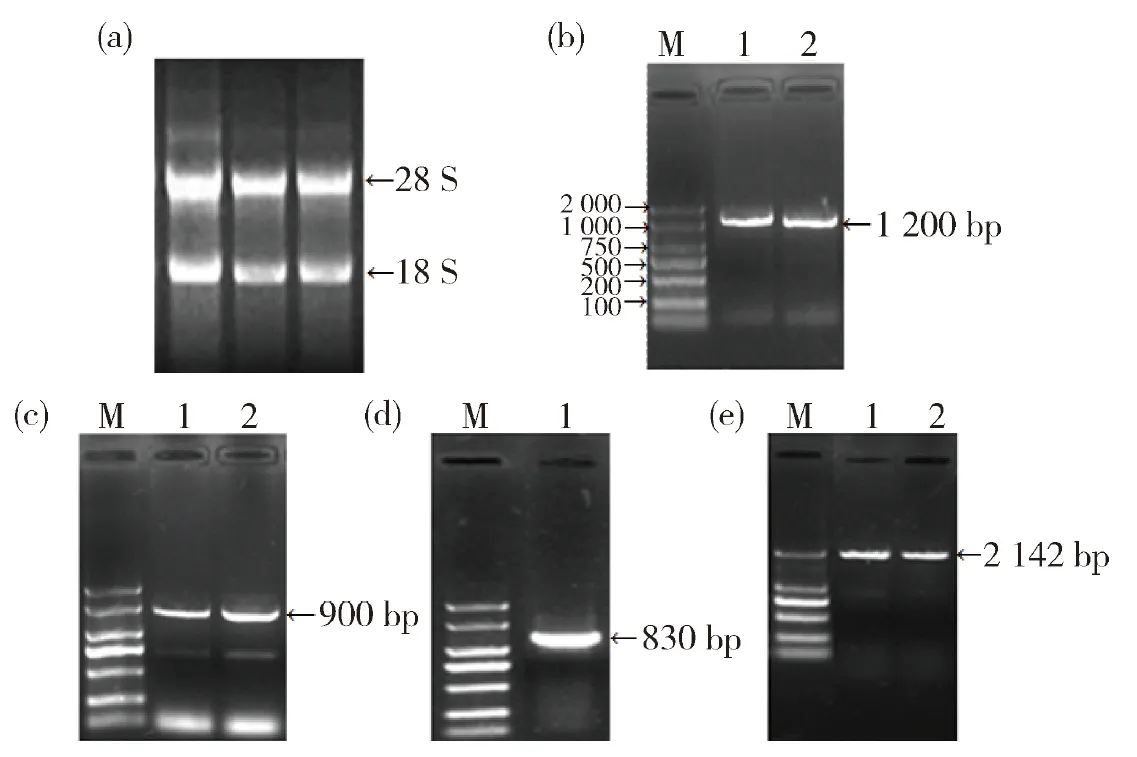
(a)RNA提取結果;(b)PALF1/PALR1結果; (c)PALF2/PALR2結果; (d)PALF2/PALR3結果;(e)PALF3/PALR4結果. M,DL 2 000 Marker
2.2 青稞PAL基因生物信息學分析
青稞PAL基因cDNA序列為2 142 bp,其堿基序列相對應的編碼氨基酸有713個。將青稞PAL的cDNA序列,命名為HvPAL基因(Genbank No. MK695675)。將青稞PAL氨基酸序列在NCBI網站上預測其功能結構域,表明HvPAL的第195~211(GTITASGDLVPLSYIA)的氨基酸是苯丙氨酸/組氨酸解氨酶的保守結構域,位于苯丙氨酸/組氨酸解氨酶的活性中心,見圖2。青稞PAL氨基酸序列同時有保守的催化激活位點(第433位-N、第434位-G、第476位-H和第483~487位的HNQDV序列),脫氨基位點(第426位-L、第427位-V和第469位-L),這些保守的激活位點與脫氨基位點與擬南芥和煙草的PAL一致。經POSORT和Motif Scan軟件分析,青稞PAL氨基酸的N-端無蛋白信號肽定位序列,推測是可溶性蛋白,在C-端序列有2個雙亮氨酸基元(LL),分別在第563~564位和第698~699位。

黃底方框中為苯丙氨酸/組氨酸解氨酶的保守結構域
2.3 青稞PAL編碼蛋白與其他植物同源性比較
由圖3可知,青稞與大麥、小麥和水稻等的PAL氨基酸序列相似性為90%。
對青稞PAL氨基酸序列進行三級結構預測顯示,其三級空間結構中α-螺旋占93.25%;無規則卷曲和β-轉角含量較少,見圖4。
2.4 PAL氨基酸構建的分子系統進化樹
由圖5可知,本研究中的青稞PAL氨基酸序列聚在禾本科分支,與植物傳統的分類結果一致。
2.5 青稞基因的表達
2.5.1青稞抗逆相關基因的表達變化
由圖6(a) 可知,青稞幼苗隨Co2+濃度增加其POD基因的表達升高,與對照相比,在200和250 mg/L Co2+處理下,POD基因表達分別是對照的15.1和13.2倍(P<0.05),在 200 mg/L Co2+濃度時POD基因表達量最大,均達到顯著水平;其他Co2+處理POD基因表達增加但沒有達到顯著水平。
30~150 mg/L Co2+處理下,青稞PAL基因表達呈逐漸增加趨勢(圖6(b)),在150 mg/L Co2+濃度時PAL基因表達是對照的42倍(P<0.05),其他濃度下,PAL基因的表達量較對照沒有顯著差異,見圖6(b)。
30~150 mg/L Co2+濃度下(圖6(c)),青稞幼苗中GSH基因表達呈逐漸增加趨勢;100~150 mg/L Co2+處理濃度時,GSH基因表達量較對照顯著增加(P<0.05);150 mg/L的Co2+濃度下基因表達為最高水平。其余濃度處理下,GSH基因相對表達量未達到顯著水平。
青稞葉片P5CS基因表達量隨著30~200 mg/L Co2+濃度增加呈逐漸升高趨勢(圖6(d)),其中100~200 mg/L Co2+濃度下P5CS基因表達是對照的13~27倍,達到顯著水平(P<0.05),150 mg/L Co2+濃度下達到最大值;其余濃度處理下,P5CS基因相對表達量未達到顯著水平。
在30~150 mg/L的Co2+處理,SOD表達量逐步升高,在70 mg/L Co2+濃度下達到最大值,是對照的9.2倍(P<0.05)。其他Co2+濃度處理下,青稞葉片中SOD表達量與對照差異不顯著,見圖6(e)。
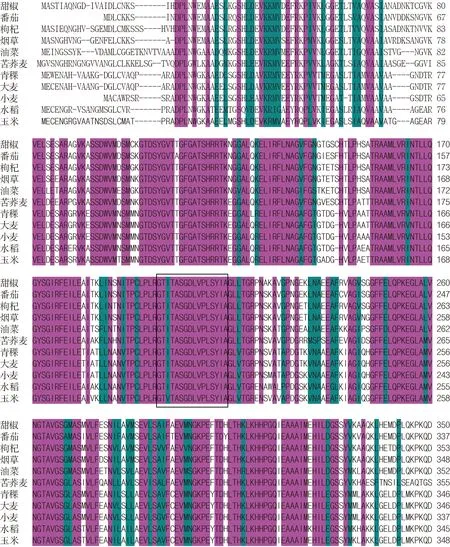
玫紅色代表氨基酸相同, 深綠色代表有2種或以上氨基酸相同。方框內為PAL/HIS的保守結構域。
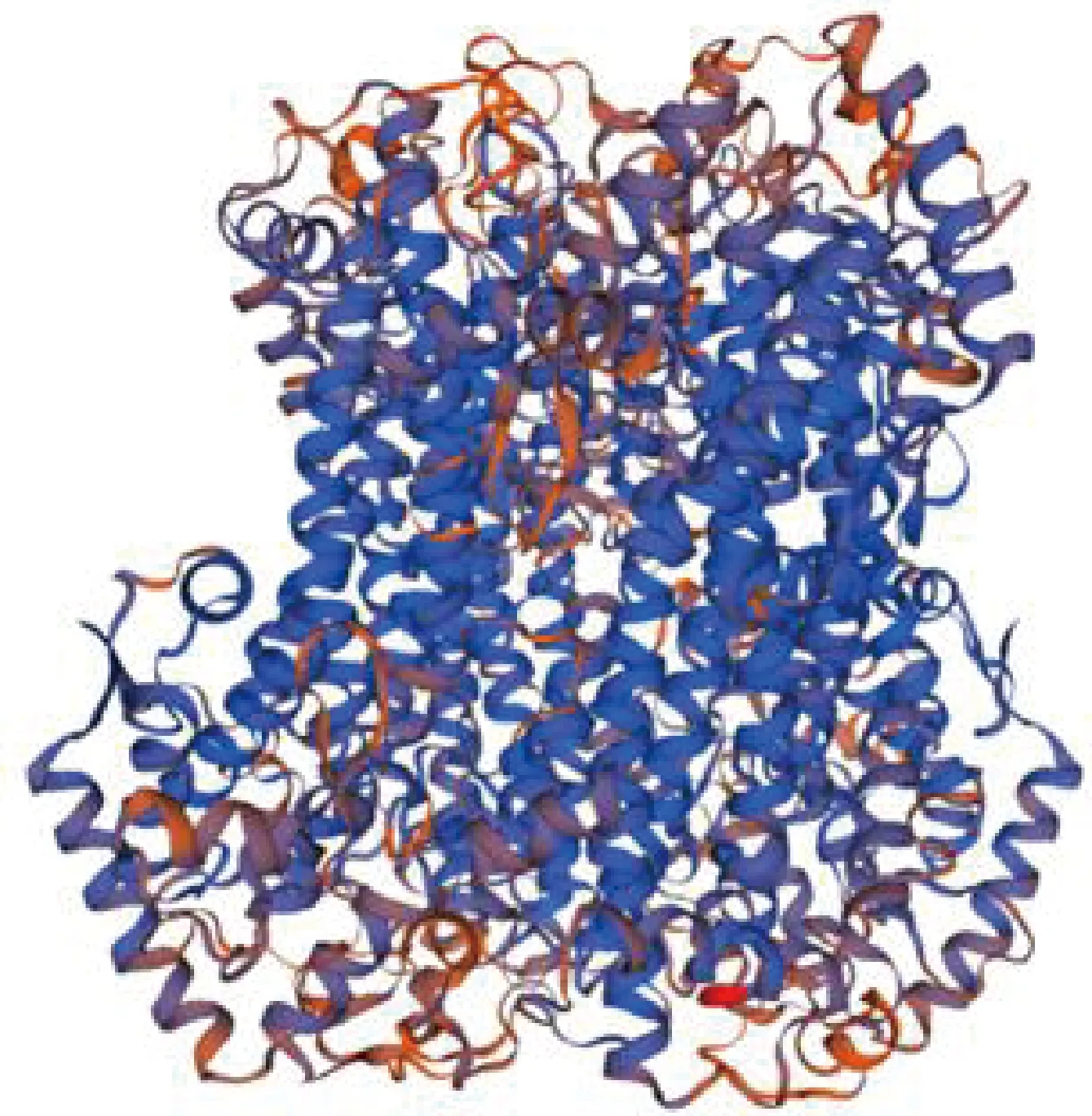
藍色,α-螺旋;紅色,無規則卷曲。Blue,α-helix; Red,Randon coil.
2.5.2基因表達量的相關性
由表2可知,青稞幼苗在不同Co2+濃度處理下,不同基因表達量之間的相關系數(R2)不同。在0.05水平上相關系數R2=0.754,在0.01水平上R2=0.874。PAL與GSH基因表達量呈顯著正相關(R2=0.850)。GSH與P5CS(R2=0.594)、POD與P5CS(R2=0.542)、GSH與SOD(R2=0.616)均呈正相關關系,但未達到顯著水平。由低到高Co2+濃度處理,青稞幼苗SOD、P5CS、PAL、POD、GSH基因表達也依次增強,共同作用以抵御和適應Co2+逆境脅迫。
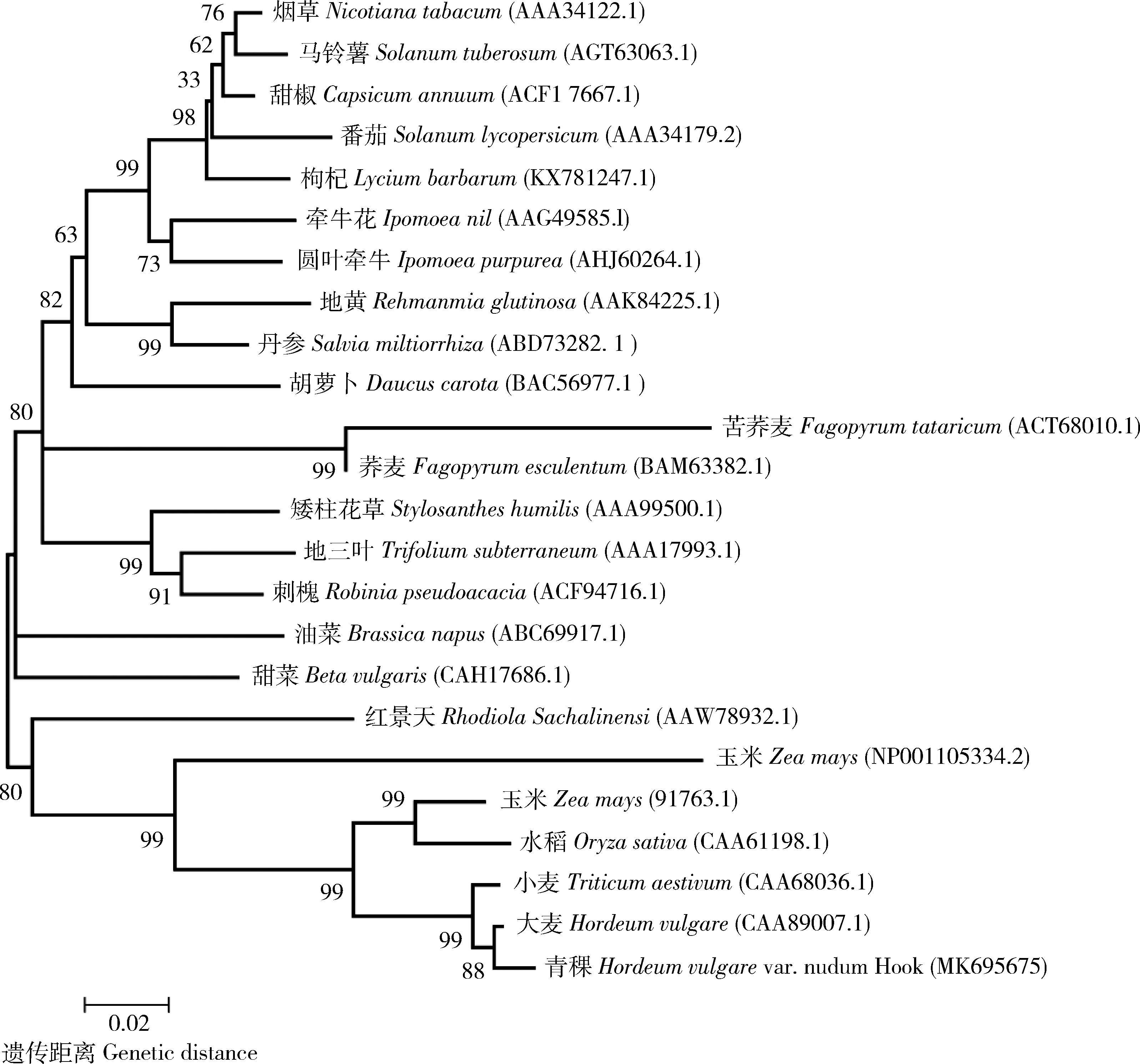
圖5 PAL氨基酸分子系統進化樹
3 討論與結論
植物苯丙氨酸解氨酶(PAL)是苯丙酸合成途徑中的第一個酶,也是其代謝途徑中的關鍵酶之一,植物PAL受干旱、低溫、紫外線和病原物等不同因素的影響而誘導基因表達增加[14-15]。PAL活性增加可使植物抵抗不良環境或病原菌侵染的能力提高。通常可以把植物的PAL活性作為誘導抗性的一個參考指標[16-18]。本研究克隆了青稞HvPAL基因,發現該基因的氨基酸序列含有苯丙氨酸解氨酶、組氨酸裂解酶和苯丙氨酸-組氨酸裂解酶3個保守結構域。青稞PAL與大麥、小麥等的氨基酸序列相似性達90%,氨基酸序列的分子系統進化樹分析表明,青稞與小麥、大麥、水稻和玉米等聚為禾本科分支,這與已有研究結果一致[19]。本研究青稞幼苗在0~150 mg/L Co2+處理下,PAL基因表達逐漸增加,在150 mg/L Co2+處理時達到最大值。綜上,逆境誘導青稞PAL基因的表達來響應和抵抗Co2+的脅迫,這與絲瓜響應逆境的PAL基因表達的變化結果一致[20]。
SOD是一類重要的抗氧化酶,能夠將超氧化物自由基轉化為過氧化氫和分子氧,是抗氧化防御的第一道防線[21]。超氧化物歧化酶基因的表達受到轉錄、轉錄后和翻譯水平上調控,在發育和應答生物或非生物脅迫中起保護植物的作用[22]。在10~40 ℃,3個落葉松超氧化物歧化酶基因(LkSOD2、LkSOD4和LkSOD6)的擬南芥轉基因植株中,在鹽脅迫下均表現出較高的發芽率和較長的根長[22]。LkSOD5基因可恢復擬南芥atfsd2-2突變體的淺綠色和矮化表型[22]。本研究中在30~150 mg/L的Co2+處理,SOD表達量升高,植物通過提高SOD酶的表達量來清除超氧化物自由基,降低活性氧對青稞幼苗的傷害。
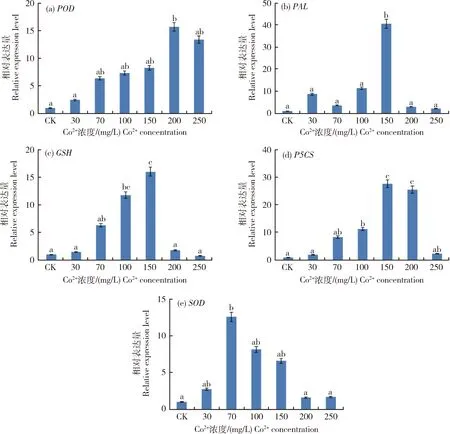
小寫字母表示在0.05水平的差異。
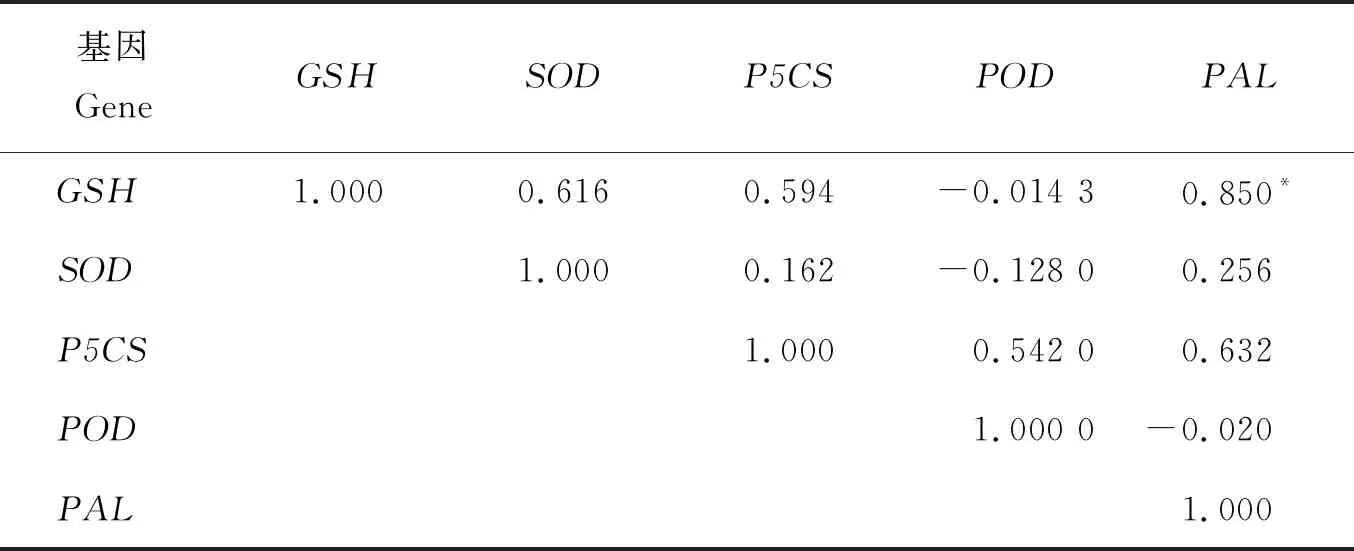
表2 鈷脅迫下青稞關鍵酶基因相對表達量間的相關分析
在30~150 mg/L Co2+濃度處理下,與對照組相比,青稞SOD、POD、P5CS、GSH和PAL基因的相對表達量均表現為增加,但是這些抗逆基因表達的響應時間順序不同。在200和250 mg/L Co2+處理下,P5CS和POD基因的相對表達量增加。結果顯示,Co2+脅迫下青稞幼苗中多種抗氧化酶表達量升高,共同提高植物抵御逆境脅迫的能力。
Co2+脅迫下青稞葉片響應重金屬脅迫的相關基因之間也存在著普遍的相關關系。PAL與GSH基因表達量的相關系數為0.850,達到顯著正相關關系。PAL與P5CS、POD與P5CS、GSH與SOD、GSH與P5CS基因的相關系數分別為0.632、0.542、0.616、0.594,均為正相關關系。SOD、P5CS、PAL、POD和GSH基因的最大表達量隨Co2+處理濃度的增加而依次增加,這些基因通過相互協調表達以便提高青稞幼苗對環境脅迫的響應能力,還有待進一步試驗驗證基因表達與逆境生理指標的關系。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
