miR-27b-3p 對膿毒癥肺內皮細胞YAP 信號表達的調控機制研究
2021-03-24 07:22:02黃愛華
浙江中西醫結合雜志 2021年3期
關鍵詞:小鼠
黃愛華 王 旸
敗血癥是重癥監護病房入院的主要原因,其死亡率和發病率均極高。在敗血病患者中,內皮細胞活化和功能障礙繼發的彌漫性血管滲漏和炎癥反應是多器官衰竭的主要原因[1-3]。膿毒癥中的內皮激活導致多種促炎細胞因子的產生以及粘附分子選擇素(E/P-selectins)和細胞間粘附分子-1(ICAM-1)的上調,導致多形核中性粒細胞大量浸潤,過度炎癥,組織損傷,并最終導致器官功能障礙[4-5]。Yes 相關蛋白(YAP)是Hippo 信號通路中的關鍵分子,在多種腫瘤中發揮著促癌作用[6]。此前研究發現,膿毒癥模型中,內皮細胞YAP 表達量增加,而內皮特異性缺失YAP 小鼠加重膿毒癥中血管功能障礙和彌漫性炎癥反應,提示YAP 在膿毒癥內皮細胞活化過程中起到負反饋調節的作用[7]。然而膿毒癥中YAP 表達量增加的原因尚未明確。研究表明,多種MicroRNA 在膿毒癥的多種細胞中發揮著重要的調控作用[8-9]。因此本研究擬探討膿毒癥小鼠miRNA 對YAP 的調控機制。
1 實驗材料
1.1 動 物 40 只SPF 級C57BL/6 小鼠(雄性,6~8周齡,體質量20~25g)購買并飼養于杭州醫學院實驗動物中心,生產許可證號SCXK(浙)2019-0002,使用許可證號SYXK(浙)2019-0011。飼養條件:SPF 級屏障中,白天黑夜各12h,給予充足食物和無菌水。所有操作均符合杭州醫學院實驗動物倫理委員會審查要求(倫理批號:20200126)。
1.2 試 劑 小鼠肺微血管內皮細胞完全培養基(批號025646)購自武漢普諾賽生物科技有限公司;腫瘤壞死因子-α(TNF-α,批號1321)、白介素-6(IL-6,批號03216、85165)等炎癥因子檢測試劑盒購自美國R&D Systems。抗體:YAP 兔單抗(批號16651)、ICAM 兔單抗(批號a2365)、選擇素(E-selectin)兔單抗(批號a32356)、磷酸化核因子激活的B 細胞的κ-輕鏈增強p65 亞基(p-p65)兔單抗(批號a21320)、核因子激活的B 細胞的κ-輕鏈增強p65 亞基(p65)兔單抗(批號a15135)均購自美國CST 公司。甘油醛-3-磷酸脫氫酶(GAPDH)作為內參。mimic NC 及miR-27b-3p mimic(批號15461)購自西格瑪奧德里奇(上海)貿易有限公司。
1.3 儀 器 酶標儀SpectraMax 5 購自美谷分子儀器(上海)有限公司;CFX96 Touch 熒光定量PCR 儀,Trans-Blot Turbo 全能型蛋白轉印系統均購自伯樂生命醫學產品(上海)有限公司;Odyssey TMLi-COR 檢測系統購自上海基因有限公司;-80℃超低溫冰箱購自美國Thermo fisher。
2 實驗方法
2.1 小鼠CLP 模型構建 盲腸結扎穿孔(CLP)模型構建參考文獻[10]進行操作。將40 只SPF 級C57BL/6 小鼠按照隨機數字表法分為對照組、CLP 組、miRNA 類似物(mimic NC)組、miR-27b-3p 類似物(miR-27b-3p mimic)組,每組10 只。CLP 組、mimic NC 組、miR-27b-3p 組使用三棱針穿刺盲腸輕輕地壓縮擠出少量盲腸內容物誘導小鼠膿毒癥發生,對照組進行假手術。術后miR-27b-3p 組小鼠予尾靜脈注射miR-27b-3p mimic(1×108/mL,200μL/只),mimic NC 組小鼠予尾靜脈注射mimic NC 陰性對照物(1×108/mL,200μL/只)。24h 后取小鼠肺組織分離小鼠原代肺內皮細胞進行后續實驗。
2.2 小鼠原代肺內皮細胞培養及處理 將上述四組小鼠原代肺內皮細胞提取出來后進行體外培養及后續實驗。小鼠原代肺內皮細胞利用小鼠肺微血管內皮細胞完全培養基進行培養,細胞長至90%時傳代或鋪板。取2~4 代細胞進行后續實驗。
2.3 小鼠肺內皮上清TNF-α、IL-6 含量測定 上述四組細胞體外鋪板培養24h 后,取上清經過離心,過0.22μm 濾膜,應用酶聯免疫吸附試驗(ELISA)試劑盒檢測上清TNF-α、IL-6 含量,應用SpectraMax 5檢測450nm 處吸光度值,取3 次獨立重復實驗數據進行統計分析。
2.4 qRT-PCR 檢測miR-27b-3p、YAP、ICAM、Eselectin mRNA 水平 將上述四組細胞鋪板于6 孔板中,24h 后提取細胞總RNA,經過逆轉為cDNA后,進行qRT-PCR(熒光定量PCR),qRT-PCR 引物如下:miR-27b-3p(Forward:5' ATGACACCAAGGACCAGAGC 3',Reverse:5' GTGTAAGGACCCATCGGAGA 3');YAP(Forward:5' CAAGAAAGCAGGCTCACAGAA 3',Reverse:5' GCTGGGTGTTAGGGCTTCG 3');ICAM(Forward:5' GAAATGCCACCTTTTGACAGTG 3',Reverse:5' CTGGATGCTCTCATCAGGACA 3');E-selectin(Forward:5' GGCGTTAGAAAGCATCCTTCC 3',Reverse:5' GCAGAGGGCACACTCAAAGT 3')。所有引物均購自南京金斯瑞生物科技有限公司。
2.5 Western blot 法檢測YAP、ICAM、E-selectin 及p-p65 蛋白水平 將上述四組細胞鋪板24h 后,應用蛋白裂解液裂解細胞蛋白,經過BCA 法測定蛋白濃度后各取20μg/孔進行蛋白免疫印跡(Western blot),經過電泳、轉膜、封閉后,于4℃孵育一抗過夜,一 抗 包 括YAP、ICAM、E -selectin、p -p65、p65 及GAPDH。洗去一抗后室溫孵育熒光二抗1h。應用Odyssey TMLi-COR 檢測系統進行熒光定量檢測。
2.6 統計學方法 應用SPSS 20.0 軟件進行統計分析,計量資料采用均數±標準差()表示,多組間比較采用單因素方差分析,兩兩比較采用SNK-q 檢驗。P<0.05 表示差異有統計學意義。
3 實驗結果
3.1 CLP 模型小鼠內皮細胞YAP、ICAM、E-selectin mRNA 表達水平 應用CLP 誘導小鼠膿毒癥模型,取小鼠肺組織進行HE 染色,病理觀察顯示,對照組小鼠肺泡結構基本完整,無炎性細胞浸潤。而CLP 組小鼠肺組織具有嚴重的組織學改變,包括肺泡充血、滲出液和炎性細胞浸潤。進一步qRT-PCR 結果顯示,與假手術組小鼠比較,CLP 組小鼠肺內皮細胞YAP、ICAM、E-selectin mRNA 表達水平明顯升高(P<0.05)。見表1,圖1。
表1 各組小鼠肺內皮細胞YAP、ICAM、E-selectin mRNA相對表達量()
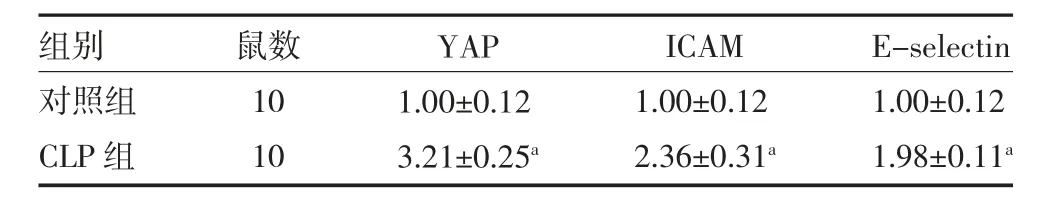
表1 各組小鼠肺內皮細胞YAP、ICAM、E-selectin mRNA相對表達量()
注:對照組小鼠進行假手術操作;CLP 組小鼠通過CLP 誘導膿毒癥;CLP 為盲腸結扎穿孔;YAP 為Yes 相關蛋白;ICAM 為細胞間粘附分子;E-selectin 為選擇素;與對照組比較,aP<0.05

圖1 CLP 模型小鼠肺組織形態(HE 染色×400)
3.2 CLP 小鼠肺內皮細胞miR-27b-3p 表達水平miRNA 靶基因數據庫PITA 預測調控YAP 的miRNA 為miR-27b-3p,預測結果顯示,miR-27b-3p 與YAP mRNA 3' UTR 序列存在6 個完全互補配對堿基UGUGAA。進一步qRT-PCR 檢測CPL 組小鼠肺內皮細胞miR-27b-3p 表達,結果顯示,與對照組(1.00±0.11)比較,CLP 組miR-27b-3p(0.31±0.15)表達明顯下調(P<0.05)。見圖2。

圖2 miR-328-5p 靶向SPON2 基因3'UTR 序列
3.3 miR-27b-3p 調控YAP 表達及內皮激活 與對照組比較,CLP 組YAP、ICAM、E-selectin 表達水平明顯升高,同時p-p65 磷酸化活化水平明顯升高;與mimic NC 組比較,miR-27b-3p mimic 組ICAM、Eselectin 表達水平明顯升高,同時p-p65 磷酸化活化水平明顯升高。見圖3。
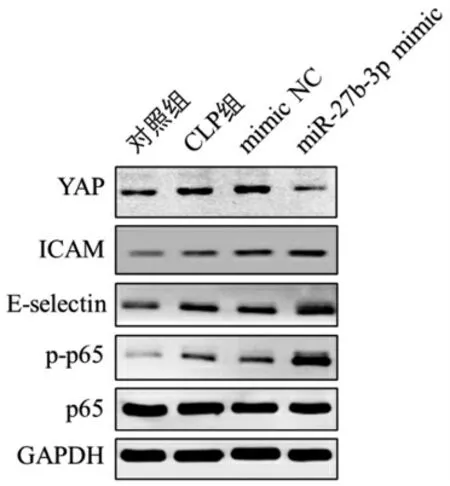
圖3 各組小鼠肺內皮YAP、ICAM、E-selectin 表達及p65 活化水平
3.4 miR-328-5p 直接靶向SPON2 抑制其表達 與對照組比較,CLP 組小鼠內皮細胞上清液中TNF-α、IL-6 的含量明顯升高(P<0.05);與mimic NC 組比較,miR-27b-3p mimic 組小鼠內皮細胞上清液中TNF-α、IL-6 的含量明顯升高(P<0.05)。見表2。
表2 各組內皮細胞上清液TNF-α、IL-6 水平比較(pg/mL,)

表2 各組內皮細胞上清液TNF-α、IL-6 水平比較(pg/mL,)
注:對照組小鼠進行假手術操作;CLP 組小鼠通過CLP 誘導膿毒癥;mimic NC 組小鼠通過CLP 誘導膿毒癥,同時術后尾靜脈注射mimic NC 陰性對照物;miR-27b-3p mimic 組小鼠通過 CLP 誘導膿毒癥,同時術后尾靜脈注射miR-27b-3p mimic;TNF-α 為腫瘤壞死因子-α;IL-6 為白介素-6;CLP 為盲腸結扎穿孔;與對照組比較,aP<0.05;與mimic NC 組比較,bP<0.05
4 討論
創傷、休克及感染導致的血管內皮功能障礙和炎癥反應是膿毒癥發生的主要機制[11-12]。調控內皮細胞異常激活對防止膿毒癥發生、降低死亡率具有重要的意義。miRNA 是一類微小RNA,在基因轉錄調控中發揮著重要的作用。Essandoh 和Fan[13]通過芯片篩選發現,膿毒癥組和健康對照組人群之間共有22個差異表達的miRNA,其中9 個表達上調、13 個表達下調。Qiu 等[14]發現長非編碼RNA TUG1 通過調控miR-34b-5p 表達,參與膿毒癥過程中內皮細胞的活化和炎癥反應。本研究發現,miR-27b-3p 在膿毒癥內皮細胞中低表達(P<0.05),提示miR-27b-3p 在內皮細胞穩態中可能發揮著重要的調控作用。
YAP 是一種轉錄共激活因子,通過促進細胞增殖和抑制細胞凋亡而成為組織生長和器官大小的關鍵調控分子。心臟和血管平滑肌細胞YAP 在心血管發育過程中也起到重要的調控作用[15-16]。而在膿毒癥研究中發現,內皮細胞缺失YAP 小鼠內皮細胞異常激活,炎癥反應異常加劇[17]。本研究同樣發現,膿毒癥模型小鼠內皮細胞YAP 表達量明顯升高(P<0.05),同時在膿毒癥模型小鼠尾靜脈注射miR-27b-3p mimic 后,內皮細胞YAP 表達量明顯下調(P<0.05)。研究報道,內毒素作用于內皮細胞可以激活打孔蛋白(Gasdermin D),進而釋放線粒體DNA 到胞質中,后者作為第二形式參與調控YAP1 信號來抑制內皮細胞炎癥反應[18]。然而YAP 調控的炎癥下游信號并未明確。基于此,本研究結果顯示,miR-27b-3p mimic 組p-p65 亞基磷酸化水平明顯升高,下游激活標志物ICAM、E-selectin 表達明顯上調,同時內皮細胞分泌的炎癥因子TNF-α、IL-6 水平明顯上調(P<0.05),表明miR-27b-3p 可能負向調控YAP 表達,參與調控膿毒癥過程中內皮細胞的活化以及炎癥反應。
綜上所述,膿毒癥內皮細胞miR-27b-3p 表達下調,通過靶向調控YAP 轉錄水平,促進炎癥相關信號p65 亞基的激活,導致下游內皮細胞活化相關蛋白ICAM、E-selectin 表達增加,加劇膿毒癥炎癥反應。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
